Potential Distribution and Niche Differentiation of Spodoptera frugiperda in Africa


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
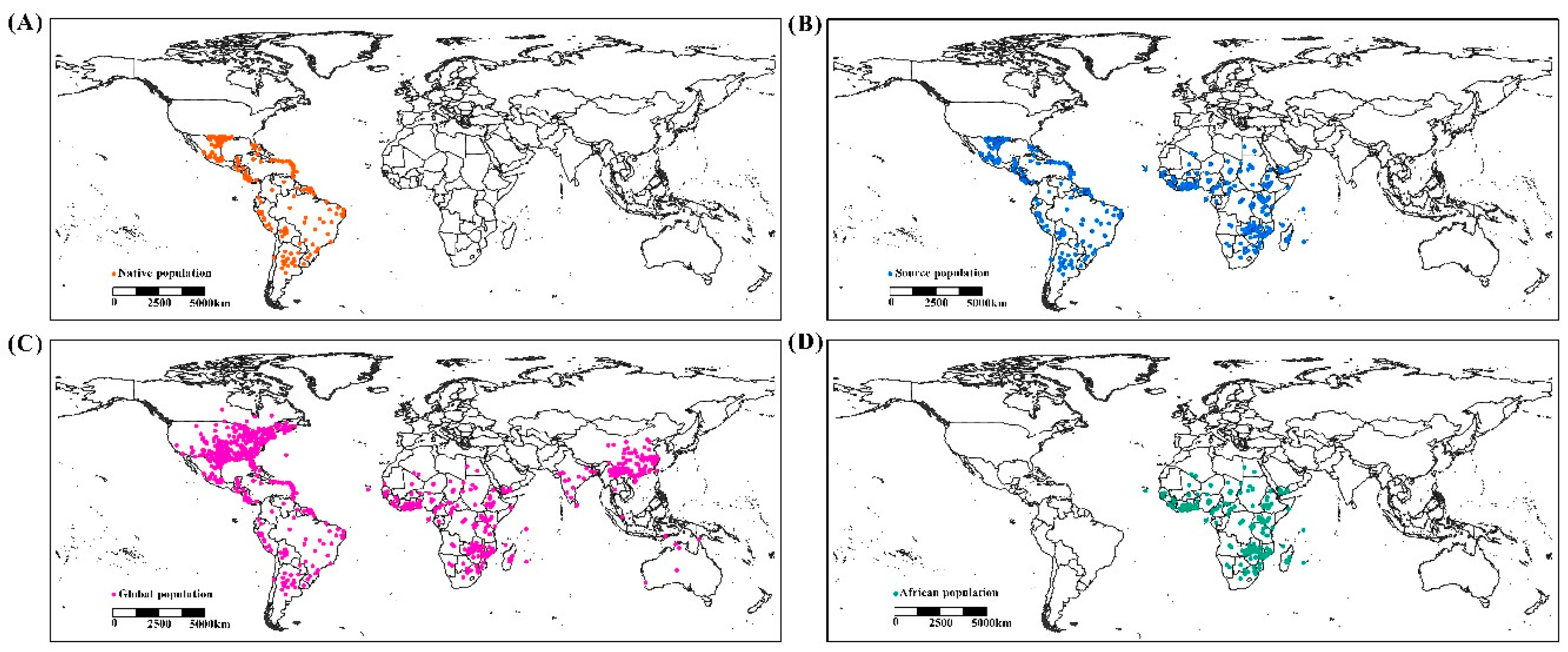
2.1. Occurrence Data
2.2. Environmental Variables
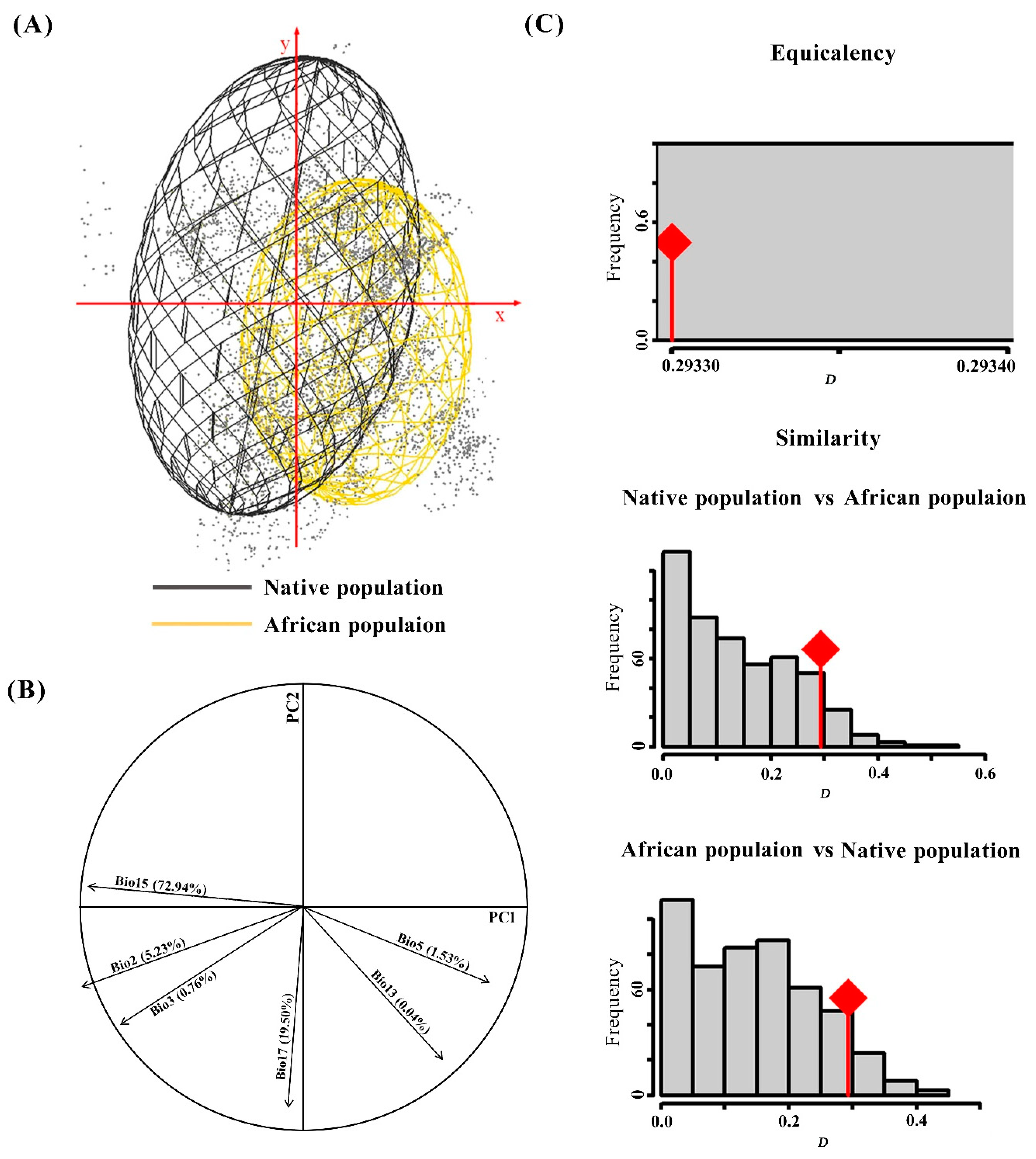
2.3. Climate Space Comparison
2.4. Model Calibration
3. Results
3.1. Climate Spaces
3.2. Model Predictions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, C.Y.; Zhang, W.J.; Lu, B.R.; Chen, J.K. Progress in studies on mechanisms of biological invasion. Biodivers. Sci. 2001, 9, 430–438. [Google Scholar]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldtman, R.; Lado, T.F.; Botes, A.; Procheş, Ş.; Timm, A.E.; Geertsema, H.; Chown, S.L. Creating novel food weds on introduced Australian acacias: Indirect effects of galling biological control agents. Divers. Distrib. 2011, 17, 958–967. [Google Scholar] [CrossRef]
- Anthony, S.; Sardain, E.; Brian, L. Global forecasts of shipping traffic and biological invasions to 2050. Nat. Sustain. 2019, 2, 274–282. [Google Scholar]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Todd, E.L.; Poole, R.W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere. Ann. Entomol. Soc. Am. 1980, 73, 722–738. [Google Scholar] [CrossRef]
- Sparks, A.N. Fall armyworm symposium: A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Nagoshi, R.N. Improvements in the identification of strains facilitate population studies of fall armyworm subgroups. Ann. Entomol. Soc. Am. 2012, 105, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Early, R.; Gonzalez-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. Global Monitoring for Fall Armyworm Control. 2020. Available online: http://www.fao.org/fall-armyworm/monitoring-tools/faw-map/en/ (accessed on 3 June 2020).
- Lima, M.S.; Silva, P.S.L.; Oliveira, O.F.; Silva, K.M.B.; Freitas, F.C.L. Corn yield response to weed and fall armyworm controls. Planta Daninha 2010, 28, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.E.; Hellin, J.; Sonder, K.; Araus, J.L.; MacRobert, J.F.; Thierfelder, C.; Prasanna, B.M. Adapting maize production to climate change in sub-Saharan Africa. Food Secur. 2013, 5, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, F.; Rieckmann, U.; Winter, S. The spread of the fall armyworm Spodoptera frugiperda in Africa—What should be done next? J. Plant Dis. Prot. 2019, 126, 97–101. [Google Scholar] [CrossRef]
- Göergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS ONE 2016, 11, e165632. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N. Evidence that a major subpopulation of fall armyworm found in the Western Hemisphere is rare or absent in Africa, which may limit the range of crops at risk of infestation. PLoS ONE 2019, 14, e0208966. [Google Scholar] [CrossRef] [Green Version]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Center for Agriculture and Bioscience International. Datasheet. Spodoptera frugiperda (fall armyworm). Invasive Species Compendium 2016. Available online: http://www.cabi.orglisc/datasheet129810 (accessed on 5 June 2020).
- Zhu, G.P.; Liu, G.Q.; Bu, W.J.; Gao, Y.B. Ecological niche modeling and its applications in biodiversity conservation. Biodivers. Sci. 2013, 21, 90–98. [Google Scholar]
- Escobar, L.E.; Lira-Noriega, A.; Medina-Vogel, G.; Peterson, A.T. Potential for spread of the white-Nose fungus (Pseudogymnoascus destructans) in the Americas: Use of MaxEnt and NicheA to assure strict model transference. Geospat. Health 2014, 9, 221–229. [Google Scholar] [CrossRef]
- Schulte, U.; Hochkirch, A.; Lötters, S.; Rödder, D.; Schweiger, S.; Weimann, T.; Veith, M. Cryptic niche conservatism among evolutionary lineages of an invasive lizard. Glob. Ecol. Biogeogr. 2012, 21, 198–211. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strubbe, D.; Beauchard, O.; Matthysen, E. Niche conservatism among non-native vertebrates in Europe and North America. Ecography 2015, 38, 321–329. [Google Scholar] [CrossRef]
- Kenis, M.; Plessis, H.; Berg, J.V.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a candidate parasitoid for the biological control of Spodoptera frugiperda in Africa, is already present on the continent. Insects 2019, 10, 92. [Google Scholar]
- Sisay, B.; Simiyu, J.; Mendesil, E.; Likhayo, P.; Ayalew, G.; Mohamed, S.; Subramanian, S.; Tefera, T. Fall armyworm, Spodoptera frugiperda infestations in East Africa: Assessment of damage and parasitism. Insects 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Qiao, H.J.; Peterson, A.T.; Campbell, L.P.; Soberón, J.; Ji, L.Q.; Escobar, L.E. NicheA: Creating virtual species and ecological niches in multivariate environmental scenarios. Ecography 2016, 39, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Potter, J.; Pio, D.; Dubuis, A.; et al. Ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2011, 14, 484–492. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Fitzpatrickm, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A. Insights into the area under the receiver operating characteristic curve (AUC) as a discrimination measure in species distribution modelling. Glob. Ecol. Biogeogr. 2012, 21, 498–507. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberón, J. Species distribution modeling and ecological niche modeling: Getting the concepts right. Natureza Conservação 2012, 10, 1–6. [Google Scholar] [CrossRef]
- Muhammad, N.B.; Fan, J.; Haseeb, M.; Zhang, R. Mapping potential distribution of Spodoptera frugiperda (Lepidoptera: Noctuidae) in central Asia. Insects 2020, 11, 172. [Google Scholar]
- Tingley, R.; Thompson, M.B.; Hartley, S.; Chapple, D.G. Patterns of niche filling and expansion across the invaded ranges of an Australian lizard. Ecography 2016, 39, 270–280. [Google Scholar] [CrossRef]
- Peterson, A.T. Ecological niche conservatism: A time structured review of evidence. J. Biogeogr. 2011, 38, 817–828. [Google Scholar] [CrossRef]
- Václavík, T.; Meentemeyer, R.K. Equilibrium or not? Modelling potential distribution of invasive species in different stages of invasion. Divers. Distrib. 2012, 18, 73–83. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Validation | Model | Mean AUC 1 at 0.95 | Mean AUC at 0.5 | Ratio 2 |
|---|---|---|---|---|
| Interpolative validation | Native model | 0.70 | 0.50 | 1.40 |
| Source model | 0.83 | 0.50 | 1.66 | |
| Global model | 0.85 | 0.50 | 1.71 | |
| Africa model | 0.64 | 0.50 | 1.28 | |
| Extrapolative validation | Native model to Africa | 0.61 | 0.50 | 1.22 |
| Source model to Africa | 0.57 | 0.50 | 1.14 | |
| Africa model to native 3 | 0.73 | 0.50 | 1.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Wu, P.; Tian, T.; Ren, Q.; Haseeb, M.; Zhang, R. Potential Distribution and Niche Differentiation of Spodoptera frugiperda in Africa. Insects 2020, 11, 383. https://doi.org/10.3390/insects11060383
Fan J, Wu P, Tian T, Ren Q, Haseeb M, Zhang R. Potential Distribution and Niche Differentiation of Spodoptera frugiperda in Africa. Insects. 2020; 11(6):383. https://doi.org/10.3390/insects11060383
Chicago/Turabian StyleFan, Jingyu, Pengxiang Wu, Tianqi Tian, Qilin Ren, Muhammad Haseeb, and Runzhi Zhang. 2020. "Potential Distribution and Niche Differentiation of Spodoptera frugiperda in Africa" Insects 11, no. 6: 383. https://doi.org/10.3390/insects11060383
APA StyleFan, J., Wu, P., Tian, T., Ren, Q., Haseeb, M., & Zhang, R. (2020). Potential Distribution and Niche Differentiation of Spodoptera frugiperda in Africa. Insects, 11(6), 383. https://doi.org/10.3390/insects11060383