Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Identification of Symbionts in Different Whitefly Populations
2.3. Transfer of Whitefly Populations to Natal and Novel Host-Plant Species
2.4. Quantification of Symbionts
2.5. Amino-Acid Analysis
2.6. Data Analysis
3. Results
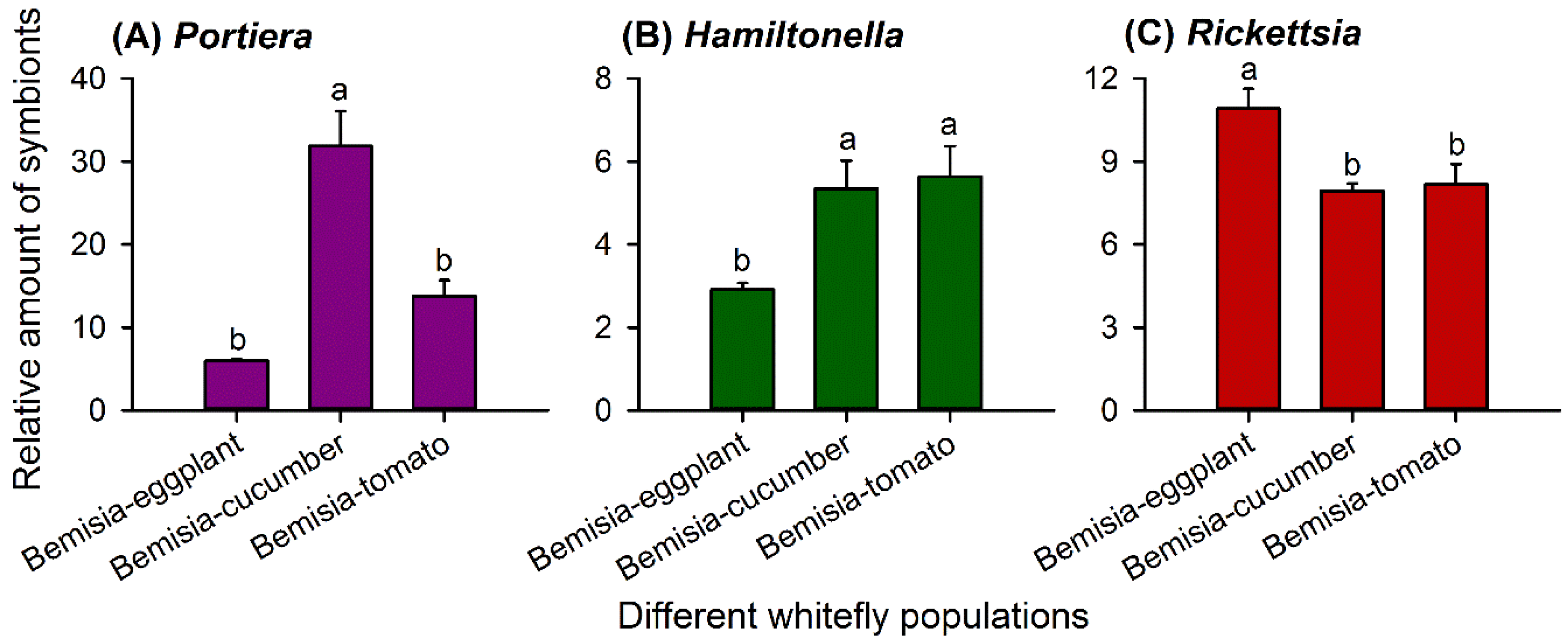
3.1. Different Quantities of the Same Symbiont Taxa in Three Whitefly Populations
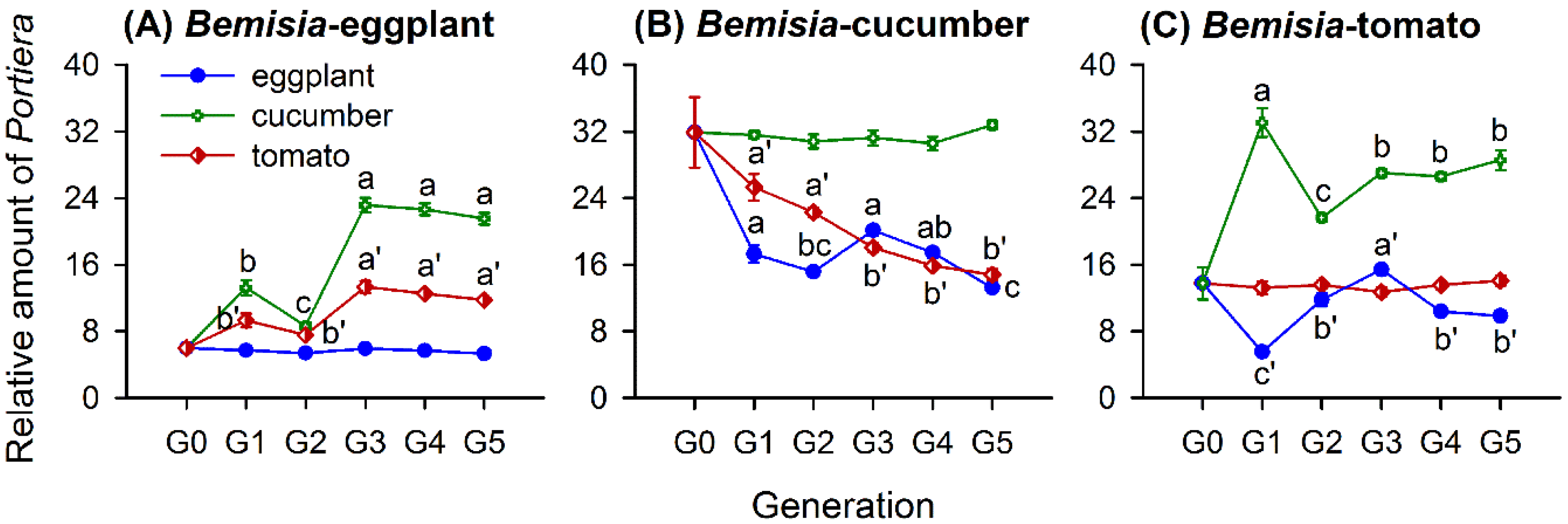
3.2. Host-Plant-Switching Affects Portiera Abundance
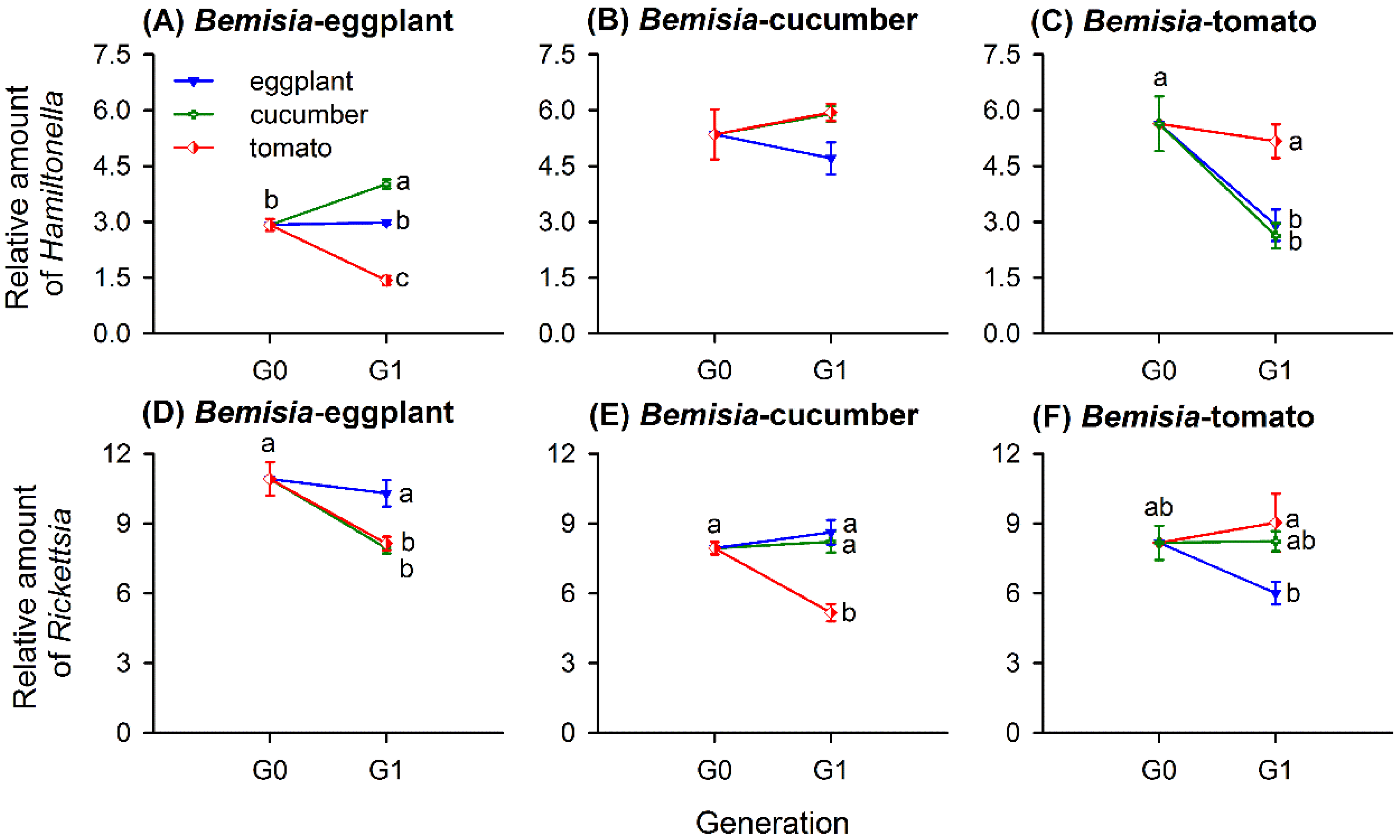
3.3. Host-Plant-Switching Affects S-Symbiont Abundance
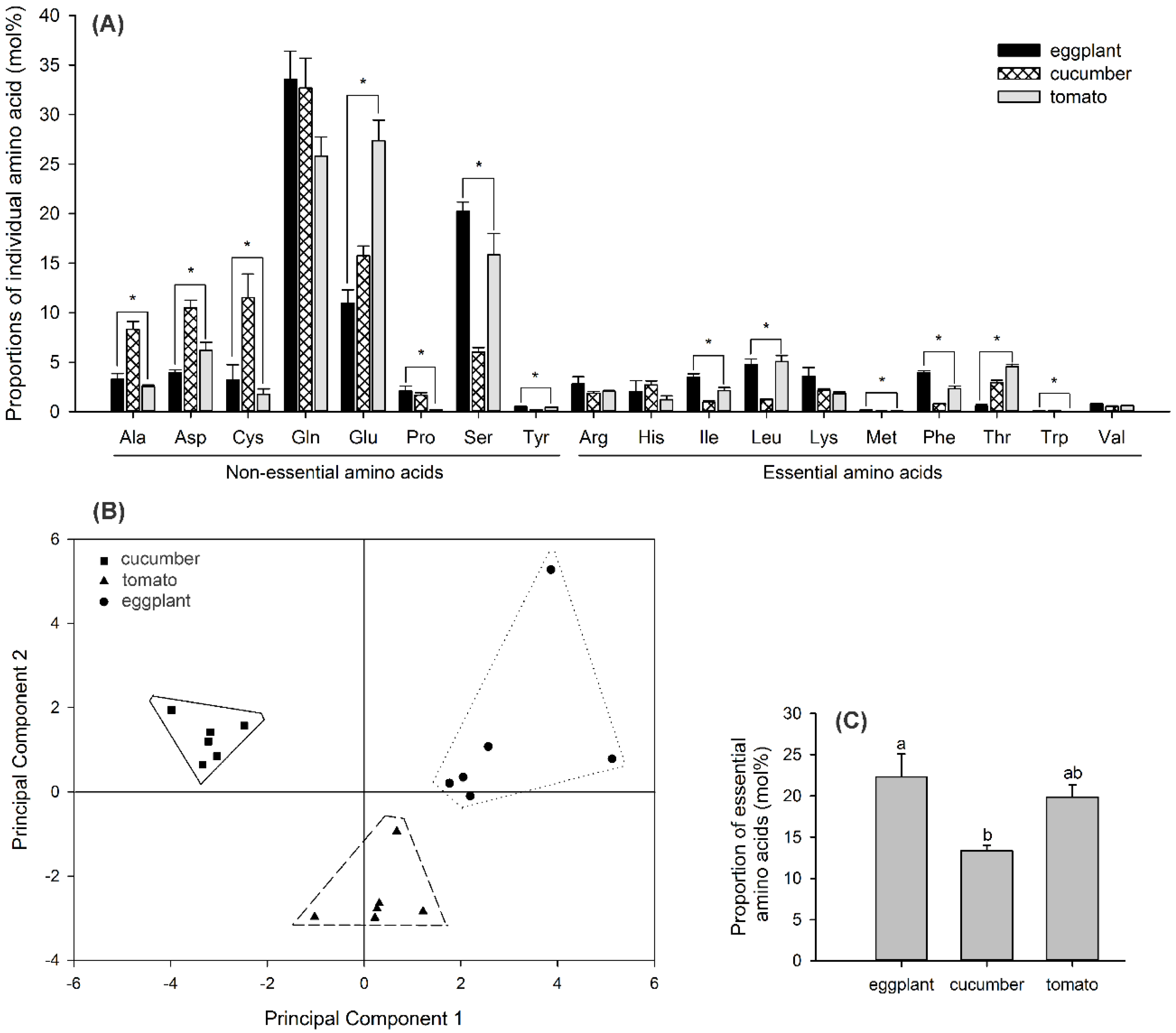
3.4. Phloem Sap Amino Acid–Composition Profiles Differ among Plant Species
3.5. No Significant Differences in Amino Acid–Composition Profiles among Whitefly Populations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cock, M.J.W. Bemisia tabaci–A Literature Survey on the Cotton Whitefly with an Annotated Bibliography; C.A.B International Institute of Biological Control: Ascot, UK, 1986; pp. 1–121. [Google Scholar]
- Li, S.J.; Xue, X.; Ahmed, M.Z.; Ren, S.X.; Du, Y.Z.; Wu, J.H.; Cuthbertson, A.G.S.; Qiu, B.L. Host Plants and Natural Enemies of Bemisia tabaci (Hemiptera: Aleyrodidae) in China. Insect Sci. 2011, 18, 101–120. [Google Scholar] [CrossRef]
- Barro, P.J.D.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A Statement of Species Status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hasegawa, D.K.; Kaur, N.; Kliot, A.; Pinheiro, P.V.; Luan, J.; Stensmyr, M.C.; Zheng, Y.; Liu, W.; Sun, H.; et al. The Draft Genome of Whitefly Bemisia tabaci MEAM1, a Global Crop Pest, Provides Novel Insights into Virus Transmission, Host Adaptation, and Insecticide Resistance. BMC Biol. 2016, 14, 110. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.F.; Xia, F.; Johnson, K.W.; Bartom, E.; Tuteja, J.H.; Stevens, R.; Grossman, R.L.; Brumin, M.; White, K.P.; Ghanim, M. Genome Sequences of the Primary Endosymbiont “Candidatus Portiera aleyrodidarum” in the Whitefly Bemisia tabaci B and Q Biotypes. J. Bacteriol. 2012, 194, 6678–6679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marubayashi, J.M.; Kliot, A.; Yuki, V.A.; Rezende, J.A.M.; Krause-Sakate, R.; Pavan, M.A.; Ghanim, M. Diversity and Localization of Bacterial Endosymbionts from Whitefly Species Collected in Brazil. PLoS ONE 2014, 9, e108363. [Google Scholar] [CrossRef]
- Macfadyen, S.; Paull, C.; Boykin, L.M.; De Barro, P.; Maruthi, M.N.; Otim, M.; Kalyebi, A.; Vassão, D.G.; Sseruwagi, P.; Tay, W.T.; et al. Cassava Whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in East African Farming Landscapes: A Review of the Factors Determining Abundance. Bull. Entomol. Res. 2018, 108, 565–582. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.X. Population Dynamics of Bemisia argentifolii (Homoptera: Aleyrodidae) on Spring Collard and Relationship to Yield in the Lower Rio Grande Valley of Texas. J. Econ. Entomol. 2000, 93, 750–756. [Google Scholar] [CrossRef]
- Wilkinson, T.L.; Adams, D.; Minto, L.B.; Douglas, A.E. The Impact of Host Plant on the Abundance and Function of Symbiotic Bacteria in an Aphid. J. Exp. Biol. 2001, 204, 3027. [Google Scholar]
- Ghosh, S.; Bouvaine, S.; Richardson, S.C.W.; Ghanim, M.; Maruthi, M.N. Fitness Costs Associated with Infections of Secondary Endosymbionts in the Cassava Whitefly Species Bemisia. tabaci. J. Pest Sci. 2018, 91, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Chiel, E.; Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Katzir, N.; Inbar, M.M. Biotype-dependent Secondary Symbiont Communities in Sympatric Populations of Bemisia tabaci. Bull. Entomol. Res. 2007, 97, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Bing, X.L.; Ruan, Y.M.; Rao, Q.; Wang, X.W.; Liu, S.S. Diversity of Secondary Endosymbionts among Different Putative Species of the Whitefly Bemisia tabaci. Insect Sci. 2013, 20, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Zchori-Fein, E.; Lahav, T.; Freilich, S. Variations in the Identity and Complexity of Endosymbiont Combinations in Whitefly Hosts. Front. Microbiol. 2014, 5, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Q.; Xie, W.; Wang, S.L.; Wu, Q.J.; Liu, B.M.; Fang, Y.; Xu, B.Y.; Zhang, Y.J. The Endosymbiont Hamiltonella Increases the Growth Rate of its Host Bemisia tabaci during Periods of Nutritional Stress. PLoS ONE 2014, 9, e89002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bing, X.L.; Yang, J.; Zchori-Fein, E.; Wang, X.W.; Liu, S.S. Characterization of a Newly Discovered Symbiont of the Whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Appl. Environ. Microbiol. 2013, 79, 569–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Garcia, D.; Juravel, K.; Freilich, S.; Zchori-Fein, E.; Latorre, A.; Moya, A.; Morin, S.; Silva, F. To B or not to B: Comparative Genomics Suggests Arsenophonus as a Source of B Vitamins in Whiteflies. Front. Microbiol. 2018, 9, 2254. [Google Scholar] [CrossRef]
- Rao, Q.; Wang, S.; Su, Y.L.; Bing, X.L.; Liu, S.S.; Wang, X.W. Draft Genome Sequence of “Candidatus Hamiltonella defensa,” an Endosymbiont of the Whitefly Bemisia tabaci. J. Bacteriol. 2012, 194, 3558. [Google Scholar] [CrossRef] [Green Version]
- Luan, J.B.; Chen, W.B.; Hasegawa, D.K.; Simmons, A.M.; Wintermantel, W.M.; Ling, K.S.; Fei, Z.J.; Liu, S.S.; Douglas, A.E. Metabolic Coevolution in the Bacterial Symbiosis of Whiteflies and Related Plant Sap-Feeding Insects. Genome Biol. Evol. 2015, 7, 2635–2647. [Google Scholar] [CrossRef] [Green Version]
- Rao, Q.; Rollat-Farnier, P.A.; Zhu, D.T.; Santos-Garcia, D.; Silva, F.J.; Moya, A.; Latorre, A.; Klein, C.C.; Vavre, F.; Sagot, M.F.; et al. Genome Reduction and Potential Metabolic Complementation of the Dual Endosymbionts in the whItefly Bemisia tabaci. BMC Genom. 2015, 16, 226. [Google Scholar] [CrossRef] [Green Version]
- Ankrah, N.Y.D.; Luan, J.B.; Douglas, A.E. Cooperative Metabolism in a Three-Partner Insect-Bacterial Symbiosis Revealed by Metabolic Modeling. J. Bacteriol. 2017, 199, e00872-16. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.B.; Ren, F.R.; Yao, Y.L.; Sun, X.; Walling, L.L.; Li, N.N.; Bai, B.; Bao, X.Y.; Xu, X.R.; Luan, J.B. Intracellular Symbionts Drive Sex Ratio in the Whitefly by Facilitating Fertilization and Provisioning of B Vitamins. ISME J. 2020. [Google Scholar] [CrossRef]
- Ren, F.R.; Sun, X.; Wang, T.Y.; Yao, Y.L.; Huang, Y.Z.; Zhang, X.; Luan, J.B. Biotin Provisioning by Horizontally Transferred Genes from Bacteria Confers Animal Fitness Benefits. ISME J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Lai, C.Y.; Kuo, M.H. Temperature Effect on the Growth of Buchnera Endosymbiont in Aphis craccivora (Hemiptera: Aphididae). Symbiosis 2009, 49, 53. [Google Scholar] [CrossRef]
- Wilkinson, T.L.; Koga, R.; Fukatsu, T. Role of Host Nutrition in Symbiont Regulation: Impact of Dietary Nitrogen on Proliferation of Obligate and Facultative Bacterial Endosymbionts of the Pea aphid Acyrthosiphon pisum. Appl. Environ. Microbiol. 2007, 73, 1362–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.H.; Kang, Z.W.; Guo, Y.; Zhu, G.S.; Shah, M.M.R.; Song, Y.; Fan, Y.L.; Jing, X.F.; Liu, T.X. Nitrogen Hurdle of Host Alternation for a Polyphagous Aphid and the Associated Changes of Endosymbionts. Sci. Rep. 2016, 6, 24781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frohlich, D.R.; Torresjerez, I.I.; Bedford, I.D.; Markham, P.G.; Brown, J.K. A Phylogeographical Analysis of the Bemisia tabaci Species Complex Based on Mitochondrial DNA Markers. Mol. Ecol. 1999, 8, 1683–1691. [Google Scholar] [CrossRef]
- Shah, M.M.R.; Liu, T.X. Feeding Experience of Bemisia tabaci (Hemiptera: Aleyrodidae) Affects their Performance on Different Host Plants. PLoS ONE 2013, 8, e77368. [Google Scholar] [CrossRef] [Green Version]
- Zchori-Fein, E.; Brown, J.K. Diversity of Prokaryotes Associated with Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2002, 95, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, Y.; Ghanim, M.; Chiel, E.; Gerling, D.; Portnoy, V.; Steinberg, S.; Tzuri, G.; Horowitz, A.R.; Belausov, E.; Mozesdaube, N. Identification and Localization of a Rickettsia sp. in Bemisia tabaci (Homoptera: Aleyrodidae). Appl. Environ. Microbiol. 2006, 72, 3646–3652. [Google Scholar] [CrossRef] [Green Version]
- Heddi, A.; Grenier, A.M.; Khatchadourian, C.; Charles, H.; Nardon, P. Four Intracellular Genomes Direct Weevil Biology: Nuclear, Mitochondrial, Principal Endosymbiont, and Wolbachia. Proc. Natl. Acad. Sci. USA 1999, 96, 6814–6819. [Google Scholar] [CrossRef] [Green Version]
- Zchorifein, E.; Perlman, S.J.; Kelly, S.E.; Katzir, N.; Hunter, M.S. Characterization of a ‘Bacteroidetes’ Symbiont in Encarsia wasps (Hymenoptera: Aphelinidae): Proposal of ‘Candidatus Cardinium Hertigii’. Int. J. Syst. Evol. Microbiol. 2004, 54, 961–968. [Google Scholar] [CrossRef]
- Thao, M.L.; Baumann, P. Evolutionary Relationships of Primary Prokaryotic Endosymbionts of Whiteflies and their Hosts. Appl. Environ. Microbiol. 2004, 70, 3401–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everett, K.D.; Thao, M.; Horn, M.; Dyszynski, G.E.; Baumann, P. Novel Chlamydiae in Whiteflies and Scale Insects: Endosymbionts ‘Candidatus Fritschea bemisiae’ Strain Falk and ‘Candidatus Fritschea eriococci’ Strain Elm. Int. J. Syst. Evol. Microbiol. 2005, 55, 1581–1587. [Google Scholar] [CrossRef] [Green Version]
- Ghanim, M.; Kontsedalov, S. Susceptibility to Insecticides in the Q Biotype of Bemisia tabaci is Correlated with Bacterial Symbiont Densities. Pest Manag. Sci. 2009, 65, 939–942. [Google Scholar] [CrossRef] [PubMed]
- Brumin, M.; Kontsedalov, S.; Ghanim, M. Rickettsia Influences Thermotolerance in the Whitefly Bemisia tabaci B Biotype. Insect Sci. 2011, 18, 57–66. [Google Scholar] [CrossRef]
- Pan, H.P.; Chu, D.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, B.Y.; Zhang, Y.J. Relative Amount of Symbionts in Insect Hosts Changes with Host-Plant Adaptation and Insecticide Resistance. Environ. Entomol. 2013, 42, 74–78. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-∆∆Ct Method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Weibull, J.; Ronquist, F.; Brishammar, S. Free Amino Acid Composition of Leaf Exudates and Phloem Sap: A Comparative Study in Oats and Barley. Plant Physiol. 1990, 92, 222–226. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The Nutritional Quality of Phloem Sap Utilized by Natural Aphid Populations. Ecol. Entomol. 1993, 18, 31–38. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Hansen, A.K.; Moran, N.A. The impact of Microbial Symbionts on Host Plant Utilization by Herbivorous Insects. Mol. Ecol. 2014, 23, 1473–1496. [Google Scholar] [CrossRef] [PubMed]
- Shan, H.W.; Lu, Y.H.; Bing, X.L.; Liu, S.S.; Liu, Y.Q. Differential Responses of the Whitefly Bemisia tabaci Symbionts to Unfavorable Low and High Temperatures. Microb. Ecol. 2014, 68, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A.; Moran, N.A. Horizontal Transfer of Bacterial Symbionts: Heritability and Fitness Effects in a Novel Aphid Host. Appl. Environ. Microbiol. 2005, 71, 7987–7994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, N.A.; Dunbar, H.E. Sexual Acquisition of Beneficial Symbionts in Aphids. Proc. Natl. Acad. Sci. USA 2006, 103, 12803–12806. [Google Scholar] [CrossRef] [Green Version]
- Caspi-Fluger, A.; Inbar, M.; Mozes-Daube, N.; Katzir, N.; Portnoy, V.; Belausov, E.; Hunter, M.S.; Zchori-Fein, E. Horizontal Transmission of the Insect Symbiont Rickettsia is Plant-Mediated. Proc. Biol. Sci. 2012, 279, 1791–1796. [Google Scholar] [CrossRef] [Green Version]
- Gonella, E.; Pajoro, M.; Marzorati, M.; Crotti, E.; Mandrioli, M.; Pontini, M.; Bulgari, D.; Negri, I.; Sacchi, L.; Chouaia, B.; et al. Plant-Mediated Interspecific Horizontal Transmission of an Intracellular Symbiont in Insects. Sci. Rep. 2015, 5, 15811. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Ahmed, M.Z.; Li, S.J.; Lv, N.; Shi, P.Q.; Chen, X.S.; Qiu, B.L. Plant-Mediated Horizontal Transmission of Rickettsia Endosymbiont Between Different Whitefly Species. FEMS Microbiol. Ecol. 2017, 93, fix138. [Google Scholar] [CrossRef] [Green Version]
- Wernegreen, J.J. Mutualism Meltdown in Insects: Bacteria Constrain Thermal Adaptation. Curr. Opin. Microbiol. 2012, 15, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Shan, H.W.; Deng, W.H.; Luan, J.B.; Zhang, M.J.; Zhang, Z.; Liu, S.S.; Liu, Y.Q. Thermal Sensitivity of Bacteriocytes Constrains the Persistence of Intracellular Bacteria in Whitefly Symbiosis under Heat Stress. Environ. Microbiol. Rep. 2017, 9, 706–716. [Google Scholar] [CrossRef]
- Lemoine, M.M.; Engl, T.; Kaltenpoth, M. Microbial Symbionts Expanding or Constraining Abiotic Niche Space in Insects. Curr. Opin. Insect Sci. 2020, 39, 14–20. [Google Scholar] [CrossRef]
- Pan, H.P.; Su, Q.; Jiao, X.G.; Zhou, L.; Liu, B.M.; Xie, W.; Wang, S.L.; Wu, Q.J.; Xu, B.Y.; Zhang, Y.J. Relative Amount of Symbionts in Bemisia tabaci (Gennadius) Q Changes with Host Plant and Establishing the Method of Analyzing Free Amino Acid in B. tabaci. Commun. Integr. Biol. 2013, 6, e23397. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Cao, W.J.; Zhong, L.R.; Godfray, H.C.J.; Liu, X.D. Host Plant Determines the Population Size of an Obligate Symbiont (Buchnera aphidicola) in Aphids. Appl. Environ. Microbiol. 2016, 82, 2336–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, T.L.; Douglas, A.E. Phloem Amino Acids and the Host Plant Range of the Polyphagous Aphid, Aphis. fabae. Entomol. Exp. Appl. 2003, 106, 103–113. [Google Scholar] [CrossRef]
- Kakimoto, K.; Inoue, H.; Yamaguchi, T.; Ueda, S.; Honda, K.i.; Yano, E. Host Plant Effect on Development and Reproduction of Bemisia argentifolii Bellows et Perring (B. tabaci [Gennadius] B-biotype) (Homoptera: Aleyrodidae). Appl. Entomol. Zool. 2007, 42, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.H.; Wang, K.H. Development and Reproduction of Bemisia argentifolii (Homoptera: Aleyrodidae) on Five Host Plants. Environ. Entomol. 1996, 25, 810–816. [Google Scholar] [CrossRef]
- Chandler, S.M.; Wilkinson, T.L.; Douglas, A.E. Impact of Plant Nutrients on the Relationship between a Herbivorous Insect and its Symbiotic Bacteria. Proc. Biol. Sci. 2008, 275, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E.; Minto, L.B.; Wilkinson, T.L. Quantifying Nutrient Production by the Microbial Symbionts in an Aphid. J. Exp. Biol. 2001, 204, 349–358. [Google Scholar]
- Chiel, E.; Inbar, M.; Mozes-Daube, N.; White, J.A.; Hunter, M.S.; Zchori-Fein, E. Assessments of Fitness Effects by the Facultative Symbiont Rickettsia in the Sweetpotato Whitefly (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2009, 102, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Himler, A.G.; Adachi-Hagimori, T.; Bergen, J.E.; Kozuch, A.; Kelly, S.E.; Tabashnik, B.E.; Chiel, E.; Duckworth, V.E.; Dennehy, T.J.; Zchori-Fein, E. Rapid Spread of a Bacterial Symbiont in an Invasive Whitefly is Driven by Fitness Benefits and Female Bias. Science 2011, 332, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B. Introduction to Ecological Biochemistry, 4th ed.; Academic Press: London, UK, 1993; pp. 128–158. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism and Target Gene | Primer Sequences (5′ to 3′) | Product Length (bp) | References |
|---|---|---|---|
| Diagnostic PCR | |||
| Bemisia tabaci mtCOI | C1-J-2195: TTGATTTTTTGGTCATCCAGAAGT L2-N-3014: TCCAATGCACTAATCTGCCATATTA | 800 | [26] |
| Portiera 16S rRNA | Por-F: TGCAAGTCGAGCGGCATCAT Por-R: AAAGTTCCCGCCTTATGCGT | 1000 | [28] |
| Hamiltonella 16S rRNA | Ham-F: TGAGTAAAGTCTGGGAATCTGG Ham-R: CCCGGGAACGTATTCACCGTAG | 1000 | [28] |
| Rickettsia 16S rRNA | Rb-F: GCTCAGAACGAACGCTATC Rb-R: GAAGGAAAGCATCTCTGC | 900 | [29] |
| Wolbachia 16S rRNA | Wol-16S-F: CGGGGGAAAAATTTATTGCT Wol-16S-R: AGCTGTAATACAGAAAGTAAA | 700 | [11,30] |
| Cardinium 16S rRNA | Ch-F: TACTGTAAGAATAAGCACCGGC Ch-R: GTGGATCACTTAACGCTTTCG | 400 | [31] |
| Hemipteriphilus 16S rRNA | Hem-F: GCTCAGAACGAACGCTRKC Hem-R: TTCGCCACTGGTGTTCCTC | 670 | [15] |
| Arsenophonus 23S rRNA | Ars23S-1: CGTTTGATGAATTCATAGTCAAA Ars23S-2: GGTCCTCCAGTTAGTGTTACCCAAC | 900 | [32] |
| Fritschea 23S rRNA | U23F: GATGCCTTGGCATTGATAGGCGATGAAGGA 23SIGR: TGGCTCATCATGCAAAAGGCA | 600 | [11,33] |
| qPCR | |||
| B. tabaci β-actin | qActinF: TCTTCCAGCCATCCTTCTTG qActinR: CGGTGATTT CCTTCTGCATT | 130 | [34,35] |
| Portiera 16S rRNA | qPor-F: TAGTCCACGCTGTAAACG qPor-R: AGGCACCCTTCCATCT | 229 | [36] |
| Hamiltonella 16S rRNA | qHam-F: GCATCGAGTGAGCACAGTTT qHam-R: TATCCTCTCAGACCCGCTAGA | 243 | [35,36] |
| Rickettsia gltA | qgltA-F: AAAGGTTGCTCATCATGCGTT qgltA-R: GCCATAGGATGCGAAGAGCT | 80 | [34,35] |
| Effect | Portiera Amount | |||
|---|---|---|---|---|
| df | Mean Square | F | p | |
| Population (A) | 2 | 2713.206 | 943.388 | <0.001 |
| Host plant (B) | 2 | 5202.316 | 1808.857 | <0.001 |
| Generation (C) | 4 | 77.291 | 26.874 | <0.001 |
| A × B | 4 | 103.224 | 35.891 | <0.001 |
| A × C | 8 | 78.090 | 27.152 | <0.001 |
| B × C | 8 | 75.856 | 26.375 | <0.001 |
| A × B × C | 16 | 62.033 | 21.569 | <0.001 |
| Effect | Hamiltonella Amount | Rickettsia Amount | ||||||
|---|---|---|---|---|---|---|---|---|
| df | Mean Square | F | p | df | Mean Square | F | p | |
| Population (A) | 2 | 29.861 | 28.377 | <0.001 | 2 | 16.368 | 11.706 | <0.001 |
| Host plant (B) | 2 | 3.084 | 2.931 | 0.064 | 2 | 8.526 | 6.098 | 0.005 |
| A × B | 4 | 12.951 | 12.307 | <0.001 | 4 | 18.938 | 13.544 | <0.001 |
| Amino Acid | Plant Leaves | Whitefly | ||
|---|---|---|---|---|
| F2,15 Value | p-Value | F2,24 Value | p-Value | |
| Ala | 30.19 | <0.001 | 12.08 | <0.001 |
| Asp | 23.10 | <0.001 | undetected | |
| Cys | 9.93 | 0.002 | 0.45 | 0.642 |
| Gln | 2.61 | 0.107 | undetected | |
| Glu | 30.52 | <0.001 | 12.85 | <0.001 |
| Gly | undetected | 1.61 | 0.216 | |
| Pro | 9.20 | 0.003 | 6.26 | 0.005 |
| Ser | 27.04 | <0.001 | 32.64 | <0.001 |
| Tyr | 16.12 | <0.001 | 8.32 | 0.001 |
| Arg | 1.40 | 0.277 | 10.88 | <0.001 |
| His | 1.11 | 0.354 | 12.21 | <0.001 |
| Ile | 20.62 | <0.001 | 1.25 | 0.299 |
| Leu | 19.76 | <0.001 | 3.73 | 0.035 |
| Lys | 3.40 | 0.061 | 85.69 | <0.001 |
| Met | 35.09 | <0.001 | 0.44 | 0.646 |
| Phe | 68.71 | <0.001 | 0.02 | 0.984 |
| Thr | 80.39 | <0.001 | 0.54 | 0.588 |
| Trp | 13.22 | <0.001 | 0.12 | 0.890 |
| Val | 2.65 | 0.104 | 1.19 | 0.316 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-H.; Shah, M.M.R.; Song, Y.; Liu, T.-X. Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae). Insects 2020, 11, 501. https://doi.org/10.3390/insects11080501
Liu Y-H, Shah MMR, Song Y, Liu T-X. Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae). Insects. 2020; 11(8):501. https://doi.org/10.3390/insects11080501
Chicago/Turabian StyleLiu, Yan-Hong, M. Mostafizur Rahman Shah, Yue Song, and Tong-Xian Liu. 2020. "Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae)" Insects 11, no. 8: 501. https://doi.org/10.3390/insects11080501
APA StyleLiu, Y. -H., Shah, M. M. R., Song, Y., & Liu, T. -X. (2020). Host Plant Affects Symbiont Abundance in Bemisia tabaci (Hemiptera: Aleyrodidae). Insects, 11(8), 501. https://doi.org/10.3390/insects11080501