Long-Term Evaluation of Breeding Scheme Alternatives for Endangered Honeybee Subspecies


Abstract
:1. Introduction
2. Materials and Methods
2.1. Breeding Scheme
2.2. Genetic Models
2.3. Breeding Value Estimation
2.4. Analysis of Simulation Output
3. Results
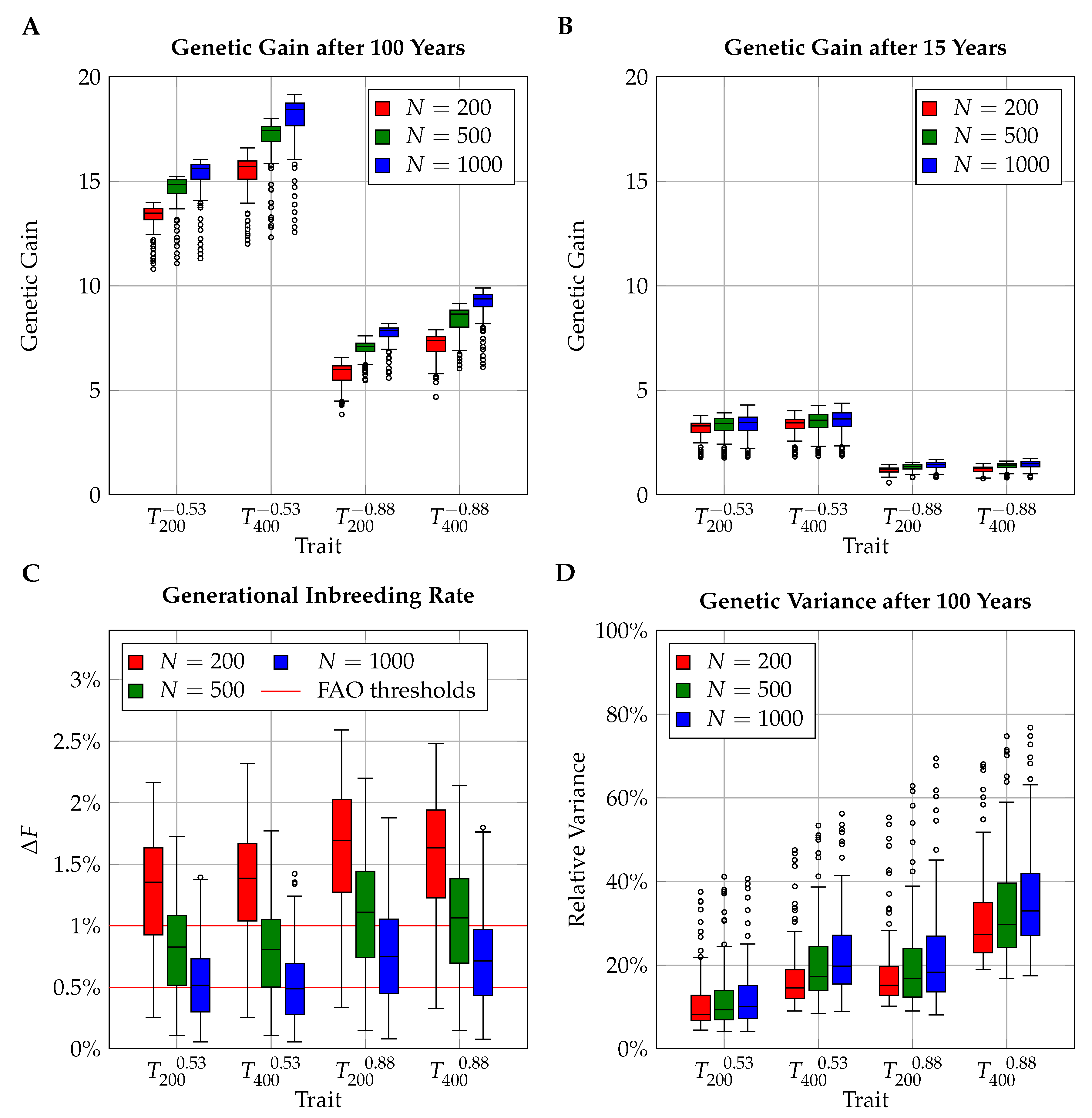
3.1. Overview of the Simulation Outcome
3.2. Comparison of Breeding Schemes
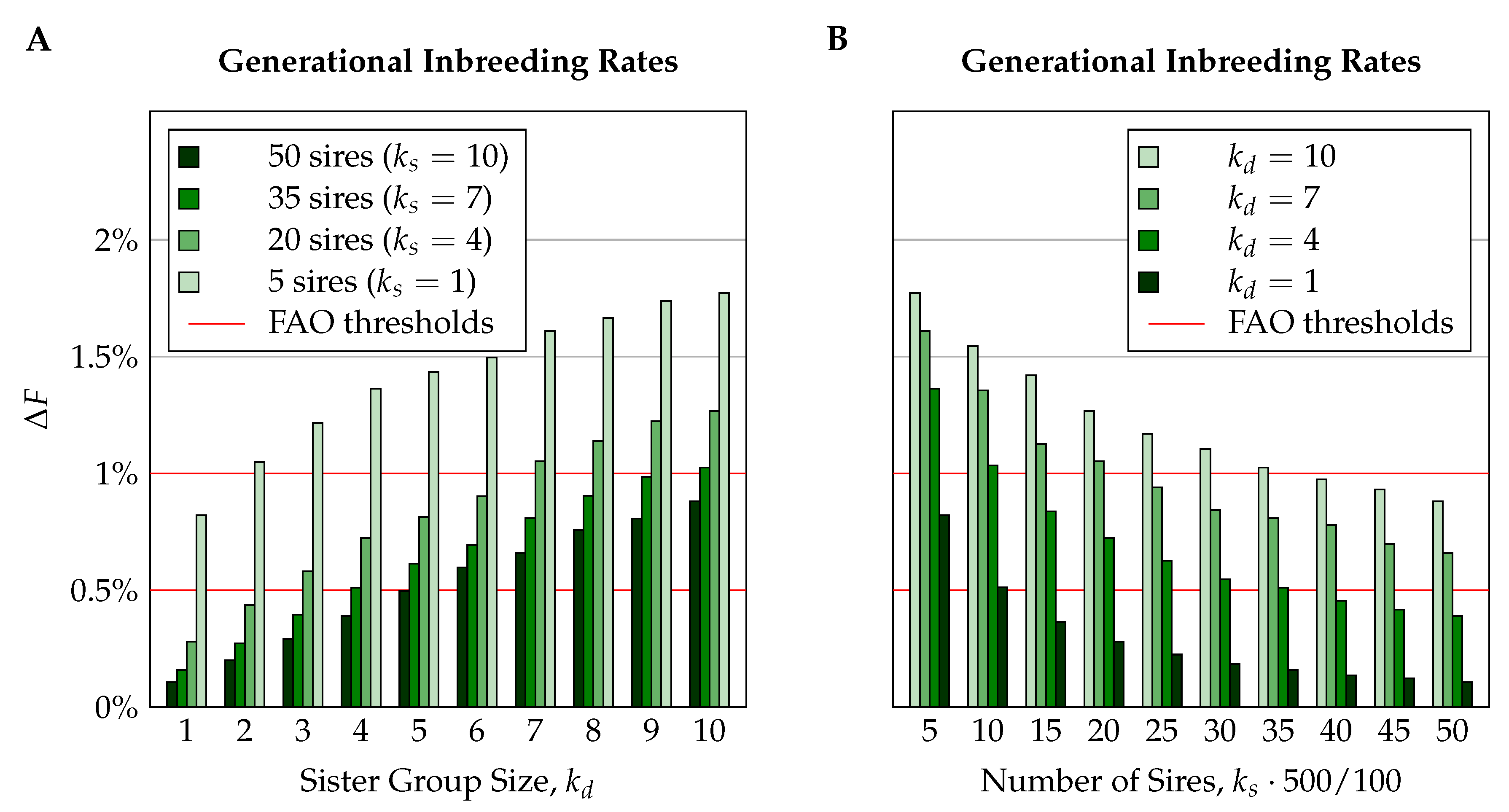
3.2.1. Generational Inbreeding Rates
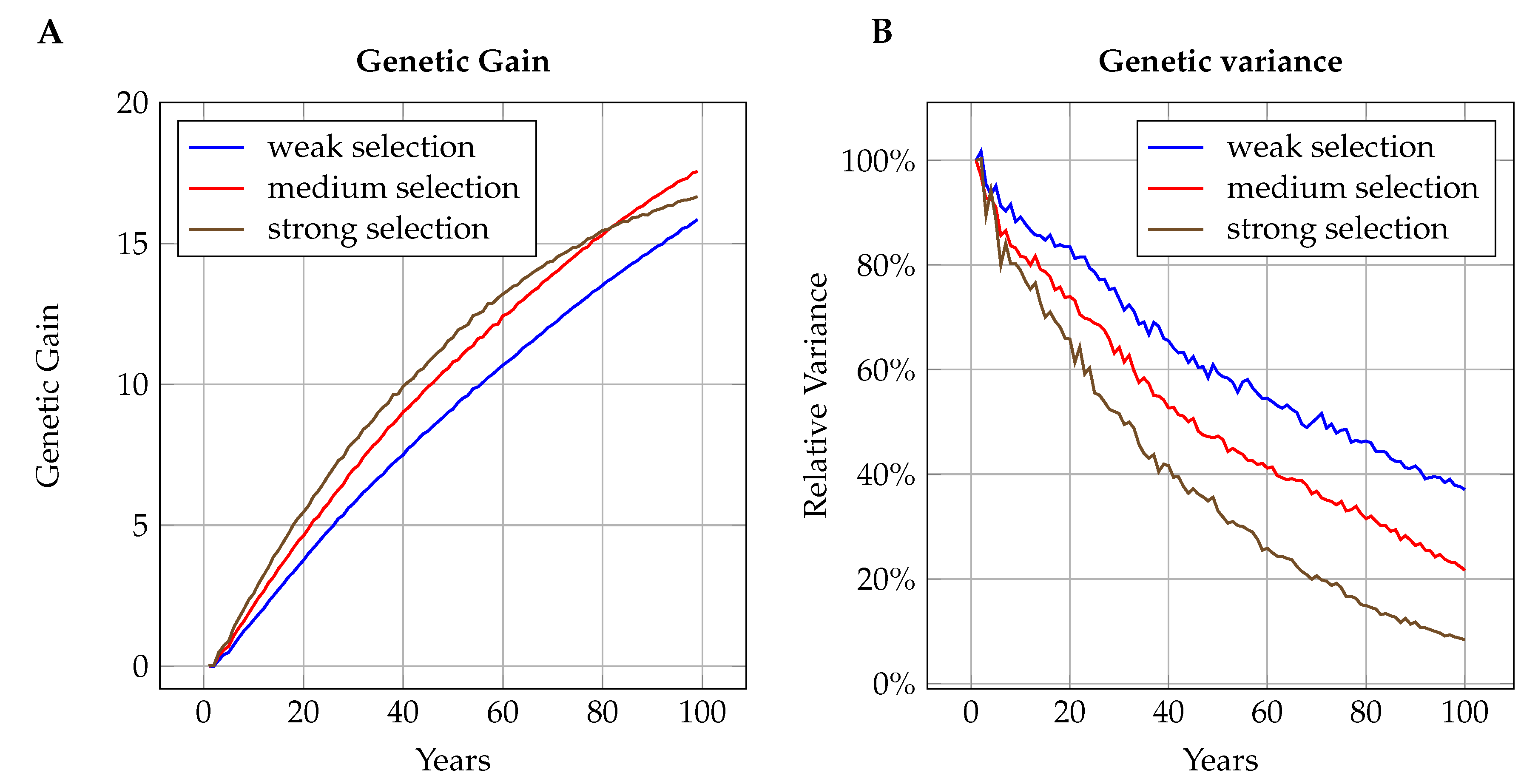
3.2.2. Sustainable Genetic Gain
3.3. Short-Term Genetic Gain
4. Discussion
4.1. Reality Check on Assumptions
4.1.1. Population Structure
4.1.2. Genetic Model
4.1.3. Analysis of Results
4.2. Influences of Simulation Parameters
4.2.1. Population Size
4.2.2. Genetic Parameters
4.2.3. Sister Group Sizes
4.2.4. Number of Sires
4.3. Further Breeding Schemes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BLUP | Best Linear Unbiased Prediction |
| CBS | Competitive Breeding Scheme |
| FAO | Food and Agriculture Organization of the United Nations |
| TBV | True Breeding Value |
References
- Brascamp, E.W.; Bijma, P. Methods to estimate breeding values in honey bees. Genet. Sel. Evol. 2014, 46, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, R.; Plate, M.; Hoppe, A.; Bienefeld, K. Computing inbreeding coefficients and the inverse numerator relationship matrix in large populations of honey bees. J. Anim. Breed. Genet. 2018, 135, 323–332. [Google Scholar] [CrossRef]
- Plate, M.; Bernstein, R.; Hoppe, A.; Bienefeld, K. Comparison of infinitesimal and finite locus models for long-term breeding simulations with direct and maternal effects at the example of honeybees. PLoS ONE 2019, 14, e0213270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büchler, R.; Berg, S.; Le Conte, Y. Breeding for resistance to Varroa destructor in Europe. Apidologie 2010, 41, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Büchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Hatjina, F.; Kezic, N.; Kryger, P.; Spivak, M.; Uzunov, A.; Wilde, J. Standard methods for rearing and selection of Apis mellifera queens. J. Apicult. Res. 2013, 52, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Uzunov, A.; Büchler, R.; Bienefeld, K. Performance Testing Protocol. A Guide for European Honey Bee Breeders; Version 1.0; Landesbetrieb Landwirtschaft Hessen: Kirchhain, Germany, 2015. [Google Scholar]
- Uzunov, A.; Brascamp, E.W.; Büchler, R. The basic concept of honey bee breeding programs. Bee World 2017, 94, 84–87. [Google Scholar] [CrossRef]
- Cauia, E.; Siceanu, A.; Patruica, S.; Bura, M.; Sapcaliu, A.; Magdici, M. The standardization of the honeybee colonies evaluation methodology, with application in honeybee breeding programs, in Romanian conditions. Sci. Pap. Anim. Sci. Biotechnol. 2010, 43, 174–180. [Google Scholar]
- Hatjina, F. Greek honey bee queen quality certification. Bee World 2012, 89, 18–20. [Google Scholar] [CrossRef]
- Uzunov, A.; Meixner, M.; Büchler, R.; Galea, T. Conservation of the endemic Maltese honey bee (Apis mellifera ruttneri). SmartBees Newsletter 2018, 5. Available online: www.smartbees-fp7.eu/Extension/newsletter-5/#ancor-no2 (accessed on 29 June 2020).
- Sheppard, W.S.; Arias, M.C.; Grech, A.; Meixner, M.D. Apis mellifera ruttneri, a new honey bee subspecies from Malta. Apidologie 1997, 28, 287–293. [Google Scholar] [CrossRef] [Green Version]
- De La Rúa, P.; Jaffé, R.; Dall’Olio, R.; Muñoz, I.; Serrano, J. Biodiversity, conservation and current threats to European honey bees. Apidologie 2009, 40, 263–284. [Google Scholar] [CrossRef] [Green Version]
- Theisen-Jones, H.; Bienefeld, K. The Asian honey bee (Apis cerana) is significantly in decline. Bee World 2016, 93, 90–97. [Google Scholar] [CrossRef]
- Schouten, C.; Lloyd, D.; Lloyd, H. Beekeeping with the Asian Honey Bee (Apis cerana javana Fabr) in the Indonesian islands of Java, Bali, Nusa Penida, and Sumbawa. Bee World 2019, 96, 45–49. [Google Scholar] [CrossRef]
- Vung, N.N.; Choi, Y.S.; Kim, I. High resistance to Sacbrood virus disease in Apis cerana (Hymenoptera: Apidae) colonies selected for superior brood viability and hygienic behavior. Apidologie 2020, 51, 61–74. [Google Scholar] [CrossRef]
- Kosgey, I.S.; Baker, R.L.; Udo, H.M.J.; Van Arendonk, J.A.M. Successes and failures of small ruminant breeding programmes in the tropics: A review. Small Rumin. Res. 2006, 61, 13–28. [Google Scholar] [CrossRef]
- Lauvie, A.; Audiot, A.; Couix, N.; Casabianca, F.; Brives, H.; Verrier, E. Diversity of rare breed management programs: Between conservation and development. Livest. Sci. 2011, 140, 161–170. [Google Scholar] [CrossRef]
- Cervantes, I.; Gutiérrez, J.P.; Meuwissen, T.H.E. Response to selection while maximizing genetic variance in small populations. Genet. Sel. Evol. 2016, 48, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meuwissen, T.H.E.; Woolliams, J.A. Effective sizes of livestock populations to prevent a decline in fitness. Theor. Appl. Genet. 1994, 89, 1019–1026. [Google Scholar] [CrossRef]
- Hill, W.G. Maintenance of quantitative genetic variation in animal breeding programmes. Livest. Prod. Sci. 2000, 63, 99–109. [Google Scholar] [CrossRef]
- Taberlet, P.; Valentini, A.; Rezaei, H.R.; Naderi, S.; Pompanon, F.; Negrini, R.; Ajmone-Marsan, P. Are cattle, sheep, and goats endangered species? Mol. Ecol. 2008, 17, 275–284. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Draft Guidelines on in vivo Conservation of Animal Genetic Resources; Number 14 in FAO Animal Production and Health Guidelines; FAO: Rome, Italy, 2013. [Google Scholar]
- Mc Parland, S.; Kearney, J.F.; Rath, M.; Berry, D.P. Inbreeding trends and pedigree analysis of Irish dairy and beef cattle populations. J. Anim. Sci. 2007, 85, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Welsh, C.S.; Stewart, T.S.; Schwab, C.; Blackburn, H.D. Pedigree analysis of 5 swine breeds in the United States and the implications for genetic conservation. J. Anim. Sci. 2010, 88, 1610–1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikhlou, M.; Abbasi, M.A. Genetic diversity of Iranian Lori-Bakhtiari sheep assessed by pedigree analysis. Small Rumin. Res. 2016, 141, 99–105. [Google Scholar] [CrossRef]
- Pham, M.H.; Tran, X.H.; Berthouly-Salazar, C.; Tixier-Boichard, M.; Chen, C.F.; Lee, Y.P. Monitoring of genetic diversity in Taiwan conserved chickens assessed by pedigree and molecular data. Livest. Sci. 2016, 184, 85–91. [Google Scholar] [CrossRef]
- Duru, S. Pedigree analysis of the Turkish Arab horse population: Structure, inbreeding and genetic variability. Animal 2017, 11, 1449–1456. [Google Scholar] [CrossRef]
- Sakthivel, M.; Balasubramanyam, D.; Kumarasamy, P.; Raja, A.; Anilkumar, R.; Gopi, H.; Devaki, A. Genetic structure of a small closed population of the New Zealand white rabbit through pedigree analyses. World Rabbit Sci. 2018, 26, 101–112. [Google Scholar] [CrossRef]
- Wijnrocx, K.; François, L.; Stinckens, A.; Janssens, S.; Buys, N. Half of 23 Belgian dog breeds has a compromised genetic diversity, as revealed by genealogical and molecular data analysis. J. Anim. Breed. Genet. 2016, 135, 375–383. [Google Scholar] [CrossRef]
- Navas, F.J.; Jordana, J.; León, J.M.; Delgado, C.B.J.V. A model to infer the demographic structure evolution of endangered donkey populations. Animal 2017, 11, 2129–2138. [Google Scholar] [CrossRef]
- de Roos, A.P.W.; Schrooten, C.; Veerkamp, R.F.; van Arendonk, J.A.M. Effects of genomic selection on genetic improvement, inbreeding, and merit of young versus proven bulls. J. Dairy Sci. 2011, 94, 1559–1567. [Google Scholar] [CrossRef] [Green Version]
- Elsen, J.M. Prediction of genetic gain in finite populations with heterogeneous predicted breeding values accuracies. J. Anim. Breed. Genet. 2016, 133, 493–502. [Google Scholar] [CrossRef]
- Brother Adam. Bienenzüchtung. Deutsch. Bienenw. 1953, 4, 161–166. [Google Scholar]
- Ruttner, H. Technical recommendations for methods of evaluating performance of bee colonies. In Controlled Mating and Selection of the Honey Bee; Ruttner, F., Ed.; Apimondia Publishing House: Bucharest, Romania, 1972; pp. 87–92. [Google Scholar]
- Bienefeld, K.; Ehrhardt, K.; Reinhardt, F. Genetic evaluation in the honey bee considering queen and worker effects—A BLUP-Animal Model approach. Apidologie 2007, 38, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Bienefeld, K.; Ehrhardt, K.; Reinhardt, F. Noticeable success in honey bee selection after the introduction of genetic evaluation by BLUP. Am. Bee J. 2008, 148, 739–742. [Google Scholar]
- Deutscher Imkerbund. Richtlinien für das Zuchtwesen des Deutschen Imkerbundes (ZRL); Deutscher Imkerbund: Wachtberg, Germany, 2017. [Google Scholar]
- AG Toleranzzucht. Methodenhandbuch der Arbeitsgemeinschaft Toleranzzucht, 2nd ed.; AG Toleranzzucht: Kirchhain, Germany, 2013. [Google Scholar]
- Guichard, M.; Neuditschko, M.; Soland, G.; Fried, P.; Grandjean, M.; Gerster, S.; Dainat, B.; Bijma, P.; Brascamp, E.W. Estimates of genetic parameters for production, behaviour, and health traits in two Swiss honey bee populations. Apidologie 2020. [Google Scholar] [CrossRef]
- Tarpy, D.R.; Nielsen, D.I. Sampling error, effective paternity, and estimating the genetic structure of honey bee colonies (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 2002, 95, 513–528. [Google Scholar] [CrossRef] [Green Version]
- Brascamp, E.W.; Willam, A.; Boigenzahn, C.; Bijma, P.; Veerkamp, R.F. Heritabilities and genetic correlations for honey yield, gentleness, calmness and swarming behaviour in Austrian honey bees. Apidologie 2016, 47, 739–748. [Google Scholar] [CrossRef] [Green Version]
- Brascamp, E.W.; Bijma, P. A note on genetic parameters and accuracy of estimated breeding values in honey bees. Genet. Sel. Evol. 2019, 51, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plate, M.; Bernstein, R.; Hoppe, A.; Bienefeld, K. The importance of controlled mating in honeybee breeding. Genet. Sel. Evol. 2019, 51, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackensen, O. Breeding and genetics of bees. In Beekeeping in the United States; US Agricultural Research Service, Ed.; US Government Printing Office: Washington, DC, USA, 1967; Volume 335, pp. 68–76. [Google Scholar]
- Jensen, A.B.; Palmer, K.A.; Chaline, N.; Raine, N.E.; Tofilski, A.; Martin, S.J.; Pedersen, B.V.; Boomsma, J.J.; Ratnieks, F.L.W. Quantifying honey bee mating range and isolation in semi-isolated valleys by DNA microsatellite paternity analysis. Conserv. Genet. 2005, 6, 527–537. [Google Scholar] [CrossRef]
- Moritz, R.F.A. Comparison of within-family and mass selection in honeybee populations. J. Apicult. Res. 1986, 25, 146–153. [Google Scholar] [CrossRef]
- Page, R.E.; Laidlaw, H.H. Closed population honeybee breeding. Bee World 1985, 66, 63–72. [Google Scholar] [CrossRef]
- Pedersen, L.D.; Sørensen, A.C.; Henryon, M.; Ansari-Mahyari, S.; Berg, P. ADAM: A computer program to simulate selective breeding schemes for animals. Livest. Sci. 2009, 121, 343–344. [Google Scholar] [CrossRef]
- Sargolzaei, M.; Schenkel, F.S. QMSim: A large-scale genome simulator for livestock. Bioinformatics 2009, 25, 680–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S. Evolution in Mendelian populations. Genetics 1931, 16, 97–159. [Google Scholar]
- Bienefeld, K.; Pirchner, F. Heritabilities for several colony traits in the honeybee (Apis mellifera carnica). Apidologie 1990, 21, 175–183. [Google Scholar] [CrossRef]
- Misztal, I.; Tsuruta, S.; Strabel, T.; Auvray, B.; Druet, T.; Lee, D.H. BLUPF90 and related programs (BGF90). In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 19–23 August 2002. [Google Scholar]
- LIB Hohen Neuendorf. beebreed.eu. 2020. Available online: www2.hu-berlin.de/beebreed/ZWS/ (accessed on 29 June 2020).
- Bulmer, M. The effect of selection on genetic variability. Am. Nat. 1971, 105, 201–211. [Google Scholar] [CrossRef]
- Moritz, R.F.A.; Jordan, M. Selection of resistance against Varroa jacobsoni across caste and sex in the honeybee (Apis mellifera L., Hymenoptera: Apidae). Exp. Appl. Acarol. 1992, 16, 345–353. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Hayfield, T.; Racine, J.S. Nonparametric econometrics: The np package. J. Stat. Softw. 2008, 27, 5. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, I.; Goyache, F.; Molina, A.; Valera, M.; Gutiérrez, J.P. Estimation of effective population size from the rate of coancestry in pedigreed populations. J. Anim. Breed. Genet. 2011, 128, 56–63. [Google Scholar] [CrossRef]
- Döke, M.A.; Frazier, M.; Grozinger, C.M. Overwintering honey bees: Biology and management. Curr. Opin. Insect Sci. 2015, 10, 185–193. [Google Scholar] [CrossRef]
- Kristensen, T.N.; Sørensen, P. Inbreeding—Lessons from animal breeding, evolutionary biology and conservation genetics. Anim. Sci. 2005, 80, 121–133. [Google Scholar] [CrossRef]
- Dempfle, L. A note on increasing the limit of selection through selection within families. Genet. Res. 1974, 24, 127–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goddard, M.E. Optimal effective population size for the global population of black and white dairy cattle. J. Dairy Sci. 1992, 75, 2902–2911. [Google Scholar] [CrossRef]
- Hill, W.G. Is continued genetic improvement of livestock sustainable? Genetics 2016, 202, 877–881. [Google Scholar] [CrossRef] [Green Version]
- Hallander, J.; Waldmann, P. The effect of non-additive genetic interactions on selection in multi-locus genetic models. Heredity 2007, 98, 349–359. [Google Scholar] [CrossRef]
- Mulder, H.A.; Lee, S.H.; Clark, S.; Hayes, B.J.; van der Werf, J.H.J. The impact of genomic and traditional selection on the contribution of mutational variance to long-term selection response and genetic variance. Genetics 2019, 213, 361–378. [Google Scholar] [CrossRef]
- Franklin, I.R. Evolutionary change in small populations. In Conservation Biology: An Evolutionary-Ecological Perspective; Soulé, M.E., Wilcox, B.A., Eds.; Sinauer: Sunderland, MA, USA, 1980; pp. 135–150. [Google Scholar]
- Soulé, M.E. Thresholds for survival: Maintaining fitness and evolutionary potential. In Conservation biology: An evolutionary-ecological perspective; Soulé, M.E., Wilcox, B.A., Eds.; Sinauer: Sunderland, MA, USA, 1980; pp. 151–170. [Google Scholar]
- Shaffer, M.L. Minimum population sizes for spezies conservation. BioScience 1981, 31, 131–134. [Google Scholar] [CrossRef]
- Willi, Y.; Buskirk, J.V.; Hoffmann, A.A. Limits to the adaptive potential of small populations. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 433–458. [Google Scholar] [CrossRef]
- Frankham, R.; Bradshaw, C.J.A.; Brook, B.W. Genetics in conservation management: Revised recommendations for the 50/500 rules, Red List criteria and population viability analyses. Biol. Conserv. 2014, 170, 56–63. [Google Scholar] [CrossRef]
- Bijma, P. Long-Term Genetic Contributions: Prediction of Rates of Inbreeding and Genetic Gain in Selected Populations. Ph.D. Thesis, Wageningen Universiteit, Wageningen, The Netherlands, 2000. [Google Scholar]
- Brückner, D. Effects of inbreeding on worker honeybees. Bee World 1979, 60, 137–140. [Google Scholar] [CrossRef]
- Bienefeld, K.; Reinhardt, F.; Pirchner, F. Inbreeding effects of queen and workers on colony traits in the honey bee. Apidologie 1989, 20, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Beye, M. The dice of fate: The csd gene and how its allelic composition regulates sexual development in the honey bee, Apis mellifera. BioEssays 2004, 26, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Zayed, A. Bee genetics and conservation. Apidologie 2009, 40, 237–262. [Google Scholar] [CrossRef] [Green Version]
- Kondrashov, A.S.; Crow, J.F. Haploidy or diploidy: Which is better? Nature 1991, 351, 314–315. [Google Scholar] [CrossRef] [PubMed]
- Dempster, E.R. Genetic models in relation to animal breeding. Biometrics 1955, 11, 535–536. [Google Scholar] [CrossRef]
- Robertson, A. A theory of limits in artificial selection. Proc. R. Soc. B 1960, 153, 234–249. [Google Scholar] [CrossRef]
- Wegener, J.; May, T.; Kamp, G.; Bienefeld, K. A successful new approach to honeybee semen cryopreservation. Cryobiology 2014, 69, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Eynard, S.E.; Windig, J.J.; Hulsegge, I.; Hiemstra, S.J.; Calus, M.P.L. The impact of using old germplasm on genetic merit and diversity—A cattle breed case study. J. Anim. Breed. Genet. 2018, 135, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Roehe, R.; Kennedy, B.W. Effect of selection for maternal and direct genetic effects on genetic improvement of litter size in swine. J. Anim. Sci. 1993, 71, 2891–2904. [Google Scholar] [CrossRef] [Green Version]
- Khaw, H.L.; Ponzoni, P.W.; Bijma, P. Indirect genetic effects and inbreeding: Consequences of BLUP selection for socially affected traits on rate of inbreeding. Genet. Sel. Evol. 2014, 46, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belonsky, G.M.; Kennedy, B.W. Selection on individual phenotype and best linear unbiased predictor of breeding value in a closed swine herd. J. Anim. Sci. 1988, 66, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Bouga, M.; Alaux, C.; Bienkowska, M.; Büchler, R.; Carreck, N.L.; Cauia, E.; Chlebo, R.; Dahle, B.; Dall’Olio, R.; De La Rúa, P.; et al. A review of methods for discrimination of honey bee populations as applied to European beekeeping. J. Apicult. Res. 2011, 50, 51–84. [Google Scholar] [CrossRef] [Green Version]
- Foulley, J.L.; Shaeffer, L.R.; Song, H.; Wilton, J.W. Progeny group size in an organized progeny test program of Al beef bulls using reference sires. Can. J. Anim. Sci. 1983, 63, 17–26. [Google Scholar] [CrossRef]
- Gandini, G.; Corvo, M.D.; Biscarini, F.; Stella, A. Genetic improvement of small ruminant local breeds with nucleus and inbreeding control: A simulation study. Small Rumin. Res. 2014, 120, 196–203. [Google Scholar] [CrossRef]
- Gandini, G.; Avon, L.; Bohte-Wilhelmus, D.; Bay, E.; Colinet, F.G.; Choroszy, Z.; Díaz, C.; Duclos, D.; Fernández, J.; Gengler, N.; et al. Motives and values in farming local cattle breeds in Europe: A survey on 15 breeds. Anim. Genet. Res. 2010, 47, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, Ă.; Lokar, V.; Smodiš Škerl, M.I. Testing of the isolation of the Rog-Ponikve mating station for Carniolan (Apis mellifera carnica) honey bee queens. J. Apic. Res. 2008, 47, 137–140. [Google Scholar] [CrossRef]
- Cobey, S.W.; Tarpy, D.R.; Woyke, J. Standard methods for instrumental insemination of Apis mellifera queens. J. Apicult. Res. 2013, 52, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Pieplow, J.T.; Brauße, J.; van Praagh, J.P.; Moritz, R.F.A.; Erler, S. A scientific note on using large mixed sperm samples in instrumental insemination of honeybee queens. Apidologie 2017, 48, 716–718. [Google Scholar] [CrossRef] [Green Version]
- Gerula, D.; Węgrzynowicz, P.; Panasiuk, B.; Bieńkowska, M.; Skowronek, W. Performance of bee colonies headed by queens instrumentally inseminated with semen of drones who come from a single colony or many colonies. J. Apic. Sci. 2014, 58, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Harbo, J.R. The value of single-drone inseminations in selective breeding of honey bees. In Apiculture for the 21st Century; Connor, L., Hoopingarner, R., Eds.; Wicwas Press: Cheshire, UK, 1999; pp. 1–5. [Google Scholar]
- Chapman, N.C.; Lim, J.; Oldroyd, B.P. Population Genetics of Commercial and Feral Honey Bees in Western Australia. J. Econ. Entomol. 2008, 101, 272–277. [Google Scholar] [CrossRef]
- Zareba, J.; Blazej, P.; Laszkiewicz, A.; Sniezewski, L.; Majkowski, M.; Janik, S.; Cebrat, M. Uneven distribution of complementary sex determiner (csd) alleles in Apis mellifera population. Sci. Rep. 2017, 7, 2317. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E. Maximizing the response of selection with a predefined rate of inbreeding. J. Anim. Sci. 1997, 75, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Reinsch, N.; Spötter, A.; Conrad, T.; Bienefeld, K. Accuracy of the unified approach in maternally influenced traits - illustrated by a simulation study in the honey bee (Apis mellifera). BMC Genet. 2013, 14, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brascamp, E.W.; Wanders, T.H.V.; Wientjes, Y.C.J.; Bijma, P. Prospects for genomic selection in honey-bee breeding. In Proceedings of the 11th World Congress on Genetics Applied to Livestock Production, Auckland, New Zealand, 11–16 February 2018. [Google Scholar]
- Jones, J.C.; Du, Z.G.; Bernstein, R.; Meyer, M.; Hoppe, A.; Schilling, E.; Ableitner, M.; Juling, K.; Dick, R.; Strauss, A.S.; et al. Tool for genomic selection and breeding to evolutionary adaptation: Development of a 100K single nucleotide polymorphism array for the honey bee. Ecol. Evol. 2020. [Google Scholar] [CrossRef]
- Doekes, H.P.; Veerkamp, R.F.; Bijma, P.; Hiemstra, S.J.; Windig, J.J. Trends in genome-wide and region-specific genetic diversity in the Dutch-Flemish Holstein–Friesian breeding program from 1986 to 2015. Genet. Sel. Evol. 2018, 50, 15. [Google Scholar] [CrossRef] [Green Version]
- Makanjuola, B.O.; Miglior, F.; Abdalla, E.A.; Maltecca, C.; Schenkel, F.S.; Baes, C.F. Effect of genomic selection on rate of inbreeding and coancestry and effective population size of Holstein and Jersey cattle populations. J. Dairy Sci. 2020, 103, 5183–5199. [Google Scholar] [CrossRef]
- de Cara, M.A.R.; Fernández, J.; Toro, M.A.; Villanueva, B. Using genome-wide information to minimize the loss of diversity in conservation programmes. J. Anim. Breed. Genet. 2011, 128, 456–464. [Google Scholar] [CrossRef]
- Bosse, M.; Megens, H.J.; Madsen, O.; Crooijmans, R.P.M.A.; Ryder, O.A.; Austerlitz, F.; Groenen, M.A.M.; de Cara, M.A.R. Using genome-wide measures of coancestry to maintain diversity and fitness in endangered and domestic pig populations. Genome Res. 2015, 25, 970–981. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Henryon, M.; Sørensen, A.C. Mating strategies with genomic information reduce rates of inbreeding in animal breeding schemes without compromising genetic gain. Animal 2017, 11, 547–555. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: The datasets used and analysed during the current study are available from the corresponding author on reasonable request. The source code of the simulation program BeeSim is available at https://doi.org/10.5061/dryad.1nh544n. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Definition | Values |
|---|---|---|
| N | Number of breeding queens per year. | |
| Sister group size, number of queens produced by one dam. | ||
| Percentage of breeding queens selected to produce sires. | ||
| Selection scheme defined by key values and . | ||
| Number of loci to determine a trait. | ||
| Maternal and direct additive genetic variance. | ||
| Genetic covariance between maternal and direct effects. | a, b | |
| Genetic correlation between maternal and direct effects. | a, b | |
| Residual variance. | 1 | |
| Genetic standard deviation, inheritance criterion. | a, b | |
| Total heritability. | a, b | |
| Maternal and direct heritabilities. | a, b | |
| Trait determined by loci and correlation between effects. | , , , |
| CBS with Lowest | CBS with Highest | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| var c | var c | |||||||||||
| 70 | 14 | 0 | 3 | 10 | 0.64% | 16.89% | 10 | 3 | 2.01% | 6.34% | ||
| 48 | 5 | 0 | 4 | 10 | 0.81% | 21.67% | 10 | 5 | 1.82% | 12.80% | ||
| 16 | 3 | 0 | 2 | 6 | 0.88% | 28.26% | 8 | 8 | 1.73% | 10.24% | ||
| 37 | 1 | 0 | 3 | 9 | 0.92% | 36.76% | 7 | 4 | 1.94% | 15.80% | ||
| 74 | 46 | 5 | 3 | 7 | 0.39% | 15.45% | 9 | 1 | 1.67% | 5.29% | ||
| 64 | 42 | 2 | 4 | 8 | 0.46% | 25.10% | 10 | 2 | 1.55% | 12.59% | ||
| 32 | 17 | 0 | 4 | 10 | 0.60% | 27.34% | 9 | 5 | 1.51% | 8.49% | ||
| 47 | 11 | 0 | 5 | 9 | 0.74% | 37.22% | 9 | 3 | 1.64% | 15.66% | ||
| 73 | 64 | 23 | 4 | 9 | 0.25% | 15.13% | 10 | 1 | 1.39% | 5.15% | ||
| 64 | 56 | 18 | 5 | 8 | 0.32% | 25.06% | 10 | 1 | 1.42% | 11.02% | ||
| 63 | 48 | 10 | 3 | 8 | 0.32% | 34.02% | 10 | 3 | 1.41% | 7.29% | ||
| 49 | 34 | 0 | 6 | 8 | 0.58% | 35.46% | 10 | 2 | 1.51% | 13.35% | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plate, M.; Bernstein, R.; Hoppe, A.; Bienefeld, K. Long-Term Evaluation of Breeding Scheme Alternatives for Endangered Honeybee Subspecies. Insects 2020, 11, 404. https://doi.org/10.3390/insects11070404
Plate M, Bernstein R, Hoppe A, Bienefeld K. Long-Term Evaluation of Breeding Scheme Alternatives for Endangered Honeybee Subspecies. Insects. 2020; 11(7):404. https://doi.org/10.3390/insects11070404
Chicago/Turabian StylePlate, Manuel, Richard Bernstein, Andreas Hoppe, and Kaspar Bienefeld. 2020. "Long-Term Evaluation of Breeding Scheme Alternatives for Endangered Honeybee Subspecies" Insects 11, no. 7: 404. https://doi.org/10.3390/insects11070404
APA StylePlate, M., Bernstein, R., Hoppe, A., & Bienefeld, K. (2020). Long-Term Evaluation of Breeding Scheme Alternatives for Endangered Honeybee Subspecies. Insects, 11(7), 404. https://doi.org/10.3390/insects11070404