The Cytotoxic Effect of Genistein, a Soybean Isoflavone, against Cultured Tribolium Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Cell Culture
2.3. Isoflavones and Organic Compounds
2.4. Dietary Intake Assay
2.5. Survival of T. Castaneum Adults
2.6. Cell Survival and Cytotoxicity Assay
3. Results
3.1. Dietary Intake of Gypsum Supplemented with Soy Flour by T. Castaneum
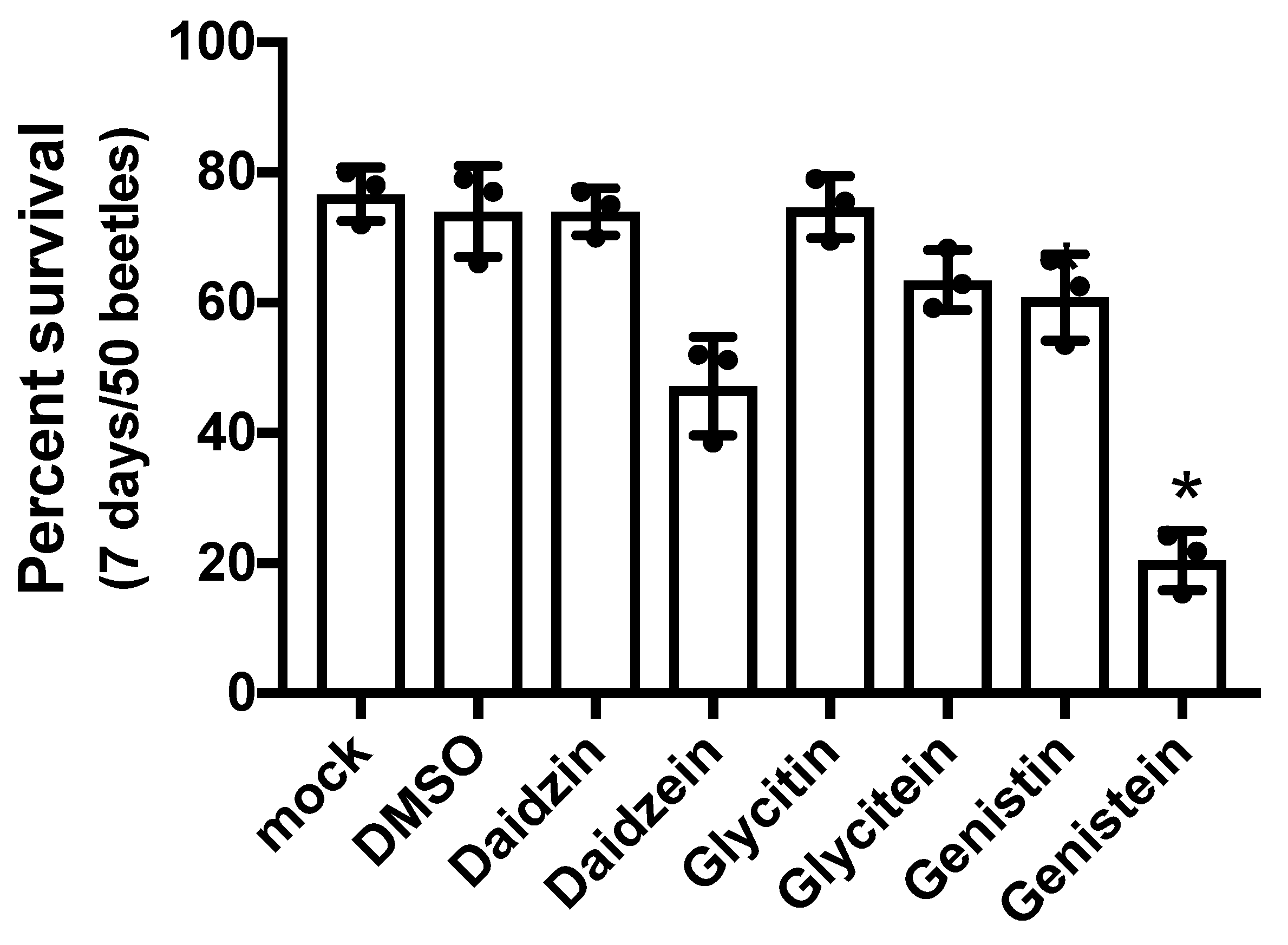
3.2. Lifespan of T. Castaneum Adults Supplemented with Soy Flour and Isoflavone
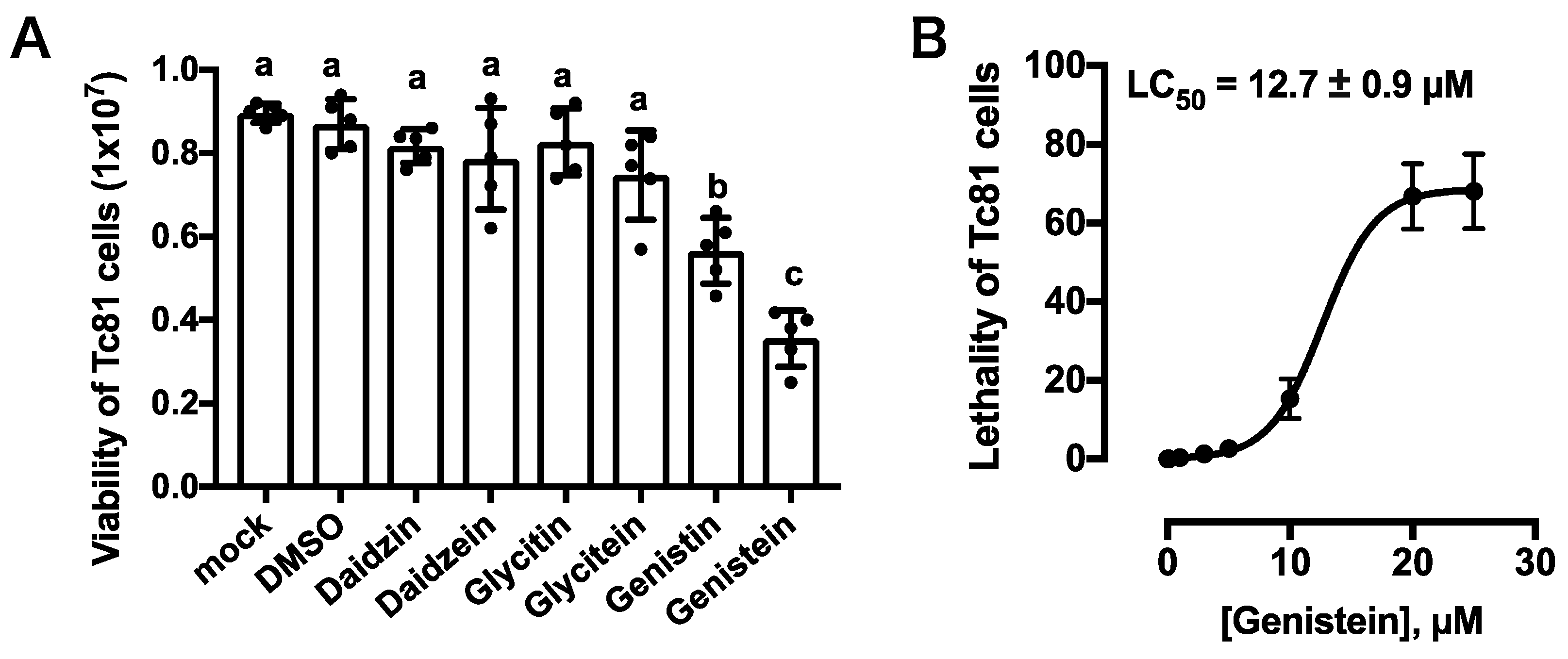
3.3. Cytotoxicity of Tc81 Cultured Cells by Isoflavones
3.4. Colorimetry Based Cell Viability Assay for Tc81 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- LeCato, G.L. Red flour beetle: Population growth on diet of corn, wheat rice or shelled peanuts supplemented with eggs or adult of the Indian meal moth. J. Econ. Entomol. 1975, 68, 763–765. [Google Scholar] [CrossRef]
- Hagstrum, D. Atlas of Stored-Product Insects and Mites, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780128104316. [Google Scholar]
- Takada, T.; Sato, R.; Kikuta, S. A mannitol/sorbitol receptor stimulates dietary intake in Tribolium castaneum. PLoS ONE 2017, 12, e0186420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Good, N.E. The flour beetles of the genus Tribolium. USDA Tech. Bul. 1936, 498, 1–57. [Google Scholar]
- Lin, S.; Richard, G. Studies on the nutritional value of soybean flour to Tribolium confusum duv. J. N. Y. Entomol. Soc. 1952, 60, 107–118. [Google Scholar]
- Mickel, C.E.; Standish, J. Susceptibility of Processed Soy Flour and Soy Grids in Storage to Attack by Tribolium castaneum (Herbst); University of Minnesota Agricultural Experiment Station: St. Paul, MN, USA, 1947; p. 178. [Google Scholar]
- Sokoloff, A.; Franklin, I.R.; Overton, L.F.; Ho, F.K. Comparative studies with Tribolium (Coleoptera, Tenebrionidiae)—I: Productivity of T. castaneum (Herbst) and T. confusum Duv. On several commercial-available diets. J. Stored Prod. Res. 1966, 1, 295–311. [Google Scholar]
- Naseri, B.; Borzoui, E.; Majd, S.; Mozaffar Mansouri, S. Influence of different food commodities on life history, feeding efficiency, and digestive enzymatic activity of Tribolium castaneum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2017, 110, 2263–2268. [Google Scholar] [CrossRef]
- Astuti, L.P.; Rizali, A.; Firnanda, R.; Widjayanti, T. Physical and chemical properties of flour products affect the development of Tribolium castaneum. J. Stored Prod. Res. 2020, 86, 101555. [Google Scholar] [CrossRef]
- Markley, K.S.; Goss, W.H. Soybean Chemistry and Technology; Chemical Publishing: New York, NY, USA, 1944; pp. 137–143. [Google Scholar]
- Osborne, T.B.; Mendel, L.B. The nutritive value of the wheat kernel and its milling products. J. Biol. Chem. 1919, 37, 557–601. [Google Scholar]
- Taggart, P. Starch as an ingredient: Manufacture and applications. In Starch in Food Structure, Function and Applications; Eliasson, A.-C., Ed.; Woodhead Publishing: Cambridge, UK, 2004; pp. 363–392. [Google Scholar]
- Fraenkel, G.; Blewett, M. The basic food requirements of several insects. J. Exp. Biol. 1943, 20, 28–34. [Google Scholar]
- Harborne, J.B. The Flavonoids: Advances in Research Since 1980; Springer US: New York, NY, USA, 1988; pp. 125–209. [Google Scholar]
- Simmonds, M.S.; Stevenson, P.C. Effects of isoflavonoids from Cicer on larvae of Heliocoverpa armigera. J. Chem. Ecol. 2001, 5, 965–977. [Google Scholar] [CrossRef]
- Kubo, I. New concept to search for alternate insect control agents from plants. In Naturally Occurring Bioactive Compounds, 1st ed.; Rai, M., Carpinella, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 3, pp. 61–80. [Google Scholar]
- Hohenstein, J.D.; Studham, M.E.; Klein, A.; Kovinich, N.; Barry, K.; Lee, Y.J.; MacIntosh, G.C. Transcriptional and chemical changes in soybean leaves in response to long-term aphid colonization. Front. Plant Sci. 2019, 10, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.D. Potential for using semiochemicals to protect stored products from insect infestation. J. Stored Prod. Res. 2004, 40, 1–25. [Google Scholar] [CrossRef]
- Goławska, S.; Lukasik, I. Antifeedant activity of luteolin and genistein against the pea aphid, Acyrthosiphon pisum. J. Pest Sci. 2012, 85, 443–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, S.; Duncan, A.M.; Yang, R.; Marcone, M.F.; Rajcan, I.; Tsao, R. Tracking isoflavones: From soybean to soy flour, soy protein isolates to functional soy bread. J. Funct. Foods 2009, 1, 119–127. [Google Scholar] [CrossRef]
- Zavala, J.A.; Mazza, C.A.; Dillon, F.M.; Chludil, H.D.; Ballare, C.L. Soybean resistance to stink bugs (Nezara viridula and Piezodorus guildinii) increases with exposure to solar UV-B radiation and correlates with isoflavonoid content in pods under field conditions. Plant Cell Environ. 2015, 38, 920–928. [Google Scholar] [CrossRef]
- Piubelli, G.C.; Hoffmann-Campo, C.B.; Moscardi, F.; Miyakubo, S.H.; Neves De Oliveira, M.C. Are chemical compounds important for soybean resistance to Anticarsia gemmatalis? J. Chem. Ecol. 2005, 31, 1509–1525. [Google Scholar] [CrossRef]
- Xie, Y.S.; Bodnaryk, R.P.; Fields, P.G. A rapid and simple flour-disk bioassay for testing substances active against stored-product insects. Can. Entomol. 1996, 128, 865–875. [Google Scholar] [CrossRef]
- Kikuta, S. TribUTE-F assay: Fluorescence-based rapid quantification of dietary intake in the red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae), facilitates evaluation of antifeedant inhibitory effects. J. Stored Prod. Res. 2018, 77, 122–125. [Google Scholar] [CrossRef]
- Kikuta, S. Deployment of an attractive toxic sugar bait system (ATSB) with insecticide, for adult Tribolium castaneum (Coleoptera: Tenebrionidae). J. Stored Prod. Res. 2019, 83, 97–102. [Google Scholar] [CrossRef]
- Kayukawa, T.; Tateishi, K.; Shinoda, T. Establishment of a versatile cell line for juvenile hormone signaling analysis in Tribolium castaneum. Sci. Rep. 2013, 3, 1570. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Wu, Y.; Wu, H.; Wu, Y.; Wu, H.; Wang, W. Functional analysis of a survivin-like gene in Bombyx mori. Cytotechnology 2014, 1, 181–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, P.; Smith, C.M.; Fronczek, F.R.; Fischer, N.H. Isoflavones from an insect-resistant variety of soybean and the molecular structure of afrormosin. J. Nat. Prod. 1986, 49, 1126–1129. [Google Scholar] [CrossRef]
- Simmonds, M.S. Importance of flavonoids in insect-plant interactions: Feeding and oviposition. Phytochemistry 2001, 56, 245–252. [Google Scholar] [CrossRef]
- Mullin, C.A.; Alfatafta, A.A.; Harman, J.L.; Everett, S.L.; Serino, A.A. Feeding and toxic effects of floral sesquiterpene lactones, diterpenes, and phenolics from sunflower on western corn rootworm. J. Agric. Food Chem. 1991, 39, 2293–2299. [Google Scholar] [CrossRef]
- Morimoto, M.; Kumeda, S.; Komai, K. Insect antifeedant flavonoids from Gnaphalium affine D. Don. J. Agric. Food Chem. 2000, 48, 1888–1891. [Google Scholar] [CrossRef]
- Smagghe, G.; Goodman, C.L.; Stanley, D. Insect cell culture and applications to research and pest management. In Vitro Cell. Dev. Biol. Anim. 2009, 45, 93–105. [Google Scholar] [CrossRef]
- Silver, K.; Jiang, H.; Fu, J.; Phillips, T.W.; Beeman, R.W.; Park, Y. The Tribolium castaneum cell line TcA: A new tool kit for cell biology. Sci. Rep. 2014, 4, 6840. [Google Scholar] [CrossRef] [Green Version]
- Yooboon, T.; Kuramitsu, K.; Bullangpoti, V.; Kainoh, Y.; Furukawa, S. Cytotoxic effects of β-asarone on Sf9 insect cells. Arch. Insect Biochem. Physiol. 2019, 102, e21596. [Google Scholar] [CrossRef]
- Rao, K.V.; Chattopadhyay, S.K.; Reddy, G.C. Flavonoids with mosquito larval toxicity. J. Agr. Food Chem. 1990, 38, 1427–1430. [Google Scholar] [CrossRef]
- Hubert, J.; Stejskal, V.; Aspaly, G.; Münzbergová, Z. Suppressive Potential of Bean (Phaseolus vulgaris) Flour Against Five Species of Stored-Product Mites (Acari: Acarididae). J. Econ. Entomol. 2014, 100, 586–590. [Google Scholar] [CrossRef]
- Yuan, E.; Yan, H.; Gao, J.; Guo, H.; Ge, F.; Sun, Y. Increases in genistein in Medicago sativa confer resistance against the Pisum host race of Acyrthosiphon pisum. Insects 2019, 10, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.T.; Sathyamoorthy, N.; Phang, J.M. Molecular effects of genistein on estrogen receptor mediated pathways. Carcinogenesis 1996, 2, 271–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimbach, G.; Boesch-Saadatmandi, C.; Frank, J.; Fuchs, D.; Wenzel, U.; Daniel, H.; Hall, W.L.; Weinberg, P.D. Dietary isoflavones in the prevention of cardiovascular disease—A molecular perspective. Food Chem. Toxicol. 2008, 46, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Wang, L.; Jin, Y.; Zhen, H.; Xu, P.; Xu, Y.; Li, C.; Xu, H. Role of metabolism in the effects of genistein and its phase II conjugates on the growth of human breast cell lines. AAPS J. 2012, 14, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, G.; Roy, D.; Khemka, V.K.; Chattopadhyay, M.; Chakrabarti, S. Genistein, the isoflavone in soybean, causes amyloid beta peptide accumulation in human neuroblastoma cell line: Implications in Alzheimer’s disease. Aging Dis. 2015, 6, 456–465. [Google Scholar] [CrossRef] [Green Version]
- Dang, Z.C.; Audinot, V.; Papapoulos, S.E.; Boutin, J.A.; Löwik, C.W. Peroxisome.proliferator-activated receptor gamma (PPARgamma) as a molecular target for the soy phytoestrogen genistein. J. Biol. Chem. 2003, 278, 962–967. [Google Scholar] [CrossRef] [Green Version]
- Hikal, W.M.; Baeshen, R.S.; Said-Al Ahl, H.A. Botanical insecticide as simple extractives for pest control. Cogent Biol. 2017, 3, 1404274. [Google Scholar] [CrossRef]





© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikuta, S. The Cytotoxic Effect of Genistein, a Soybean Isoflavone, against Cultured Tribolium Cells. Insects 2020, 11, 241. https://doi.org/10.3390/insects11040241
Kikuta S. The Cytotoxic Effect of Genistein, a Soybean Isoflavone, against Cultured Tribolium Cells. Insects. 2020; 11(4):241. https://doi.org/10.3390/insects11040241
Chicago/Turabian StyleKikuta, Shingo. 2020. "The Cytotoxic Effect of Genistein, a Soybean Isoflavone, against Cultured Tribolium Cells" Insects 11, no. 4: 241. https://doi.org/10.3390/insects11040241
APA StyleKikuta, S. (2020). The Cytotoxic Effect of Genistein, a Soybean Isoflavone, against Cultured Tribolium Cells. Insects, 11(4), 241. https://doi.org/10.3390/insects11040241