Asteraceae Paradox: Chemical and Mechanical Protection of Taraxacum Pollen




Abstract
:1. Introduction
2. Material and Methods
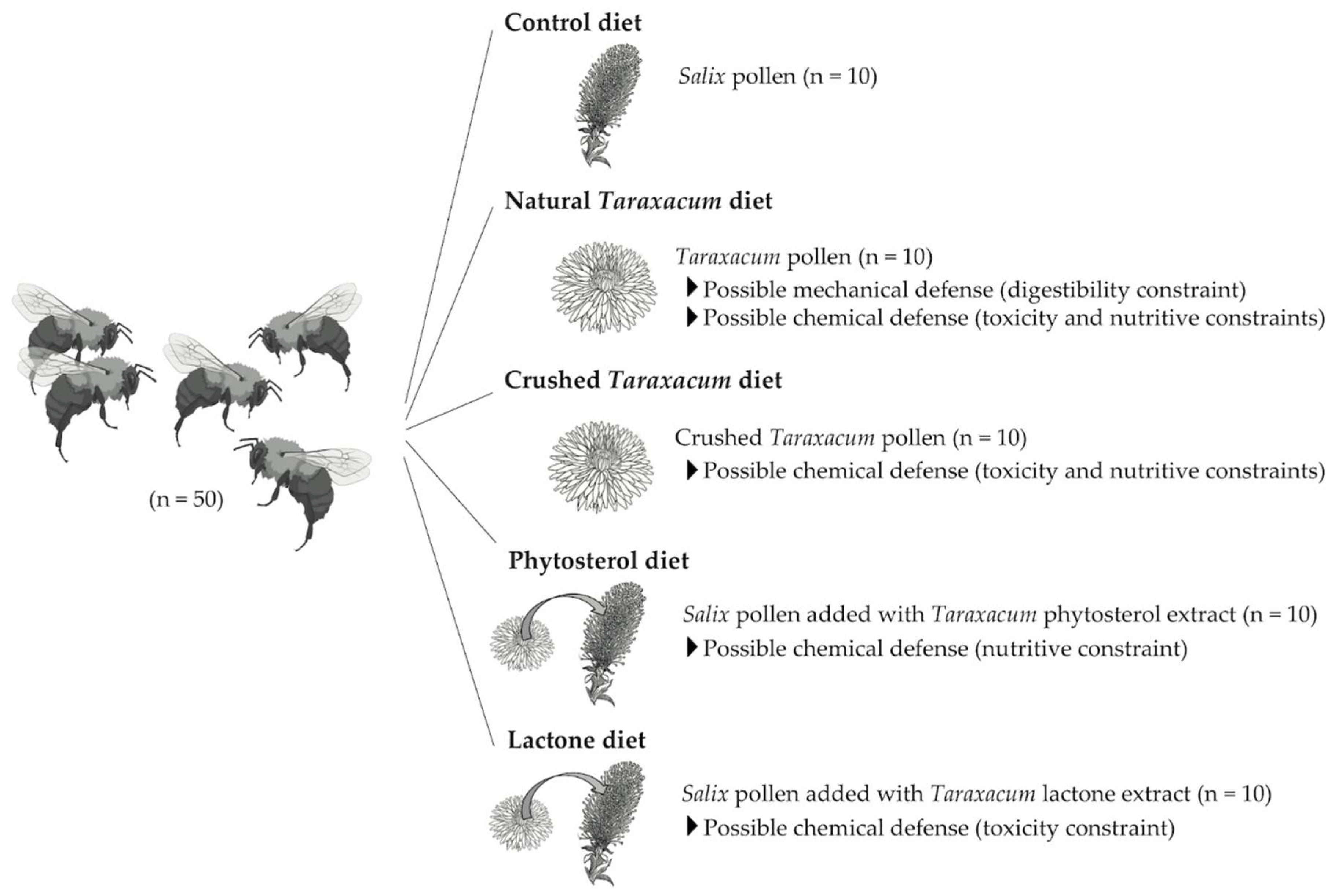
2.1. Model System
2.2. Pollen Diets
2.3. Bioassays
2.3.1. Experimental Design
2.3.2. Micro-Colony Performance
2.3.3. Digestive Damages
2.4. Data Analysis
3. Results
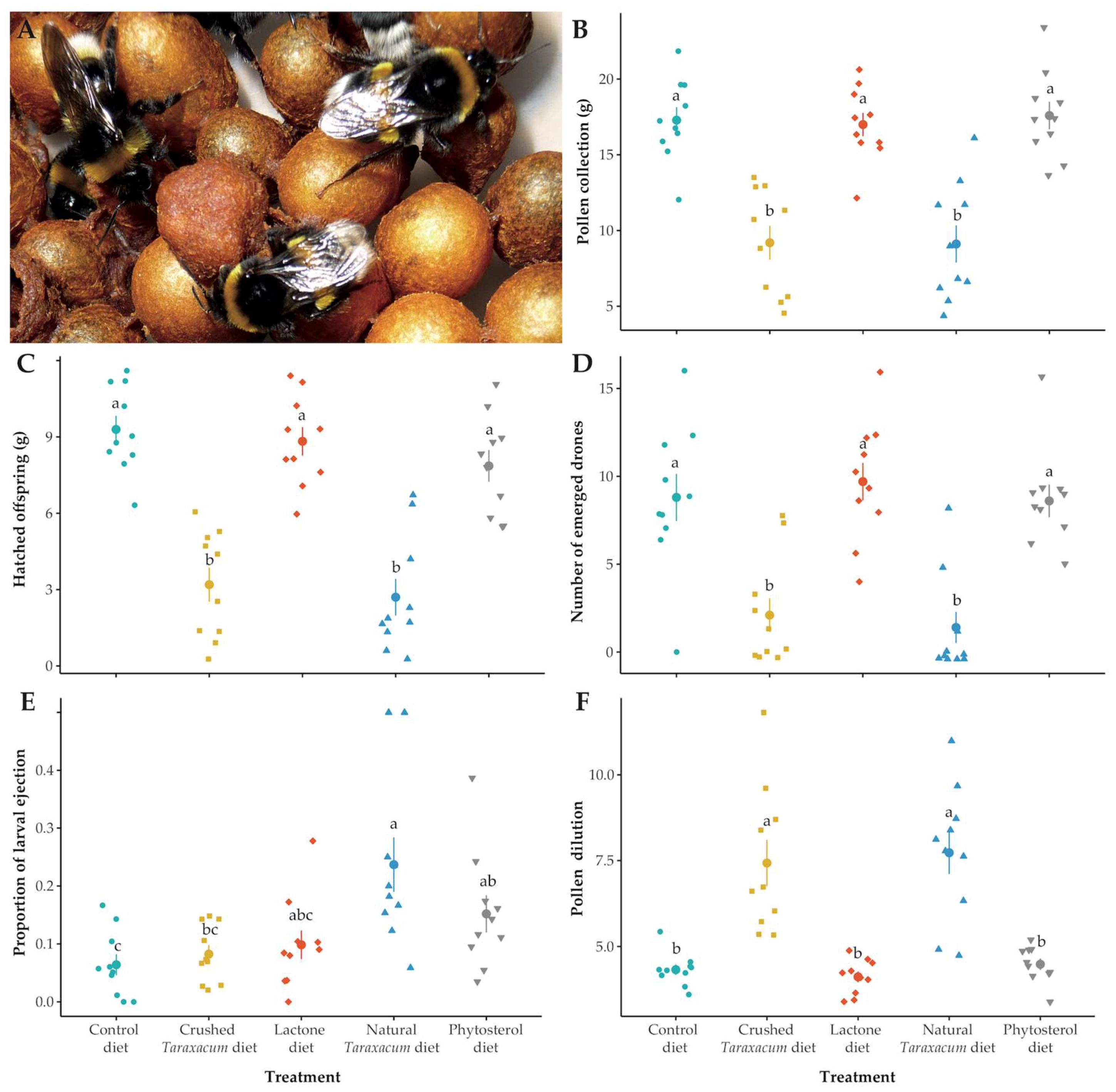
3.1. Micro-Colony Performance
3.1.1. Resource Collection
3.1.2. Fitness
3.1.3. Stress Response
3.2. Digestive Damage
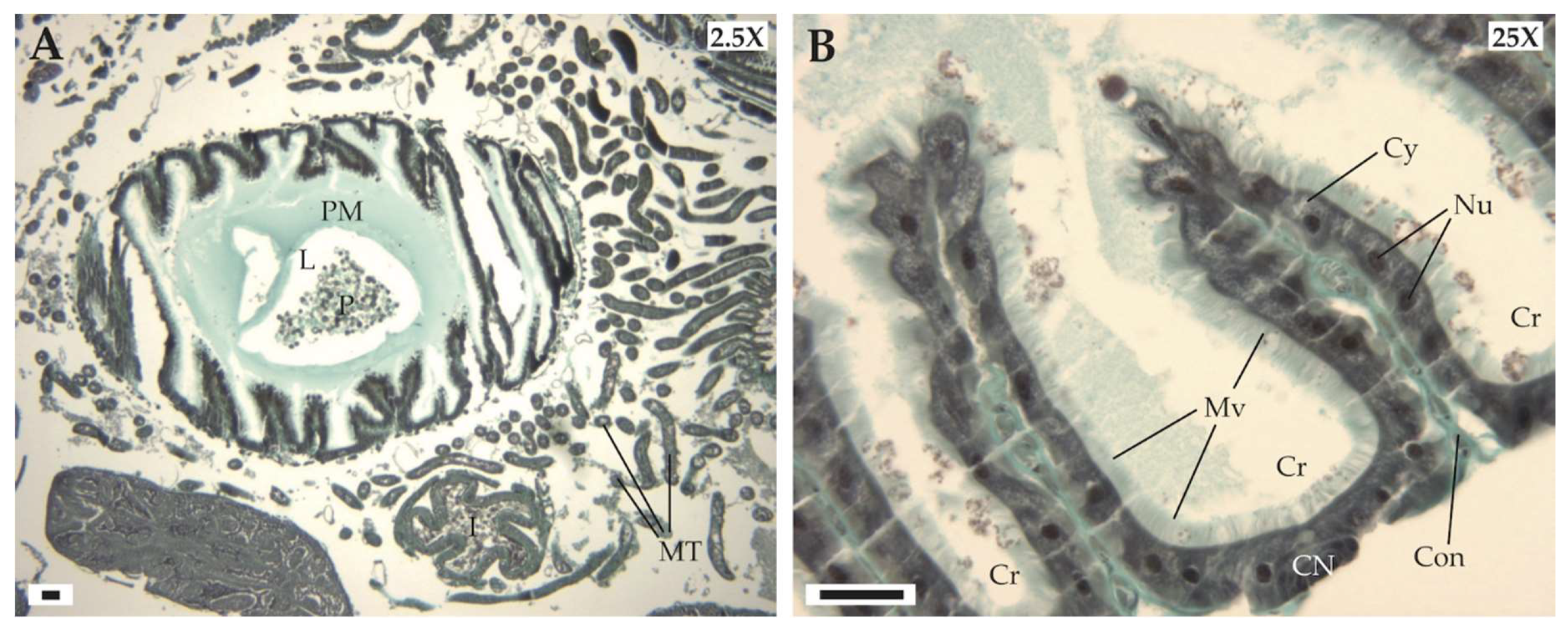
3.2.1. General Histology
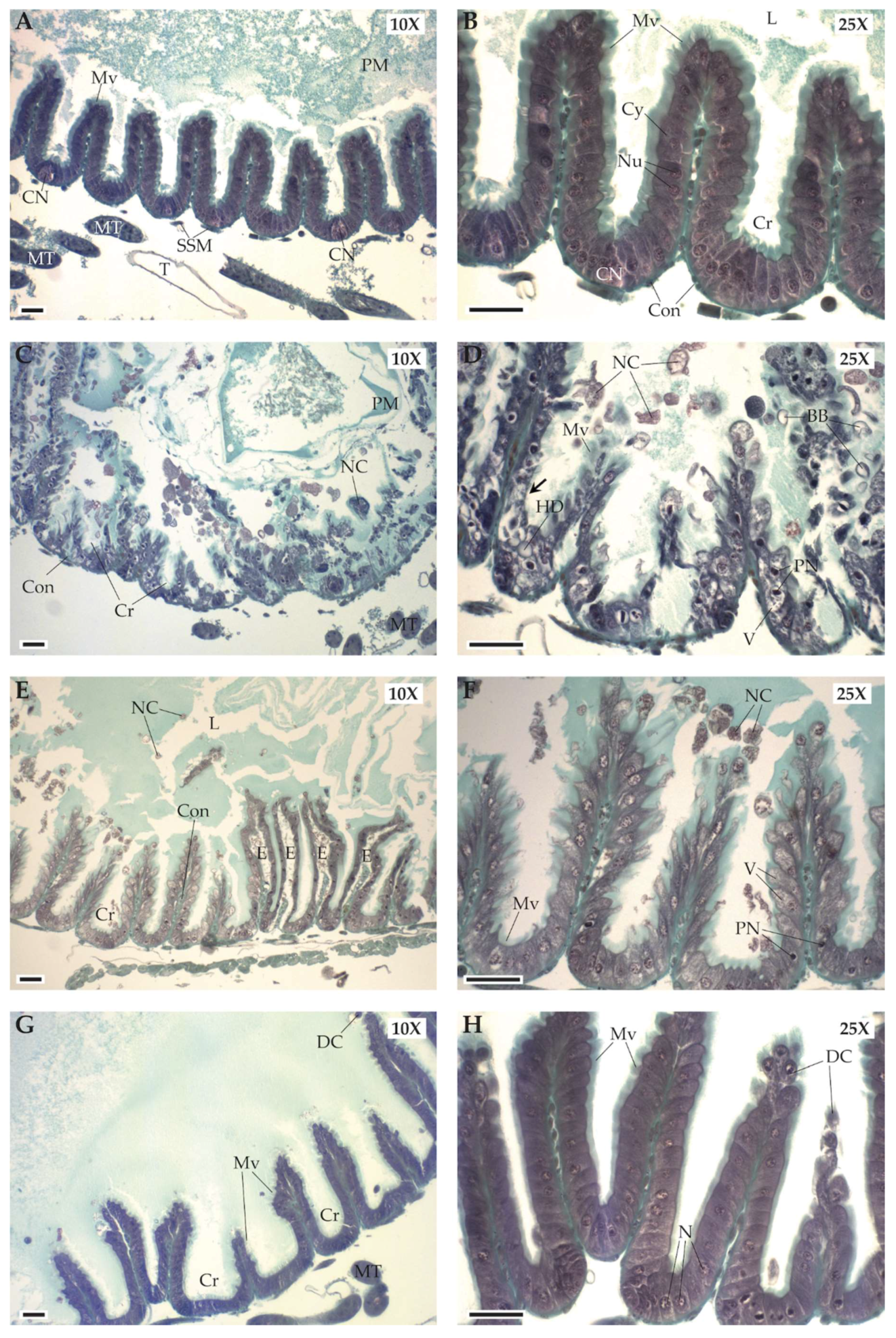
3.2.2. Treatment Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Westrich, P. Die Wildbienen Baden-Württembergs; Eugen Ulmer: Suttgart, Germany, 1989. [Google Scholar]
- Michener, C.D. The Bees of the World, 2nd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Cane, J.H.; Sipes, S.D. Characterizing floral specialization by bees: Analytical methods and revised lexicon for oligolecty. In Plant–Pollinator Interactions from Specialization to Generalization; Waser, N.M., Ollerton, J., Eds.; The University of Chicago Press: Chicago, IL, USA, 2006; pp. 99–122. [Google Scholar]
- Müller, A.; Kuhlmann, M. Pollen hosts of western palaearctic bees of the genus Colletes (Hymenoptera: Colletidae): The Asteraceae paradox. Biol. J. Linn. Soc. 2008, 95, 719–733. [Google Scholar] [CrossRef]
- Dötterl, S.; Vereecken, N.J. The chemical ecology and evolution of bee-flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697. [Google Scholar] [CrossRef]
- Müller, A. Convergent evolution of morphogical specializations in Central European bee and honey wasp species as an adaptation to the uptake of pollen from nototribic flowers (Hymenoptera, Apoidea and Masaridae). Biol. J. Linn. Soc. 1996, 57, 235–252. [Google Scholar] [CrossRef]
- Müller, A.; Diener, S.; Schnyder, S.; Stutz, K.; Sedivy, C.; Dorn, S. Quantitative pollen requirements of solitary bees: Implications for bee conservation and the evolution of bee-flower relationships. Biol. Conserv. 2006, 130, 604–615. [Google Scholar] [CrossRef]
- Westerkamp, C.; Claben-Bockhoff, R. Bilabiate Flowers: The Ultimate Response to Bees? Ann. Bot. 2007, 100, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlindwein, C.; Wittmann, D.; Martins, C.F.; Hamm, A.; Siqueira, J.A.; Schiffer, D.; Machado, I.C. Pollination of Campanula rapunculus L. (Campanulaceae): How much pollen flows into pollination and intro reproduction of oligolectic pollinators? Plant Syst. Evol. 2005, 250, 147–156. [Google Scholar] [CrossRef]
- Funk, V.A.; Bayer, R.J.; Keeley, S.; Chan, R.; Watson, L.; Gemeinholzer, B.; Schilling, E.; Panrelo, J.L.; Baldwin, B.G.; Garcia-Jacas, N.; et al. Everywhere but Antartica: Using a supertree to understand the diversity and distribution of the Compositae. Biol. Skr. 2005, 55, 343–373. [Google Scholar]
- Goulson, D.; Hanley, M.E.; Darvill, B.; Ellis, J.S.; Knight, M.E. Causes of rarity in bumblebees. Biol. Conserv. 2005, 122, 1–8. [Google Scholar] [CrossRef]
- Kleijn, D.; Raemakers, I. A retrospective analysis of pollen host plant use by stable and declining bumble bee species. Ecology 2008, 89, 1811–1823. [Google Scholar] [CrossRef]
- Levin, M.D.; Haydak, M.H. Comparative value of different pollens in the nutrition of Osmia lignaria. Bee World 1957, 38, 221–226. [Google Scholar] [CrossRef]
- Guirguis, G.N.; Brindley, W.A. Insecticide susceptibility and response to selected pollens of larval alfalfa leafcutting bees, Megachile pacifica (Panzer) (Hymenoptera: Megachilidae). Environ. Entomol. 1974, 3, 691–694. [Google Scholar] [CrossRef]
- Génissel, A.; Aupinel, P.; Bressac, C.; Tasei, J.N.; Chevrier, C. Influence of pollen origin on performance of Bombus terrestris micro-colonies. Entomol. Exp. Appl. 2002, 104, 329–336. [Google Scholar] [CrossRef]
- Williams, N.M. Use of novel pollen species by specialist and generalist solitary bees (Hymenoptera: Megachilidae). Oecologia 2003, 134, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Praz, C.J.; Müller, A.; Dorn, S. Specialized bees fail to develop on non-host pollen: Do plants chemically protect their pollen? Ecology 2008, 89, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Sedivy, C.; Müller, A.; Dorn, S. Closely related pollen generalist bees differ in their ability to develop on the same pollen diet: Evidence for physiological adaptations to digest pollen. Funct. Ecol. 2011, 25, 718–725. [Google Scholar] [CrossRef]
- Haider, M.; Dorn, S.; Müller, A. Intra- and inter- populational variation in the ability of a solitary bee species to develop on non-host-pollen: Implications for host range expansion. Funct. Ecol. 2013, 27, 255–263. [Google Scholar] [CrossRef]
- Tasei, J.N.; Aupinel, P. Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumble bee workers (Bombus terrestris, Hymenoptera: Apidae). Apidologie 2008, 39, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Vanderplanck, M.; Declèves, S.; Roger, N.; Decroo, C.; Caulier, G.; Glauser, G.; Gerbaux, P.; Lognay, G.; Richel, A.; Escaravage, N.; et al. Is non-host pollen suitable for generalist bumblebees? Insect Sci. 2018, 25, 259–272. [Google Scholar] [CrossRef]
- Bennett, R.R.; Wallsgrove, R.M. Tansley review no. 72. Secondary metabolites in plant defense mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Roberts, M.F.; Wink, M. Alkaloids—Biochemistry, Ecological Functions and Medical Applications; Plenum Press: New York, NY, USA, 1998. [Google Scholar]
- Baker, H.G. Non-sugar constituents of nectar. Apidologie 1997, 8, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Detzel, A.; Wink, M. Attraction, deterrence or in- toxication of bees (Apis mellifera) by plant allelochemicals. Chemoecology 1993, 4, 8–18. [Google Scholar] [CrossRef]
- Adler, L.S. The ecological significance of toxic nectar. Oikos 2000, 91, 409–420. [Google Scholar] [CrossRef]
- Dobson, H.E.M.; Bergström, G. The ecology and evolution of pollen odors. Plant Syst. Evol. 2000, 222, 63–87. [Google Scholar] [CrossRef]
- London-Shafir, I.; Shafir, S.; Eisikowitch, D. Amygdalin in almond nectar and pollen–Facts and possible roles. Plant Syst. Evol. 2003, 238, 87–95. [Google Scholar] [CrossRef]
- Kevan, P.G.; Ebert, T. Can almond nectar and pollen poison honey bees? Am. Bee J. 2005, 145, 507–509. [Google Scholar]
- Kempf, M.; Heil, S.; Haßlauer, I.; Schmidt, L.; von der Ohe, K.; Theuring, C.; Reinhard, A.; Schreier, P.; Beuerle, T. Pyrrolizidine alkaloids in pollen and pollen products. Mol. Nutr. Food Res. 2010, 54, 292–300. [Google Scholar] [CrossRef]
- Heil, M. Nectar: Generation, regulation and ecological functions. Trends Plant Sci. 2011, 16, 191–200. [Google Scholar] [CrossRef]
- Funk, V.A. Sesquiterpene lactones as toxonomis characters in the Asteraceae. Taxon 1983, 32, 1–27. [Google Scholar] [CrossRef]
- Scotti, M.T.; Emerenciano, V.; Ferreira, M.J.P.; Scitti, L.; Stefani, R.; da Silva, M.S.; Mendonça Junior, F.J.B. Self-organizing maps of molecular descriptors for sesquiterpene lactones and their application to the chemotaxonomy of the Asteraceae family. Molecules 2012, 17, 4684–4702. [Google Scholar] [CrossRef]
- Picman, A.K. Biological activities of sesquiterpene lactones. Biochem. Syst. Ecol. 1986, 14, 255–281. [Google Scholar] [CrossRef]
- Božičević, A.; De Mieri, M.; Nassenstein, C.; Wiegand, S.; Hamburger, M. Secondary Metabolites in Allergic Plant Pollen Samples Modulate Afferent Neurons and Murine Tracheal Rings. J. Nat. Prod. 2017, 80, 2953–2961. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Gonzalez, G.F.; dos Santos, F.A.; Da Costa, F.B. Sesquiterpene lactones: More than protective plant compounds with high toxicity. Crit. Rev. Plant Sci. 2016, 35, 18–37. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Patch, H.M.; Mortensen, D.A.; Tooker, J.F.; Grozinger, C.M. Macronutrient ratios in polln shape bumble bee (Bombus impatiens) foraging strategies and floral preferences. Proc. Natl. Acad. Sci. USA 2016, 113, E4035–E4042. [Google Scholar] [CrossRef] [Green Version]
- Vaudo, A.D.; Stabler, D.; Patch, H.M.; Tooker, J.F.; Grozinger, C.M.; Wright, G.A. Bumble bees regulate their intake of essential protein and lipid pollen macronutrients. J. Exp. Biol. 2016, 219, 3962–3970. [Google Scholar] [CrossRef] [Green Version]
- Vaudo, A.D.; Tooker, J.F.; Patch, H.M.; Biddiger, D.J.; Coccia, M.; Crone, M.K.; Fiely, M.; Francis, J.S.; Hines, H.M.; Hogdes, M.; et al. Pollen protein:lipid macronutrient ratios may guide broad patterns of bee species floral preferences. Insects 2020, 11, 132. [Google Scholar] [CrossRef] [Green Version]
- Ruedenauer, F.A.; Raubenheiner, D.; Kessner-Beierlein, D.; Grund-Mueller, N.; Noack, L.; Spaeteh, J.; Leonhardt, S.D. Best be(e) on low fat: Linking nutrient perception, regulation and fitness. Ecol. Lett. 2020, 3, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Behmer, S.T.; Nes, W.D. Insect sterol nutrition and physiology: A global overview. Adv. Insect Physiol. 2003, 31, 1–72. [Google Scholar]
- Cohen, A.C. Insect Diet: Science and Technology; CRC: Boca Raton, FL, USA, 2004. [Google Scholar]
- Meier-Melikyan, N.R.; Gabaraeva, N.I.; Polevova, S.V.; Grigor’eva, V.V.; Kosenko, Y.V.; Tekleva, M.V. Development of pollen grain walls and accumulation of sporopollenin. Russ. J. Plant Physiol. 2003, 50, 330–338. [Google Scholar] [CrossRef]
- Holm, L.; Doll, J.; Holm, E.; Pancho, J.; Herberger, J. World Weeds. Natural Histories and Distribution; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Cavieres, L.A.; Quiroz, C.L.; Molina-Montenegro, M.A.; Muñoz, A.A.; Pauchard, A. Nurse effect of the native cushion plant Azorella monantha on the invasive non-native Taraxacum officinale in the high-Andes of central Chile. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 217–226. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Quiroz, C.L.; Molina-Montenegro, M.A. Facilitation of the non-native Taraxacum officinale by native nurse cushion species in the high-Andes of central Chile: Are there differences between nurses? Funct. Ecol. 2008, 22, 148–156. [Google Scholar] [CrossRef]
- Jones, K.N. Do dandelion flowers influence seed set of a native plant (Delphinium nuttallianum) in subalpine meadows? Am. Midl. Nat. 2004, 151, 201–205. [Google Scholar] [CrossRef]
- Solbrig, O.; Simpson, B. Components of regulation of a population of dandelions in Michigan. J. Ecol. 1974, 62, 473–486. [Google Scholar] [CrossRef]
- Rasmont, P.; Coppée, A.; Michez, D.; De Meulemeester, T. An overview of the Bombus terrestris (L. 1758) subspecies (Hymenoptera: Apidae). Ann. Soc. Entomol. Fr. 2008, 44, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Leonhardt, S.D.; Blüthgen, N. The same, but different: Pollen foraging in honeybee and bumblebee colonies. Apidologie 2010, 43, 449–464. [Google Scholar] [CrossRef]
- Moerman, R.; Vanderplanck, M.; Fournier, D.; Jacquemart, A.-L.; Michez, D. Pollen nutrients better explain bumblebee colony development than pollen diversity. Insect Conserv. Diver. 2017, 10, 171–179. [Google Scholar] [CrossRef]
- Tasei, J.N.; Aupinel, P. Validation of a method using queenless Bombus terrestris micro-colonies for testing the nutritive value of commercial pollen mixes by comparison with queenright colonies. J. Econ. Entomol. 2008, 101, 1737–1742. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Harrison, X.A. Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ 2014, 2, e616. [Google Scholar] [CrossRef] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametricmodels. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 24 May 2017).
- Herbert, E.W.; Bickley, W.E.; Shimanuki, H. The brood-rearing capability of caged honey bees fed dandelion and mixed pollen diets. J. Econ. Entomol. 1970, 63, 215–218. [Google Scholar] [CrossRef]
- Loper, G.M.; Berdel, R.L. The effects of nine pollen diets on broodrearing of honeybees. Apidologie 1980, 11, 351–359. [Google Scholar] [CrossRef]
- Auclair, J.L.; Jamieson, C.A. A qualitative analysis of amino acids in pollen collected by bees. Science 1948, 108, 357–358. [Google Scholar] [CrossRef] [PubMed]
- Herbert, E.W. Honey Bee Nutrition. In The Hive and the Honey Bee; Graham, J.M., Ed.; Dadant & Sons: Hamilton, IL, USA, 1992; pp. 197–233. [Google Scholar]
- Arnold, S.E.J.; Peralta Idrovo, M.E.; Lomas Arias, L.J.; Belmain, S.R.; Stevenson, P.C. Herbivore defense compounds occur in pollen and reduce bumblebee colony fitness. J. Chem. Ecol. 2014, 40, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Irwin, R.E.; Cook, D.; Richardson, L.L.; Manson, J.S.; Gardner, D.R. Secondary compounds in floral rewards of toxic rangeland plants: Impacts on pollinators. J. Agr. Food Chem. 2014, 62, 7335–7344. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.S.; Nasr, M.E.; Marston, J.M.; Yuenzhen, F. The digestion of dandelion pollen by adult worker honeybees. Physiol. Entomol. 1985, 10, 75–82. [Google Scholar] [CrossRef]
- Brochu, K.K.; van Dyke, M.T.; Milano, N.J.; Petersen, J.D.; McArt, S.H.; Nault, B.A.; Kessler, A.; Danforth, B.N. Pollen defenses negatively impact foraging and fitness in a generalist bee (Bombus impatiens: Apidae). Sci. Rep. 2020, 10, 3112. [Google Scholar] [CrossRef] [Green Version]
- Ruedenauer, F.A.; Spaethe, J.; Leonhardt, S.D. How to know which food is good for you: Bumblebees use taste to discriminate between different concentrations of food differing in nutrient content. J. Exp. Biol. 2015, 218, 2233–2240. [Google Scholar] [CrossRef] [Green Version]
- Ruedenauer, F.A.; Leonhardt, A.D.; Lunau, K.; Spaethe, J. Bumblebees are able to perceive amino acids via chemotactile antennal stimulation. J. Comp. Physiol. A 2019, 2015, 321–331. [Google Scholar] [CrossRef]
- Eckhardt, M.; Haider, M.; Dorn, S.; Müller, A. Pollen mixing in pollen generalist solitary bees: A possible strategy to complement or mitigate unfavourable pollen properties? J. Anim. Ecol. 2014, 83, 588–597. [Google Scholar] [CrossRef]
- Vanderplanck, M.; Moerman, R.; Rasmont, P.; Lognay, G.; Wathelet, B.; Wattiez, R.; Michez, D. How does pollen chemistry impact development and feeding behavior of polylectic bees? PLoS ONE 2014, 9, e86209. [Google Scholar] [CrossRef] [PubMed]
- Champagne, D.; Bernays, E.A. Phytosterol unsuitability as a factor mediating food aversion learning in the grasshopper Schistocerca americana. Physiol. Entomol. 1991, 16, 391–400. [Google Scholar] [CrossRef]
- Behmer, S.T.; Elias, D.O. The nutritional significance of sterol metabolic constraints in the generalist grasshopper Schistocerca americana. J. Insect Physiol. 1999, 45, 339–348. [Google Scholar] [CrossRef]
- Behmer, S.T.; Elias, D.O. Phytosterol structure and its impact on feeding behavior in the generalist grasshopper Schistocerca americana. Physiol. Entomol. 1999, 24, 18–27. [Google Scholar] [CrossRef]
- Behmer, S.T.; Elias, D.O.; Bernays, E.A. Post-ingestive feedbacks and associative learning regulate the intake of unsuitable sterols in a generalist grasshopper. J. Exp. Biol. 1999, 202, 739–748. [Google Scholar]
- Ritter, K.S.; Nes, W.R. The effects of the structure of sterols on the development of Heliothis zea. J. Insect Physiol. 1981, 27, 419–424. [Google Scholar] [CrossRef]
- Behmer, S.T.; Grebenok, R.J. Impact of sterols on life-history traits of a caterpillar. Physiol. Entomol. 1998, 23, 165–175. [Google Scholar] [CrossRef]
- Engel, P.; Martinson, V.G.; Moran, N.A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef] [Green Version]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [Green Version]
- Koch, H.; Schmid-Hempel, P. Gut microbiota instead of host genotype drive the specificity in the interaction of a natural host-parasite system. Ecol. Lett. 2012, 15, 1095–1103. [Google Scholar] [CrossRef]
- Moran, N.A. Genomics of the honey bee microbiome. Curr. Opin. Insect Sci. 2015, 10, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segers, F.H.I.D.; Kešnerová, L.; Kosoy, M.; Engel, P. Genomic changes associated with the evolutionary transition of an insect gut symbiont into a blood-borne pathogen. ISME J. 2017, 11, 1232–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billiet, A.; Meeus, I.; Van Nieuwerburgh, F.; Deforce, D.; Wäckers, F.; Smagghe, G. Impact of sugar syrup and pollen diet on the bacterial diversity in the gut of indoor-reared bumblebees (Bombus terrestris). Apidologie 2015, 47, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Raubenheimer, D.; Simpson, S.J. The geometry of compensatory feeding in the locust. Anim. Behav. 1993, 45, 953–964. [Google Scholar] [CrossRef] [Green Version]
- Pirk, C.W.W.; Boodhoo, C.; Human, H.; Nicolson, S. The importance of protein type and protein to carbohydrate ratio for survival and ovarian activation of caged honeybees (Apis mellifera scutellata). Apidologie 2010, 41, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Archer, C.R.; Pirk, C.W.W.; Wright, G.A.; Nicolson, S.W. Nutrition affects survival in African honeybees exposed to interacting stressors. Funct. Ecol. 2014, 28, 913–923. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Digestive Damages | Treatment | ||||
|---|---|---|---|---|---|
| Control | Natural Taraxacum | Crushed Taraxacum | Phytosterol | Lactone | |
| Cytoplasmic blebbing | X | ||||
| Cytoplasm vacuolization | X | X | |||
| Disorganization of the brush-like border | X | ||||
| Hydropic degeneration | X | ||||
| Interstitial edema | X | ||||
| Necrotic cells | X | X | |||
| Pyknotic nuclei | X | X | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanderplanck, M.; Gilles, H.; Nonclercq, D.; Duez, P.; Gerbaux, P. Asteraceae Paradox: Chemical and Mechanical Protection of Taraxacum Pollen. Insects 2020, 11, 304. https://doi.org/10.3390/insects11050304
Vanderplanck M, Gilles H, Nonclercq D, Duez P, Gerbaux P. Asteraceae Paradox: Chemical and Mechanical Protection of Taraxacum Pollen. Insects. 2020; 11(5):304. https://doi.org/10.3390/insects11050304
Chicago/Turabian StyleVanderplanck, Maryse, Hélène Gilles, Denis Nonclercq, Pierre Duez, and Pascal Gerbaux. 2020. "Asteraceae Paradox: Chemical and Mechanical Protection of Taraxacum Pollen" Insects 11, no. 5: 304. https://doi.org/10.3390/insects11050304
APA StyleVanderplanck, M., Gilles, H., Nonclercq, D., Duez, P., & Gerbaux, P. (2020). Asteraceae Paradox: Chemical and Mechanical Protection of Taraxacum Pollen. Insects, 11(5), 304. https://doi.org/10.3390/insects11050304