Prophylactic Avoidance of Hazardous Prey by the Ant Host Myrmica rubra


Abstract
:1. Introduction
2. Material and Methods
2.1. Studied Ant Species and Rearing of Ant Colonies
2.2. Rearing of Fruit Flies
2.3. Metarhizium Brunneum Entomopathogenic Fungus
2.4. Tested Conditions
2.5. Experimental Design
2.6. Data Collected on Video Recordings
2.7. Statistical Analyses
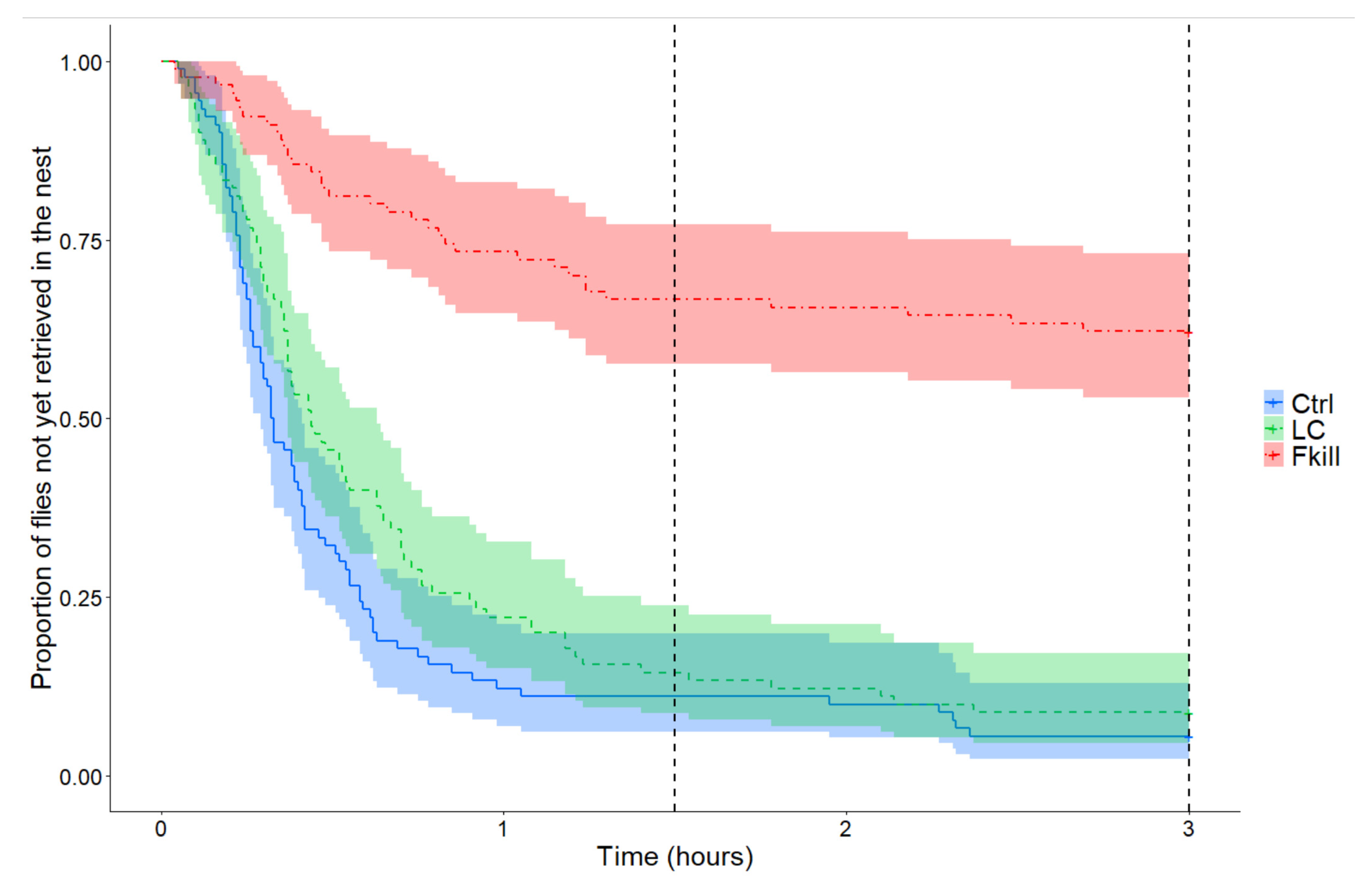
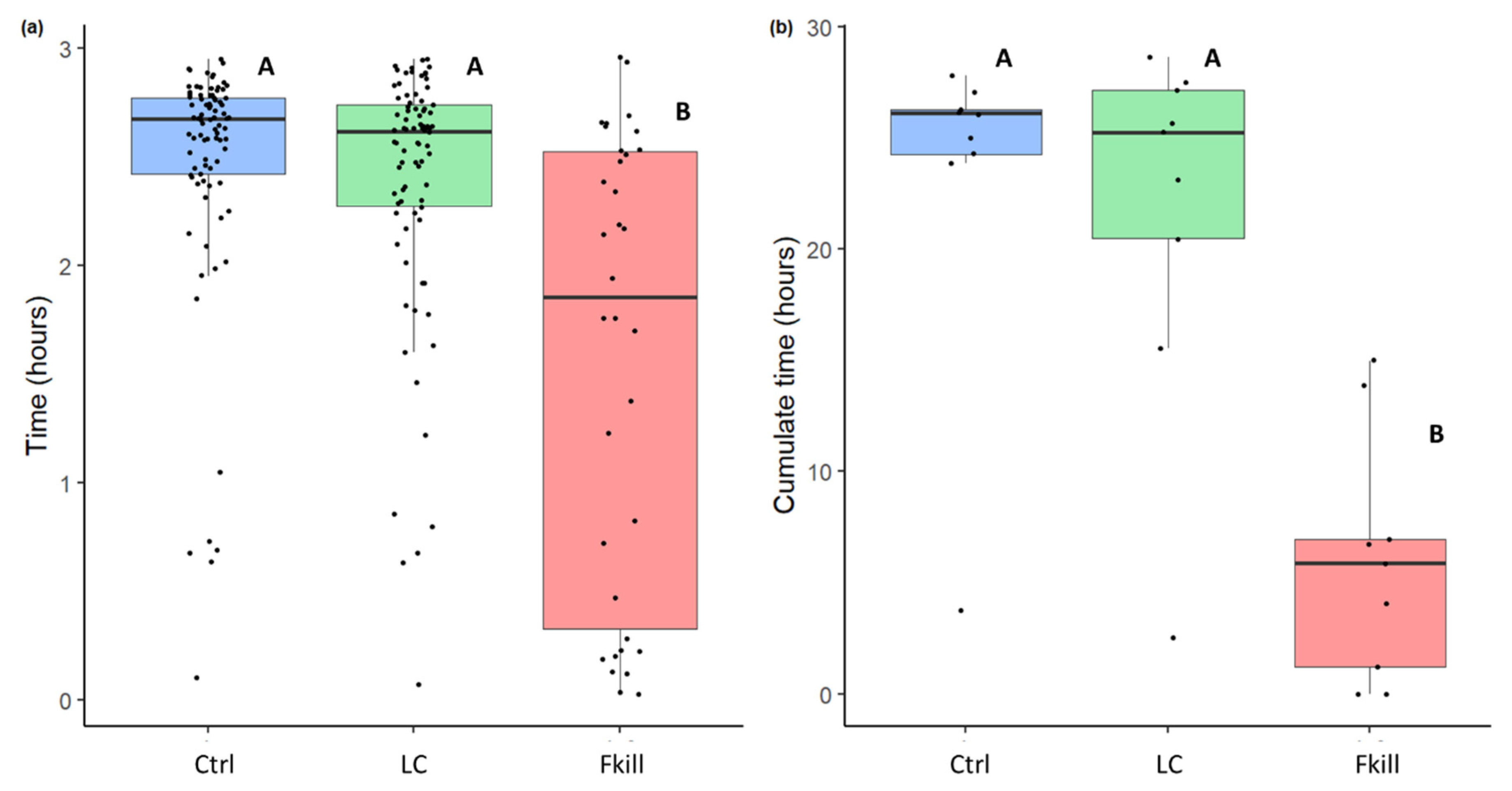
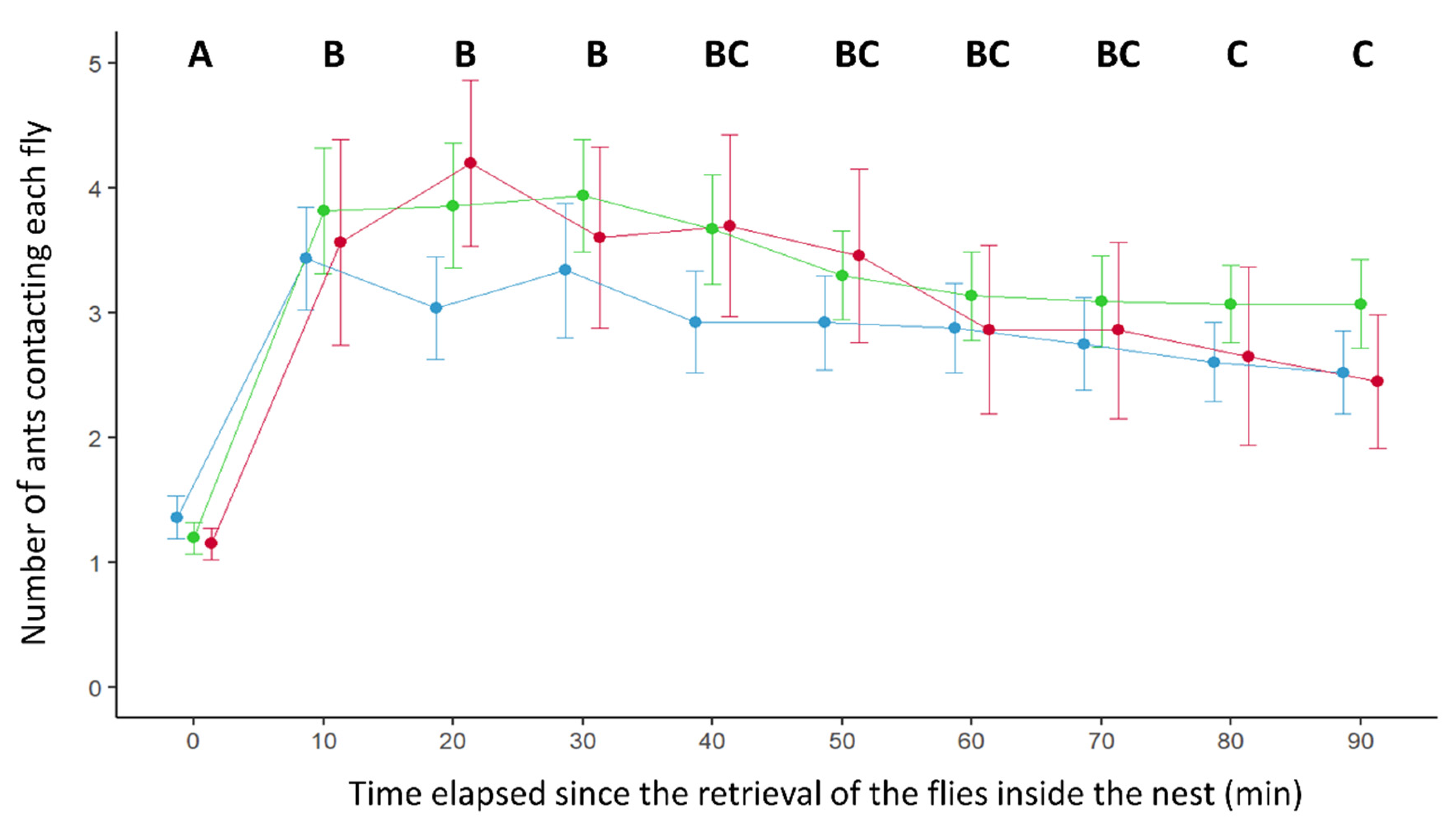
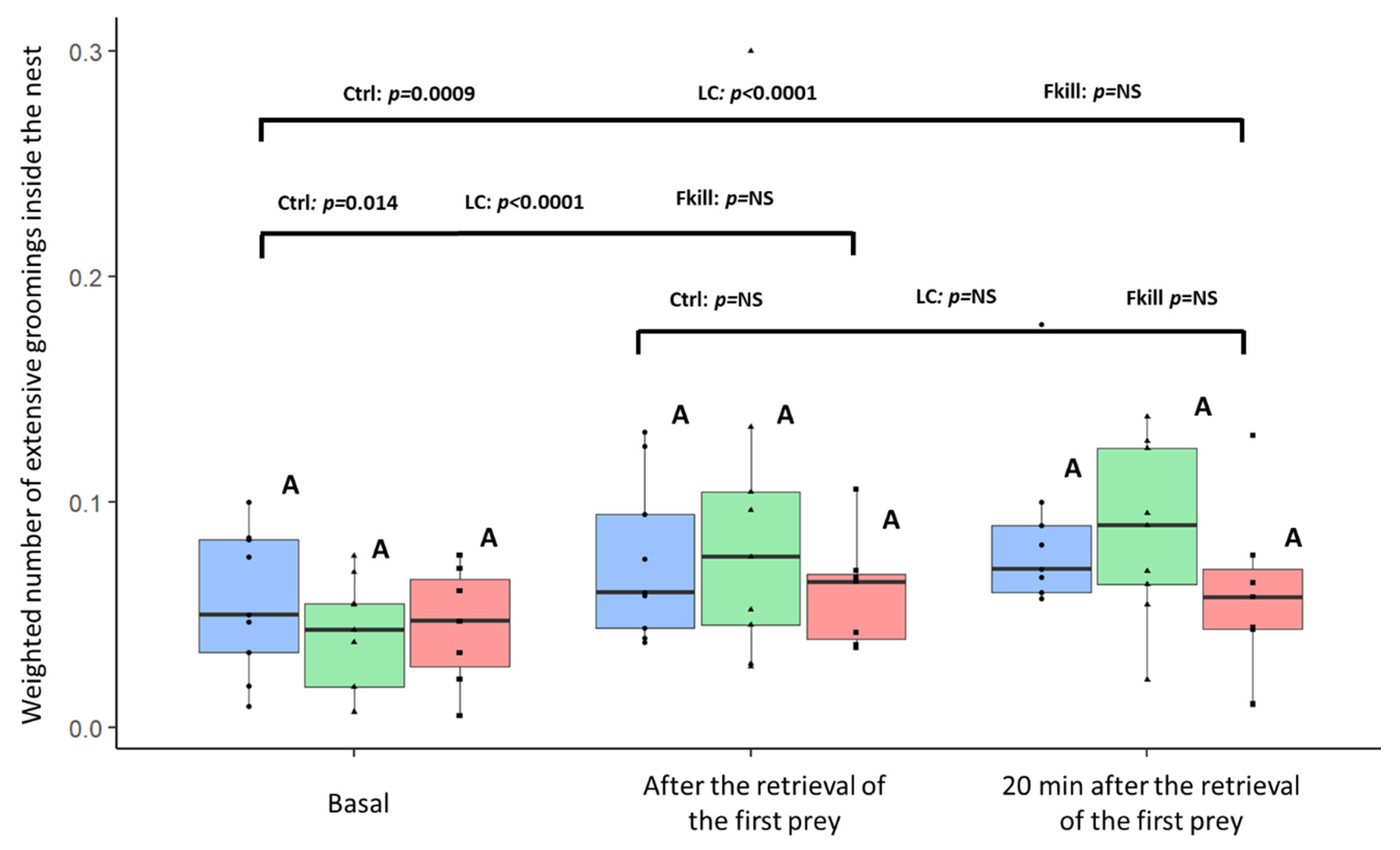
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Availability
References
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990; ISBN 978-0-674-04075-5. [Google Scholar]
- Duarte, A.; Weissing, F.J.; Pen, I.; Keller, L. An evolutionary perspective on self-organized division of labor in social insects. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 91–110. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Hempel, P. Parasites in Social Insects; Princeton University Press: Princeton, NJ, USA, 1998; ISBN 978-0-691-05924-2. [Google Scholar]
- Boomsma, J.J.; Schmid-Hempel, P.; Hughes, W.O.H. Life histories and parasite pressure across the major groups of social insects. In Insect Evolutionary Ecology; CABI: Wallingford, UK, 2005; Volume 211. [Google Scholar]
- Siva-Jothy, M.T.; Moret, Y.; Rolff, J. Insect immunity: An evolutionary ecology perspective. In Advances in Insect Physiology; Elsevier: Amsterdam, The Netherlands, 2005; Volume 32, pp. 1–48. ISBN 978-0-12-024232-0. [Google Scholar]
- Tsakas, S.; Marmaras, V.J. Insect immunity and its signalling: An overview. Invertebr. Surviv. J. 2010, 7, 228–238. [Google Scholar]
- Cremer, S.; Armitage, S.A.O.; Schmid-Hempel, P. Social immunity. Curr. Biol. 2007, 17, 693–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, S. Social immunity in insects. Curr. Biol. 2019, 29, R458–R463. [Google Scholar] [CrossRef] [PubMed]
- Meunier, J. Social immunity and the evolution of group living in insects. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140102. [Google Scholar] [CrossRef] [Green Version]
- Cremer, S.; Pull, C.D.; Fürst, M.A. Social immunity: Emergence and evolution of colony-level disease protection. Annu. Rev. Entomol. 2017, 63, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Konrad, M.; Pull, C.D.; Metzler, S.; Seif, K.; Naderlinger, E.; Grasse, A.V.; Cremer, S. Ants avoid superinfections by performing risk-adjusted sanitary care. Proc. Natl. Acad. Sci. USA 2018, 201713501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calleri, D.V.; Rosengaus, R.B.; Traniello, J.F.A. Disease resistance in the drywood termite, Incisitermes schwarzi: Does nesting ecology affect immunocompetence? J. Insect Sci. 2010, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Traniello, J.F.; Rosengaus, R.B.; Savoie, K. The development of immunity in a social insect: Evidence for the group facilitation of disease resistance. Proc. Natl. Acad. Sci. USA 2002, 99, 6838–6842. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D.; Spivak, M. Socialized medicine: Individual and communal disease barriers in honey bees. J. Invertebr. Pathol. 2010, 103, S62–S72. [Google Scholar] [CrossRef]
- Hoggard, S.J.; Wilson, P.D.; Beattie, A.J.; Stow, A.J. Social complexity and nesting habits are factors in the evolution of antimicrobial defences in wasps. PLoS ONE 2011, 6, e21763. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, X.-Y.; Tang, Q.-B.; Lei, C.-L.; Huang, Q.-Y. The mechanisms of social immunity against fungal infections in eusocial insects. Toxins 2019, 11, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson-Rich, N.; Spivak, M.; Fefferman, N.H.; Starks, P.T. Genetic, individual, and group facilitation of disease resistance in insect societies. Annu. Rev. Entomol. 2009, 54, 405–423. [Google Scholar] [CrossRef] [Green Version]
- Stroeymeyt, N.; Grasse, A.V.; Crespi, A.; Mersch, D.P.; Cremer, S.; Keller, L. Social network plasticity decreases disease transmission in a eusocial insect. Science 2018, 362, 941–945. [Google Scholar] [CrossRef] [Green Version]
- Naug, D.; Smith, B. Experimentally induced change in infectious period affects transmission dynamics in a social group. Proc. R. Soc. B Biol. Sci. 2007, 274, 61–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stroeymeyt, N.; Casillas-Pérez, B.; Cremer, S. Organisational immunity in social insects. Curr. Opin. Insect Sci. 2014, 5, 1–15. [Google Scholar] [CrossRef]
- Verza, S.S.; Diniz, E.A.; Chiarelli, M.F.; Mussury, R.M.; Bueno, O.C. Waste of leaf-cutting ants: Disposal, nest structure, and abiotic soil factors around internal waste chambers. Acta Ethol. 2017, 20, 119–126. [Google Scholar] [CrossRef]
- Weiss, M.R. Defecation Behavior and Ecology of Insects. Annu. Rev. Entomol. 2006, 51, 635–661. [Google Scholar] [CrossRef]
- Diez, L.; Deneubourg, J.-L.; Detrain, C. Social prophylaxis through distant corpse removal in ants. Naturwissenschaften 2012, 99, 833–842. [Google Scholar] [CrossRef]
- Diez, L.; Lejeune, P.; Detrain, C. Keep the nest clean: Survival advantages of corpse removal in ants. Biol. Lett. 2014, 10, 20140306. [Google Scholar] [CrossRef] [Green Version]
- Diez, L.; Urbain, L.; Lejeune, P.; Detrain, C. Emergency measures: Adaptive response to pathogen intrusion in the ant nest. Behav. Process. 2015, 116, 80–86. [Google Scholar] [CrossRef]
- Visscher, P.K. The honey bee way of death: Necrophoric behaviour in Apis mellifera colonies. Anim. Behav. 1983, 31, 1070–1076. [Google Scholar] [CrossRef]
- Wen, P. Death recognition by undertaker bees. bioRxiv 2020. [Google Scholar] [CrossRef]
- Munday, Z.; Brown, M.J.F. Bring out your dead: Quantifying corpse removal in Bombus terrestris, an annual eusocial insect. Anim. Behav. 2018, 138, 51–57. [Google Scholar] [CrossRef]
- López-Riquelme, G.O.; Fanjul-Moles, M.L. The funeral ways of social insects. Social strategies for corpse disposal. Trends Entomol. 2013, 9, 71–129. [Google Scholar]
- Sun, Q.; Zhou, X. Corpse management in social insects. Int. J. Biol. Sci. 2013, 9, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Heinze, J.; Walter, B. Moribund ants leave their nests to die in social isolation. Curr. Biol. 2010, 20, 249–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclerc, J.-B.; Detrain, C. Loss of attraction for social cues leads to fungal-infected Myrmica rubra ants withdrawing from the nest. Anim. Behav. 2017, 129, 133–141. [Google Scholar] [CrossRef]
- Rueppell, O.; Hayworth, M.K.; Ross, N.P. Altruistic self-removal of health-compromised honey bee workers from their hive: Self-removal in honey bees. J. Evol. Biol. 2010, 23, 1538–1546. [Google Scholar] [CrossRef] [Green Version]
- Reber, A.; Chapuisat, M. Diversity, prevalence and virulence of fungal entomopathogens in colonies of the ant Formica selysi. Insectes Sociaux 2012, 59, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Marikovsky, P.I. On some features of behavior of the ants Formica rufa L. infected with fungous disease. Insectes Sociaux 1962, 9, 173–179. [Google Scholar] [CrossRef]
- Kramm, K.R.; West, D.F.; Rockenbach, P.G. Termite pathogens: Transfer of the entomopathogen Metarhizium anisopliae between Reticulitermes sp. termites. J. Invertebr. Pathol. 1982, 40, 1–6. [Google Scholar] [CrossRef]
- Tranter, C.; LeFevre, L.; Evison, S.E.F.; Hughes, W.O.H. Threat detection: Contextual recognition and response to parasites by ants. Behav. Ecol. 2015, 26, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.; Detrain, C. Pathogen avoidance and prey discrimination in ants. R. Soc. Open Sci. 2020, 7, 191705. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Tian, M.-Y.; He, Y.-R.; Bland, J.M.; Gu, W.-X. Behavioral and electrophysiological responses of Coptotermes formosanus Shiraki towards entomopathogenic fungal volatiles. Biol. Control 2010, 55, 166–173. [Google Scholar] [CrossRef]
- Crespo, R.; Juarez, M.P.; Cafferata, L.F.R. Biochemical Interaction between Entomopathogenous Fungi and Their Insect-Host-like Hydrocarbons. Mycologia 2000, 92, 528–536. [Google Scholar] [CrossRef]
- Cheraghi, A.; Habibpour, B.; Mossadegh, M.S. Application of Bait Treated with the Entomopathogenic Fungus Metarhizium anisopliae (Metsch.) Sorokin for the control of Microcerotermes diversus Silv. Psyche J. Entomol. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Pontieri, L.; Vojvodic, S.; Graham, R.; Pedersen, J.S.; Linksvayer, T.A. Ant colonies prefer infected over uninfected nest sites. PLoS ONE 2014, 9, e111961. [Google Scholar] [CrossRef] [Green Version]
- Qiu, H.-L.; Lu, L.-H.; Zhang, C.-Y.; He, Y.-R. Pathogenicity of individual isolates of entomopathogenic fungi affects feeding preference of red imported fire ants Solenopsis invicta. Biocontrol Sci. Technol. 2014, 24, 1286–1296. [Google Scholar] [CrossRef]
- Leclerc, J.-B.; Silva, J.P.; Detrain, C. Impact of soil contamination on the growth and shape of ant nests. R. Soc. Open Sci. 2018, 5, 180267. [Google Scholar] [CrossRef] [Green Version]
- Brian, M.V.; Abbott, A. The control of food flow in a society of the ant Myrmica rubra L. Anim. Behav. 1977, 25, 1047–1055. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Elmes, G.W. Observations on the density of queens in natural colonies of Myrmica rubra L. (Hymenoptera: Formicidae). J. Anim. Ecol. 1973, 42, 761–771. [Google Scholar] [CrossRef]
- Leclerc, J.-B.; Detrain, C. Ants detect but do not discriminate diseased workers within their nest. Sci. Nat. 2016, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, S.; Kessler, P.; Schweizer, C. Distribution of insect pathogenic soil fungi in Switzerland with special reference to Beauveria brongniartii and Metharhizium anisopliae. Biocontrol 2003, 48, 307–319. [Google Scholar] [CrossRef]
- Goettel, M.S.; Eilenberg, J. Entomopathogenic fungi and their role in regulation of insect populations. In Insect Control: Biological and Synthetic Agents; Academic Press: Cambridge, MA, USA, 2010; pp. 387–432. ISBN 978-0-12-381450-0. [Google Scholar]
- Loreto, R.G.; Hughes, D.P. Disease dynamics in ants. In Advances in Genetics; Elsevier: Amsterdam, The Netherlands, 2016; Volume 94, pp. 287–306. ISBN 978-0-12-804694-4. [Google Scholar]
- Davis, H.E.; Meconcelli, S.; Radek, R.; McMahon, D.P. Termites shape their collective behavioural response based on stage of infection. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulmer, M.S.; Franco, B.A.; Fields, E.G. Subterranean termite social alarm and hygienic responses to fungal pathogens. Insects 2019, 10, 240. [Google Scholar] [CrossRef] [Green Version]
- Simone-Finstrom, M.D.; Spivak, M. Increased resin collection after parasite challenge: A case of self-medication in honey bees? PLoS ONE 2012, 7, e34601. [Google Scholar] [CrossRef] [Green Version]
- Hänel, H. The life cycle of the insect pathogenic fungus Metarhizium anisopliae in the termite Nasutitermes exitiosus. Mycopathologia 1982, 80, 137–145. [Google Scholar] [CrossRef]
- Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A. (Ed.) Academic Press imprint of Elsevier Science: Oxford, UK; New York, NY, USA, 2012; ISBN 978-0-12-386899-2. [Google Scholar]
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000; ISBN 0-387-98784-3. [Google Scholar]
- Therneau, T.M. Coxme: Mixed Effects Cox Models. R package version 2.2-16. 2020. Available online: https://CRAN.R-project.org/package=coxme (accessed on 1 July 2020).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using {lme4}. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference; R package version 1.43.15. 2019. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 1 July 2020).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R package version 1.4.4. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 1 July 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Coblentz, K.E.; Van Bael, S.A. Field colonies of leaf-cutting ants select plant materials containing low abundances of endophytic fungi. Ecosphere 2013, 4, art66. [Google Scholar] [CrossRef]
- Griffiths, H.M.; Hughes, W.O.H. Hitchhiking and the removal of microbial contaminants by the leaf-cutting ant Atta colombica. Ecol. Entomol. 2010, 35. [Google Scholar] [CrossRef]
- Knoch, T.R.; Faeth, S.H.; Arnott, D.L. Endophytic fungi alter foraging and dispersal by desert seed-harvesting ants. Oecologia 1993, 95, 470–473. [Google Scholar] [CrossRef]
- Gayahan, G.G.; Tschinkel, W.R. Fire ants, Solenopsis invicta, dry and store insect pieces for later use. J. Insect Sci. 2008, 8. [Google Scholar] [CrossRef]
- Yanagawa, A.; Yokohari, F.; Shimizu, S. The role of antennae in removing entomopathogenic fungi from cuticle of the termite, Coptotermes formosanus. J. Insect Sci. 2009, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyling, N.V.; Pell, J.K. Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol. Entomol. 2006, 31, 162–171. [Google Scholar] [CrossRef]
- Carroll, C.R.; Janzen, D.H. Ecology of Foraging by Ants. Annu. Rev. Ecol. Syst. 1973, 4, 231–257. [Google Scholar] [CrossRef]
- Vestergaard, S.; Butt, T.M.; Bresciani, J.; Gillespie, A.T.; Eilenberg, J. Light and Electron Microscopy Studies of the Infection of the Western Flower Thrips Frankliniella occidentalis (Thysanoptera: Thripidae) by the Entomopathogenic Fungus Metarhizium anisopliae. J. Invertebr. Pathol. 1999, 73, 25–33. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi. In Advances in Genetics; Elsevier: Amsterdam, The Netherlands, 2016; Volume 94, pp. 307–364. ISBN 978-0-12-804694-4. [Google Scholar]
- Beenakkers, A.M.T.; Gilbert, L.I. The fatty acid composition of fat body and haemolymph lipids in Hyalophora cecropia and its relation to lipid release. J. Insect Physiol. 1968, 14, 481–494. [Google Scholar] [CrossRef]
- Brew, C.R.; O’Dowd, D.J.; Rae, I.D. Seed dispersal by ants: Behaviour-releasing compounds in elaiosomes. Oecologia 1989, 80, 490–497. [Google Scholar] [CrossRef]
- Hughes, L.; Westoby, M.; Jurado, E. Convergence of elaiosomes and insect prey: Evidence from ant foraging behaviour and fatty acid composition. Funct. Ecol. 1994, 8, 358–365. [Google Scholar] [CrossRef]
- Gordon, D.M. Dependence of necrophoric response to oleic acid on social context in the ant, Pogonomyrmex badius. J. Chem. Ecol. 1983, 9, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Diez, L.; Moquet, L.; Detrain, C. Post-mortem changes in chemical profile and their influence on corpse removal in ants. J. Chem. Ecol. 2013, 39, 1424–1432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef]
- Lang, C.; Menzel, F. Lasius niger ants discriminate aphids based on their cuticular hydrocarbons. Anim. Behav. 2011, 82, 1245–1254. [Google Scholar] [CrossRef]
- Binz, H.; Foitzik, S.; Staab, F.; Menzel, F. The chemistry of competition: Exploitation of heterospecific cues depends on the dominance rank in the community. Anim. Behav. 2014, 94, 45–53. [Google Scholar] [CrossRef]
- Baracchi, D.; Fadda, A.; Turillazzi, S. Evidence for antiseptic behaviour towards sick adult bees in honey bee colonies. J. Insect Physiol. 2012, 58, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Rosengaus, R.B.; Jordan, C.; Lefebvre, M.L.; Traniello, J.F.A. Pathogen alarm behavior in a termite: A new form of communication in social insects. Naturwissenschaften 1999, 86, 544–548. [Google Scholar] [CrossRef]
- Cassill, D.L.; Butler, J.; Vinson, S.B.; Wheeler, D.E. Cooperation during prey digestion between workers and larvae in the ant, Pheidole spadonia. Insectes Sociaux 2005, 52, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Sorensen, A.A.; Kamas, R.S.; Vinson, S.B. The influence of oral secretions from larvae on levels of proteinases in colony members of Solenopsis invicta Buren (Hymenoptera: Formicidae). J. Insect Physiol. 1983, 29, 163–168. [Google Scholar] [CrossRef]
- Ishay, J.; Ikan, R. Food exchange between adults and larvae in Vespa orientalis F. Anim. Behav. 1968, 16, 298–303. [Google Scholar] [CrossRef]
- Detrain, C.; Pereira, H.; Fourcassié, V. Differential responses to chemical cues correlate with task performance in ant foragers. Behav. Ecol. Sociobiol. 2019, 73, 107. [Google Scholar] [CrossRef]
- Depickère, S.; Fresneau, D.; Detrain, C.; Deneubourg, J.-L. Marking as a decision factor in the choice of a new resting site in Lasius niger. Insectes Sociaux 2004, 51, 243–246. [Google Scholar] [CrossRef]
- Glancey, B.M.; Vander Meer, R.K.; Glover, A.; Lofgren, C.S.; Vinson, S.B. Filtration of microparticles from liquids ingested by the red imported fire ant Solenopsis invicta Buren. Insectes Sociaux 1981, 28, 395–401. [Google Scholar] [CrossRef]
- Eisner, T.; Happ, G.M. The Infrabuccal Pocket of a Formicine ant: A social filtration device. Psyche (Stuttg.) 1962, 69, 107–116. [Google Scholar] [CrossRef]
- Tragust, S.; Mitteregger, B.; Barone, V.; Konrad, M.; Ugelvig, L.V.; Cremer, S. Ants disinfect fungus-exposed brood by oral uptake and spread of their poison. Curr. Biol. 2013, 23, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tragust, S.; Herrmann, C.; Häfner, J.; Braasch, R.; Tilgen, C.; Hoock, M.; Milidakis, M.A.; Gross, R.; Feldhaar, H. Formicine ants swallow their highly acidic poison for gut microbial selection and control. BioRxiv 2020. [Google Scholar] [CrossRef]
- Fernández-Marín, H.; Zimmermann, J.K.; Wcislo, W.T. Nest-founding in Acromyrmex octospinosus (Hymenoptera, Formicidae, Attini): Demography and putative prophylactic behaviors. Insectes Sociaux 2003, 50, 304–308. [Google Scholar] [CrossRef]
- Binz, H.; Kraft, E.F.; Entling, M.H.; Menzel, F. Behavioral response of a generalist predator to chemotactile cues of two taxonomically distinct prey species. Chemoecology 2016, 26, 153–162. [Google Scholar] [CrossRef]
- Buehlmann, C.; Graham, P.; Hansson, B.S.; Knaden, M. Desert ants locate food by combining high sensitivity to food odors with extensive crosswind runs. Curr. Biol. 2014, 24, 960–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce-Duvet, J.M.C.; Feener, D.H. Resource discovery in ant communities: Do food type and quantity matter? Ecol. Entomol. 2010, 35, 549–556. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Prey | Conidia-Covered Prey | Fungus-Killed Prey | |
|---|---|---|---|
| Number of Flies Retrieved Inside the Nest | 10 [10,10] A (n = 9) | 10 [10,10] A (n = 9) | 4 [1,6] B (n = 9) |
| GLMM: Wald test: W = 29.9, df = 2, p < 0.0001 | |||
| Percentage of Discarded Flies | 7% A (n = 85) | 11% A (n = 82) | 42% B (n = 34) |
| GLMM: Wald test: W = 22.5, df = 2, p < 0.0001 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, H.; Detrain, C. Prophylactic Avoidance of Hazardous Prey by the Ant Host Myrmica rubra. Insects 2020, 11, 444. https://doi.org/10.3390/insects11070444
Pereira H, Detrain C. Prophylactic Avoidance of Hazardous Prey by the Ant Host Myrmica rubra. Insects. 2020; 11(7):444. https://doi.org/10.3390/insects11070444
Chicago/Turabian StylePereira, Hugo, and Claire Detrain. 2020. "Prophylactic Avoidance of Hazardous Prey by the Ant Host Myrmica rubra" Insects 11, no. 7: 444. https://doi.org/10.3390/insects11070444
APA StylePereira, H., & Detrain, C. (2020). Prophylactic Avoidance of Hazardous Prey by the Ant Host Myrmica rubra. Insects, 11(7), 444. https://doi.org/10.3390/insects11070444