Climate Change May Restrict the Predation Efficiency of Mesocyclops aspericornis (Copepoda: Cyclopidae) on Aedes aegypti (Diptera: Culicidae) Larvae


Abstract
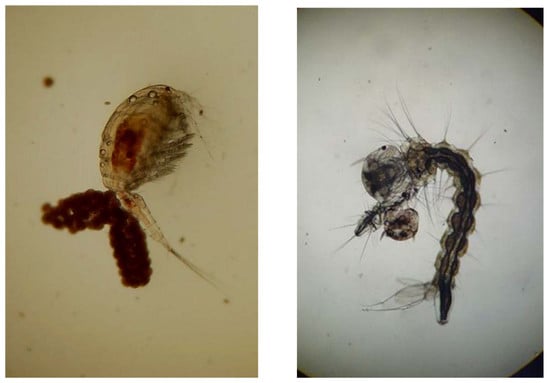
:
1. Introduction
2. Materials and Methods
2.1. Copepods
2.2. Mosquito Colonies
2.3. Experimental Design
2.4. Rational of Experimental Settings
2.5. Statistical Analyses
3. Results
3.1. Effects of Copepod Presence on the Larval Performance of Ae. aegypti
3.2. Effects of Temperature, Nutrition Conditions and Larval Density of Ae. aegypti on Copepod Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Erlanger, T.E.; Keiser, J.; Utzinger, J. Effect of dengue vector control interventions on entomological parameters in developing countries: A systematic review and meta-analysis. Med. Veter-Entomol. 2008, 22, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Kay, B.H.; Nam, V.S. New strategy against Aedes aegypti in Vietnam. Lancet 2005, 365, 613–617. [Google Scholar] [CrossRef]
- Kay, B.H.; Hanh, T.T.T.; Le, N.H.; Quy, T.M.; Nam, V.S.; Hang, P.V.D.; Yen, N.T.; Hill, P.S.; Vos, T.; Ryan, P.A. Sustainability and Cost of a Community-Based Strategy Against Aedes aegypti in Northern and Central Vietnam. Am. J. Trop. Med. Hyg. 2010, 82, 822–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, V.S.; Reid, J.W.; Yen, N.T.; Kay, B.H.; Holynska, M. National progress in dengue vector control in Vietnam: Survey for Mesocyclops (Copepoda), Micronecta (Corixidae), and fish as biological control agents. Am. J. Trop. Med. Hyg. 2000, 62, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, V.S.; Yen, N.T.; Duc, H.M.; Tu, T.C.; Thang, V.T.; Le, N.H.; San, L.H.; Le Loan, L.; Huong, V.T.Q.; Khanh, L.H.K.; et al. Community-based control of Aedes aegypti by using Mesocyclops in southern Vietnam. Am. J. Trop. Med. Hyg. 2012, 86, 850–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marten, G.G. Elimination of Aedes albopictus from tire piles by introducing Macrocyclops albidus (Copepoda, Cyclopidae). J. Am. Mosq. Control. Assoc. 1990, 6, 689–693. [Google Scholar] [PubMed]
- Marten, G.G.; Reid, J.W. CYCLOPOID COPEPODS. J. Am. Mosq. Control. Assoc. 2007, 23, 65–92. [Google Scholar] [CrossRef]
- Früh, L.; Kampen, H.; Schaub, G.A.; Werner, D. Predation on the invasive mosquito Aedes japonicus (Diptera: Culicidae) by native copepod species in Germany. J. Vector Ecol. 2019, 44, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Udayanga, L.; Ranathunge, T.; Iqbal, M.C.M.; Abeyewickreme, W.; Hapugoda, M. Predatory efficacy of five locally available copepods on Aedes larvae under laboratory settings: An approach towards bio-control of dengue in Sri Lanka. PLoS ONE 2019, 14, e0216140. [Google Scholar] [CrossRef] [Green Version]
- Vu, S.N.; Nguyen, T.Y.; Tran, V.P.; Truong, U.N.; Le, Q.M.; Le, V.L.; Le, T.N.; Bektas, A.; Briscombe, A.; Aaskov, J.G.; et al. Elimination of dengue by community programs using Mesocyclops(Copepoda) against Aedes aegypti in central Vietnam. Am. J. Trop. Med. Hyg. 2005, 72, 67–73. [Google Scholar]
- Tran, T.T.; Olsen, A.; Viennet, E.; Sleigh, A.C. Social sustainability of Mesocyclops biological control for dengue in South Vietnam. Acta Trop. 2015, 141, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, A.; Han, W.W.; George, L.; Velayudhan, R.; Toledo, J.; Ranzinger, S.R.; Horstick, O.; Manrique-Saide, P. Community effectiveness of copepods for dengue vector control: Systematic review. Trop. Med. Int. Heal. 2015, 20, 685–706. [Google Scholar] [CrossRef] [PubMed]
- Neale, J.T.; Juliano, S.A. Finding the sweet spot: What levels of larval mortality lead to compensation or overcompensation in adult production? Ecosphere 2019, 10, 02855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soumare, M.K.F.; E Cilek, J.; Schreibers, E.T. Prey and size preference of Mesocyclops longisetus (Copepoda) for Aedes albopictus and Culex quinquefasciatus larvae. J. Am. Mosq. Control. Assoc. 2004, 20, 305–310. [Google Scholar]
- Dieng, H.; Boots, M.; Tuno, N.; Tsuda, Y.; Takagi, M. Life history effects of prey choice by copepods: Implications for biocontrol of vector mosquitoes. J. Am. Mosq. Control. Assoc. 2003, 19, 67–73. [Google Scholar]
- Suarez, M.F.; Marten, G.G.; Clark, G.G. A simple method for cultivating freshwater copepods used in biological control of Aedes aegypti. J. Am. Mosq. Control. Assoc. 1992, 8, 409–412. [Google Scholar]
- Dussart, B.H.; Defaye, D. Introduction to the Copepoda; SPD Academic: Amsterdam, The Netherlands, 1995; p. 277. [Google Scholar]
- Barrera, R.; Amador, M.; Clark, G.G. Ecological factors influencing Aedes aegypti (Diptera: Culicidae) productivity in artificial containers in Salinas, Puerto Rico. J. Med Entomol. 2006, 43, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Gama, R.A.; Alves, K.D.C.; Martins, R.F.; Eiras, A.E.; De Resende, M.C. [Effect of larvae density on adult size of Aedes aegypti reared under laboratory condictions]. Rev. Soc. Bras. Med. Trop. 2005, 38, 64–66. [Google Scholar] [CrossRef] [Green Version]
- Hopp, U.; Maier, G.; Bleher, R. Reproduction and adult longevity of five species of planktonic cyclopoid copepods reared on different diets: A comparative study. Freshw. Boil. 1997, 38, 289–300. [Google Scholar] [CrossRef]
- Campbell, L.P.; Luther, C.; Moo-Llanes, D.; Ramsey, J.M.; Danis-Lozano, R.; Peterson, A.T. Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos. Trans. R. Soc. B 2015, 370, 20140135. [Google Scholar] [CrossRef]
- Farjana, T.; Tuno, N.; Higa, Y. Effects of temperature and diet on development and interspecies competition in Aedes aegypti and Aedes albopictus. Med Veter-Entomol. 2011, 26, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Armbruster, P.; A Hutchinson, R. Pupal mass and wing length as indicators of fecundity in Aedes albopictus and Aedes geniculatus (Diptera: Culicidae). J. Med. Entomol. 2002, 39, 699–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivière, F.; Thirel, R. La prèdation du cope’pode Mesocyclops leuckarti pilosa (Crustacea) sur les larves de Aedes (Stegomyia) aegypti et Ae. (St.) polynesiensis (Dip.: Culicidae): Essais prèliminaires d’utilisation comme agent de lutte biologique. Entomophaga 1981, 26, 427–439. [Google Scholar] [CrossRef]
- Jennings, C.D.; Phommasack, B.; Sourignadeth, B.; Kay, B.H. Aedes aegypti control in the Lao People’s Democratic Republic, with reference to copepods. Am. J. Trop. Med. Hyg. 1995, 53, 324–330. [Google Scholar] [CrossRef]
- Wang, S.; Phong, T.V.; Tuno, N.; Kawada, H.; Takagi, M. Sensitivity of the larvivorous copepod species, Mesocyclops pehpeiensis and Megacyclops viridis, to the insect growth regulator, pyriproxyfen. J. Am. Mosq. Control. Assoc. 2005, 21, 483–488. [Google Scholar] [CrossRef]
- Murugan, K.; Benelli, G.; Panneerselvam, C.; Subramaniam, J.; Jeyalalitha, T.; Dinesh, D.; Nicoletti, M.; Hwang, J.-S.; Suresh, U.; Madhiyazhagan, P. Cymbopogon citratus-synthesized gold nanoparticles boost the predation efficiency of copepod Mesocyclops aspericornis against malaria and dengue mosquitoes. Exp. Parasitol. 2015, 153, 129–138. [Google Scholar] [CrossRef]
- Yakob, L.; Funk, S.; Camacho, A.; Brady, O.; Edmunds, W.J. Aedes aegypti Control Through Modernized, Integrated Vector Management. PLoS Curr. 2017, 9. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Parameter | Response Variables of Ae. aegypti | |||||||
|---|---|---|---|---|---|---|---|---|
| Mortality | Larval Developing Duration | pupal Dry Weight | ||||||
| First Instar Period | Whole Larval Period | |||||||
| Likelihood Ratio χ2 | p | Likelihood Ratio χ2 | p | Likelihood Ratio χ2 | p | Likelihood Ratio χ2 | p | |
| Copepod | 1630 | <0.0001 | 63 | <0.0001 | 76 | <0.0001 | 8 | 0.0147 |
| Temperature | 31 | <0.0001 | 14 | 0.0002 | 228 | <0.0001 | 29 | <0.0001 |
| Larval density | 44 | <0.0001 | 19 | <0.0001 | 173 | <0.0001 | 9 | 0.0034 |
| Nutrition | 33 | <0.0001 | 30 | <0.0001 | 122 | <0.0001 | 17 | <0.0001 |
| Cope ×Temp | 50 | <0.0001 | 0 | NS | 14 | 0.0008 | 1 | NS |
| Cope × Larv | 63 | <0.0001 | 21 | <0.0001 | 15 | 0.0001 | 18 | <0.0001 |
| Cope × Nutr | 26 | <0.0001 | 1 | NS | 26 | <0.0001 | 0 | NS |
| Temp × Larv | 2 | NS | 0 | NS | 12 | 0.0023 | 26 | <0.0001 |
| Temp × Nutr | 5 | 0.024 | 2 | NS | 6 | 0.0449 | 12 | 0.0024 |
| Larv × Nutr | 0 | NS | 2 | NS | 0 | NS | 16 | <0.0001 |
| Parameter | Response Variables of M. aspericornis | |||
|---|---|---|---|---|
| Number of Offspring | Developmental Duration | |||
| Likelihood Ratio χ2 | p | Likelihood Ratio χ2 | p | |
| Temperature | 19 | <0.0001 | 130 | <0.0001 |
| Larval density | 23 | <0.0001 | 236 | <0.0001 |
| Nutrition | 0 | NS | 100 | <0.0001 |
| Temp × Larv | 17 | 0.0002 | 120 | <0.0001 |
| Temp × Nutr | 3 | NS | 77 | <0.0001 |
| Larv × Nutr | 0 | NS | 47 | <0.0001 |
| Food | 10 Larvae | 50 Larvae | ||||
|---|---|---|---|---|---|---|
| 20 °C | 27 °C | 32 °C | 20 °C | 27 °C | 32 °C | |
| Copepod offspring abundance | ||||||
| Rich | 15.7 ± 9.0 BCD | 79.9 ± 34.6 A | 70.7 ± 33.7 A | 9.6 ± 8.5 CD | 16.7 ± 8.6 BCD | 41.8 ± 18.3 B |
| Poor | 12.4 ± 4.5 BCD | 29.5 ± 11.4 BC | 45.9 ± 15.8 ABC | 4.5 ± 2.3 D | 7.9 ± 6.5 CD | 8.8 ± 4.8 CD |
| Developmental time (day) | ||||||
| Rich | 15.8 ± 1.4 de | 9.4 ± 1.1 g | 5.7 ±1.0 h | 21.9 ± 3.5 b | 15.1 ± 3.4 ef | 8.5 ± 1.4 g |
| Poor | 18.9 ± 1.7 c | 14.1 ± 3.2 ef | 9.2 ± 2.4 g | 52.3 ± 11.2 a | 18.2 ± 2.7 cd | 12.7 ± 2.2 f |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuno, N.; Phong, T.V.; Takagi, M. Climate Change May Restrict the Predation Efficiency of Mesocyclops aspericornis (Copepoda: Cyclopidae) on Aedes aegypti (Diptera: Culicidae) Larvae. Insects 2020, 11, 307. https://doi.org/10.3390/insects11050307
Tuno N, Phong TV, Takagi M. Climate Change May Restrict the Predation Efficiency of Mesocyclops aspericornis (Copepoda: Cyclopidae) on Aedes aegypti (Diptera: Culicidae) Larvae. Insects. 2020; 11(5):307. https://doi.org/10.3390/insects11050307
Chicago/Turabian StyleTuno, Nobuko, Tran Vu Phong, and Masahiro Takagi. 2020. "Climate Change May Restrict the Predation Efficiency of Mesocyclops aspericornis (Copepoda: Cyclopidae) on Aedes aegypti (Diptera: Culicidae) Larvae" Insects 11, no. 5: 307. https://doi.org/10.3390/insects11050307
APA StyleTuno, N., Phong, T. V., & Takagi, M. (2020). Climate Change May Restrict the Predation Efficiency of Mesocyclops aspericornis (Copepoda: Cyclopidae) on Aedes aegypti (Diptera: Culicidae) Larvae. Insects, 11(5), 307. https://doi.org/10.3390/insects11050307