How Does the Intricate Mouthpart Apparatus Coordinate for Feeding in the Hemimetabolous Insect Pest Erthesina fullo?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material
2.1. Insect Collecting
2.2. Samples for SEM
2.3. Feeding Behavior on Different Types of Substrates
2.4. Image Processing and Terminology
2.5. Data Analysis
3. Results
3.1. General Morphology and Structure of Mouthparts
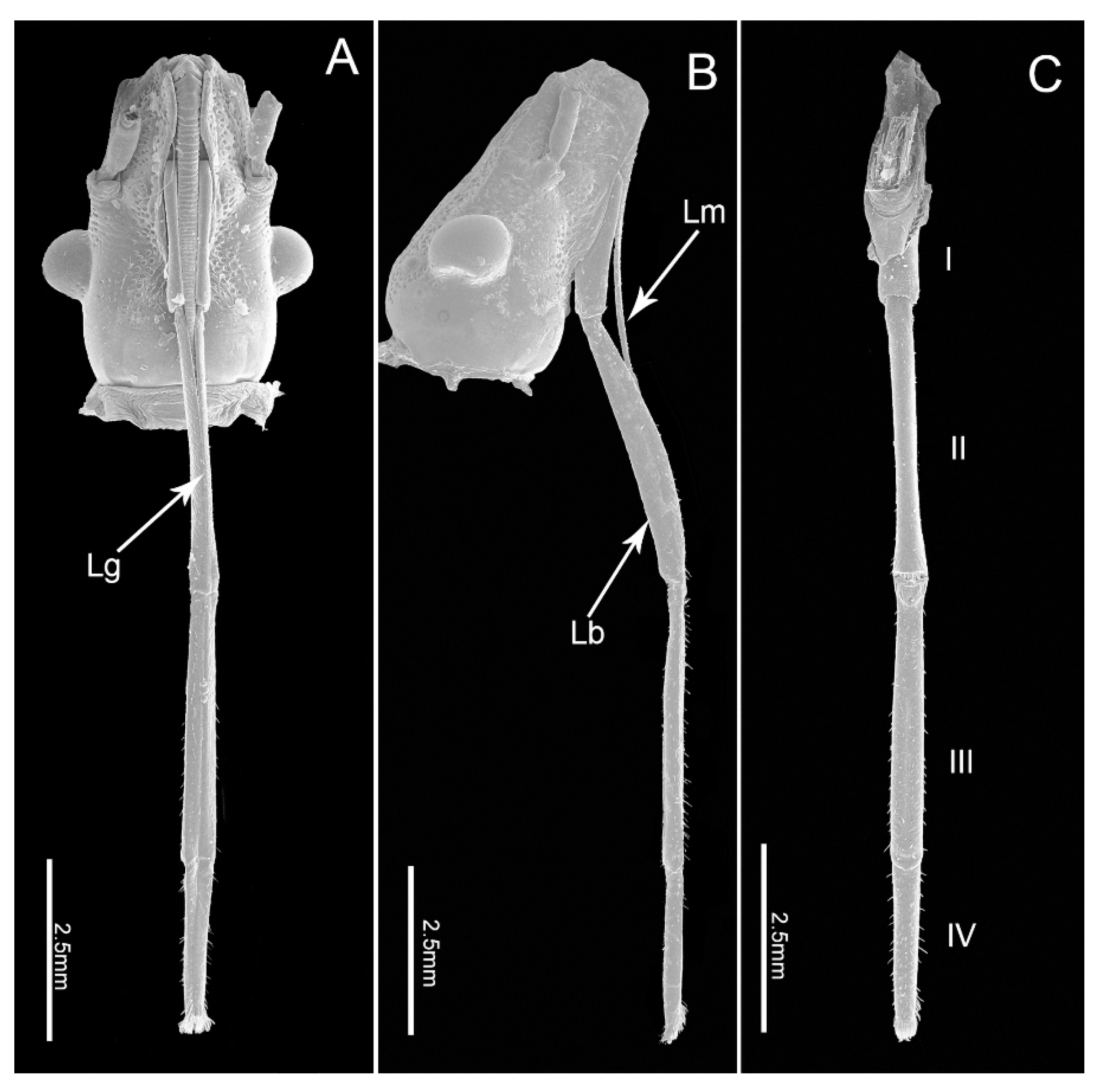
3.1.1. Labrum
3.1.2. Labium
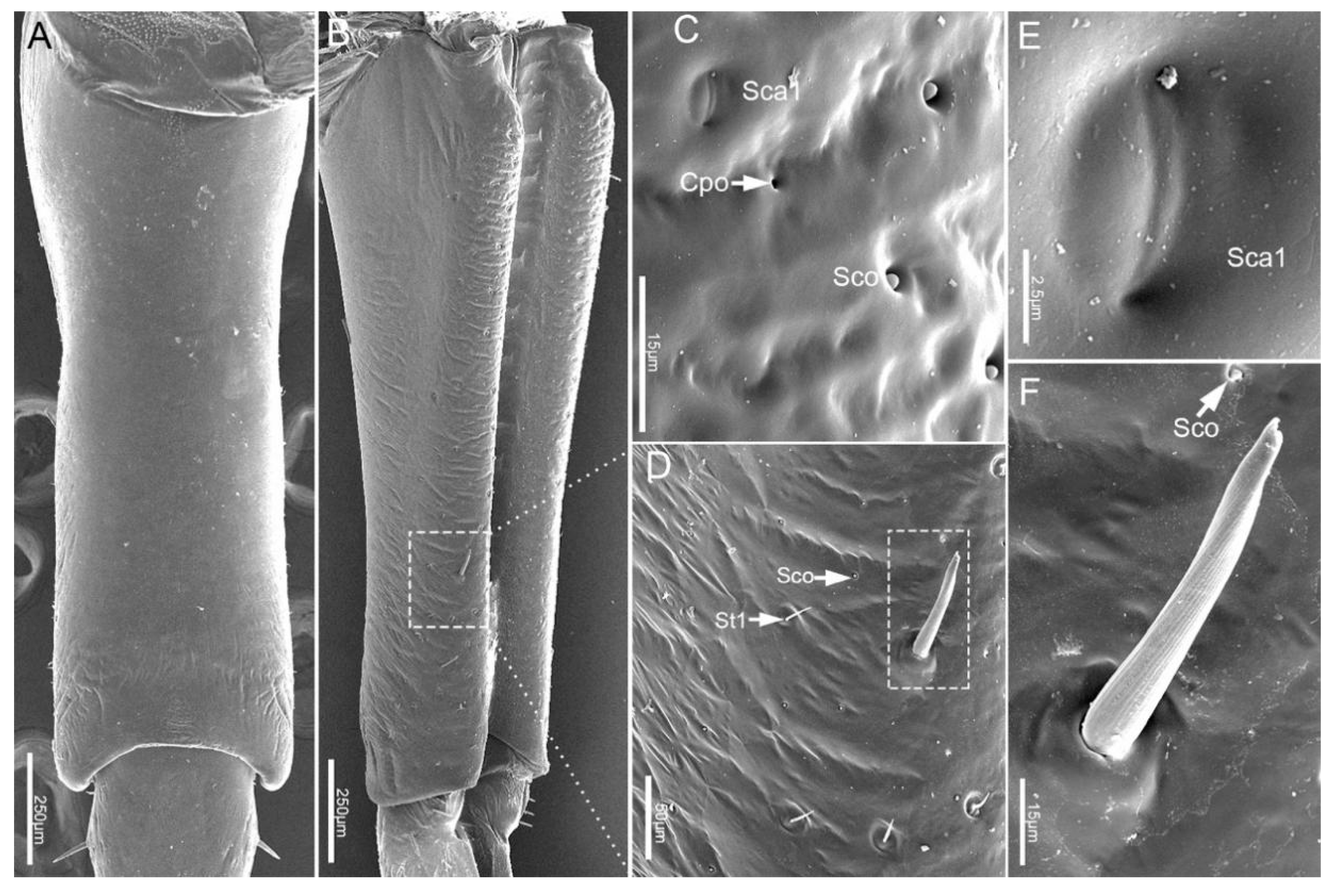
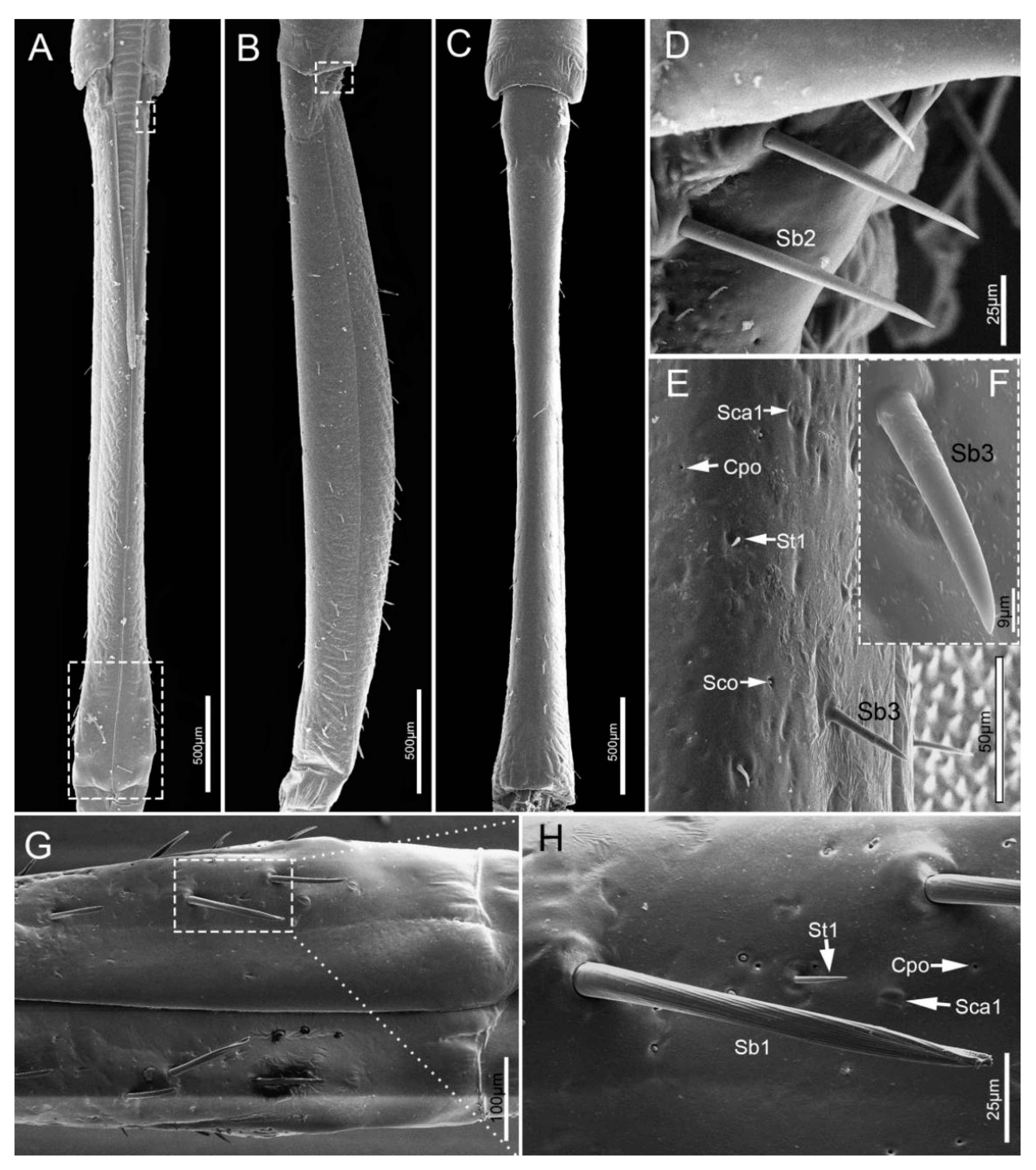
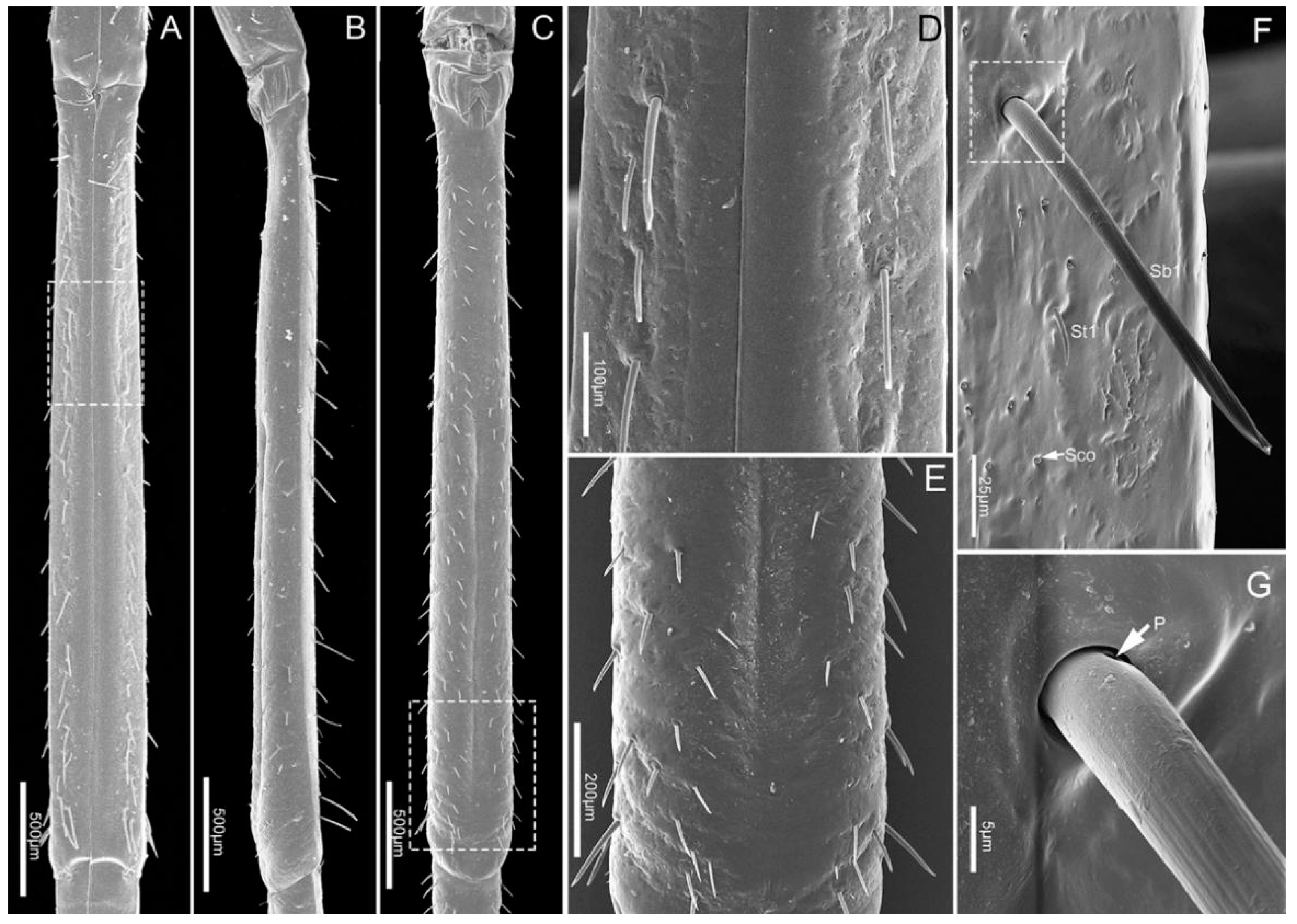
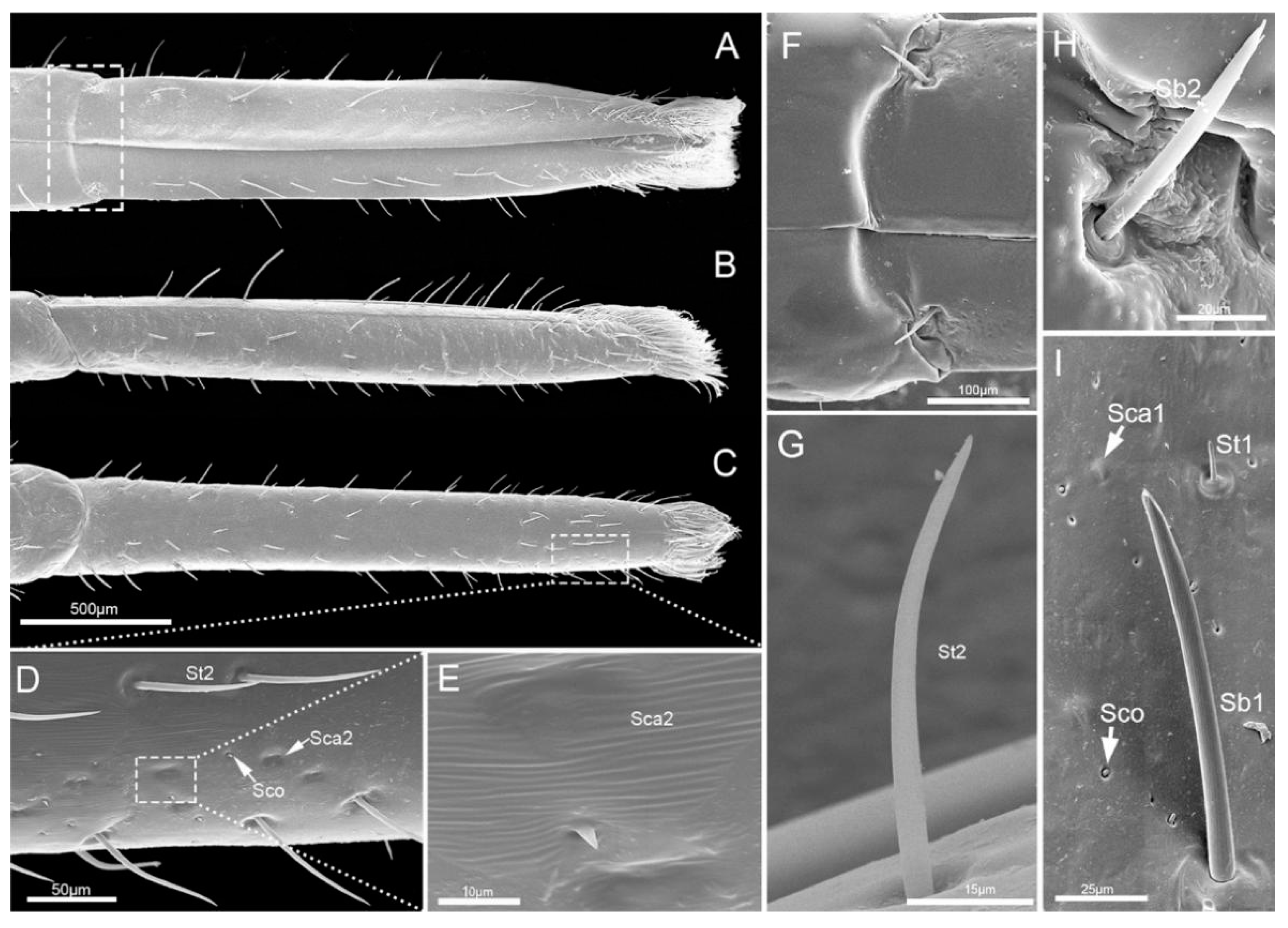
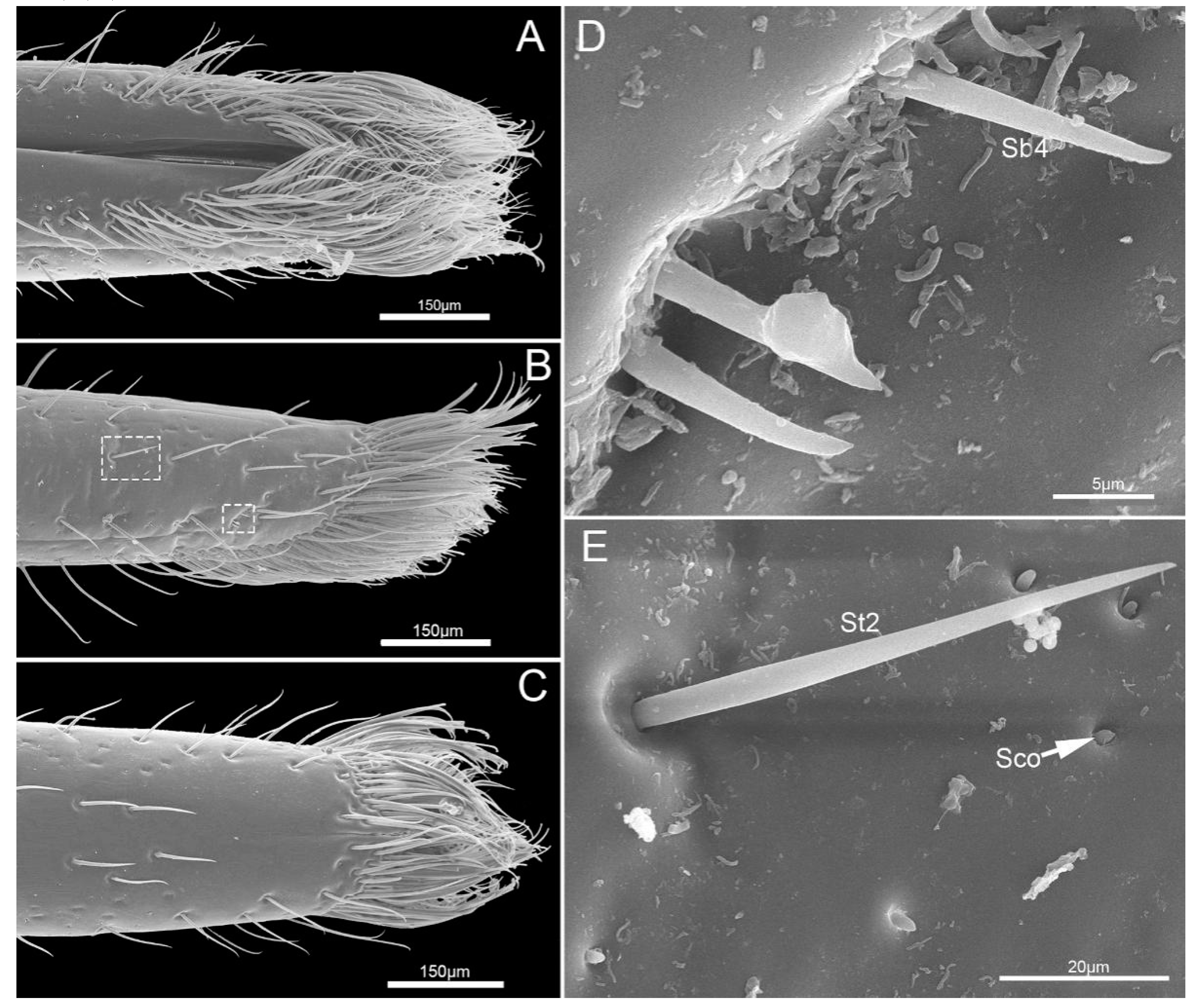
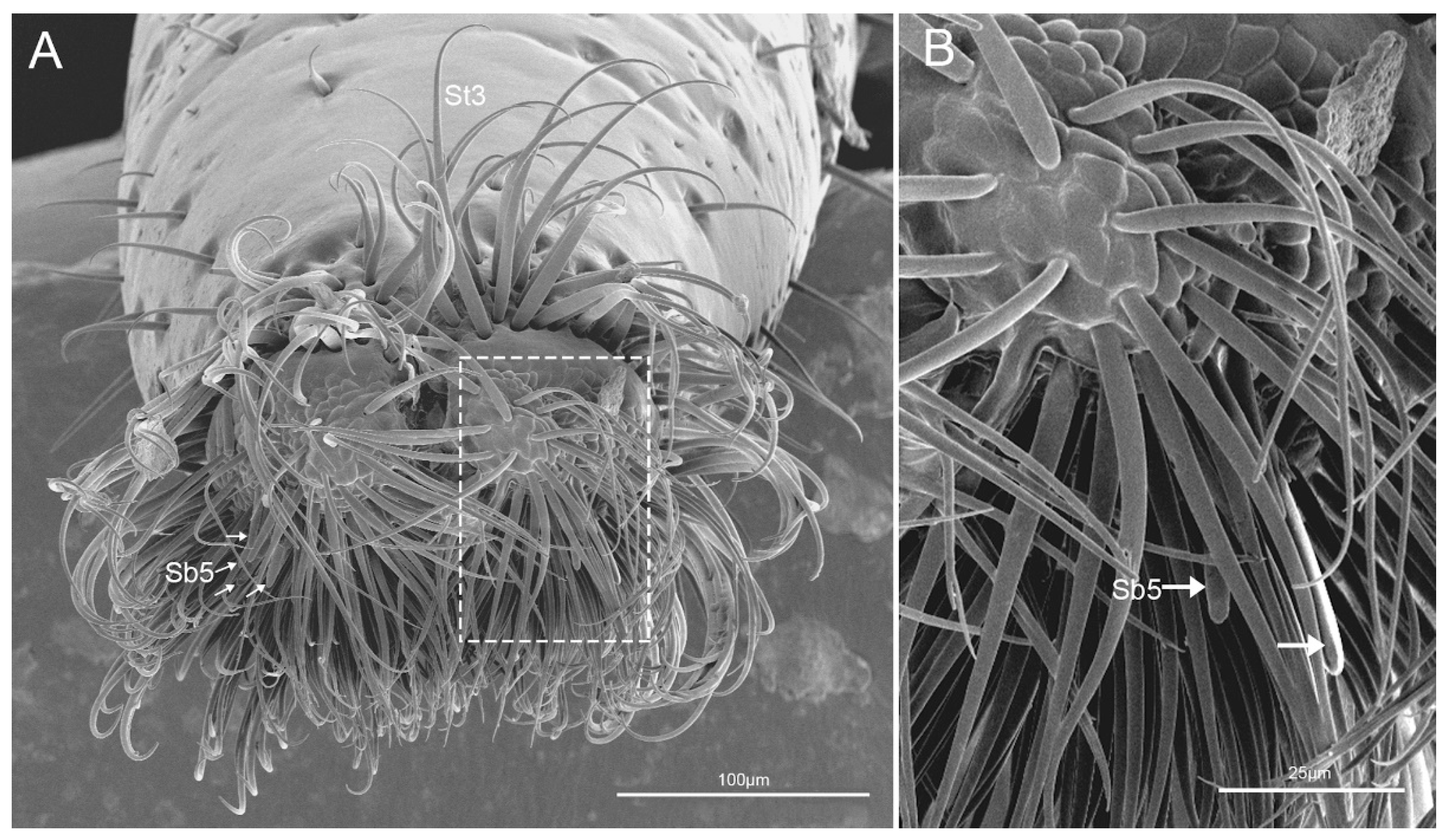
3.1.3. Labial Sensilla Types and Their Arrangement
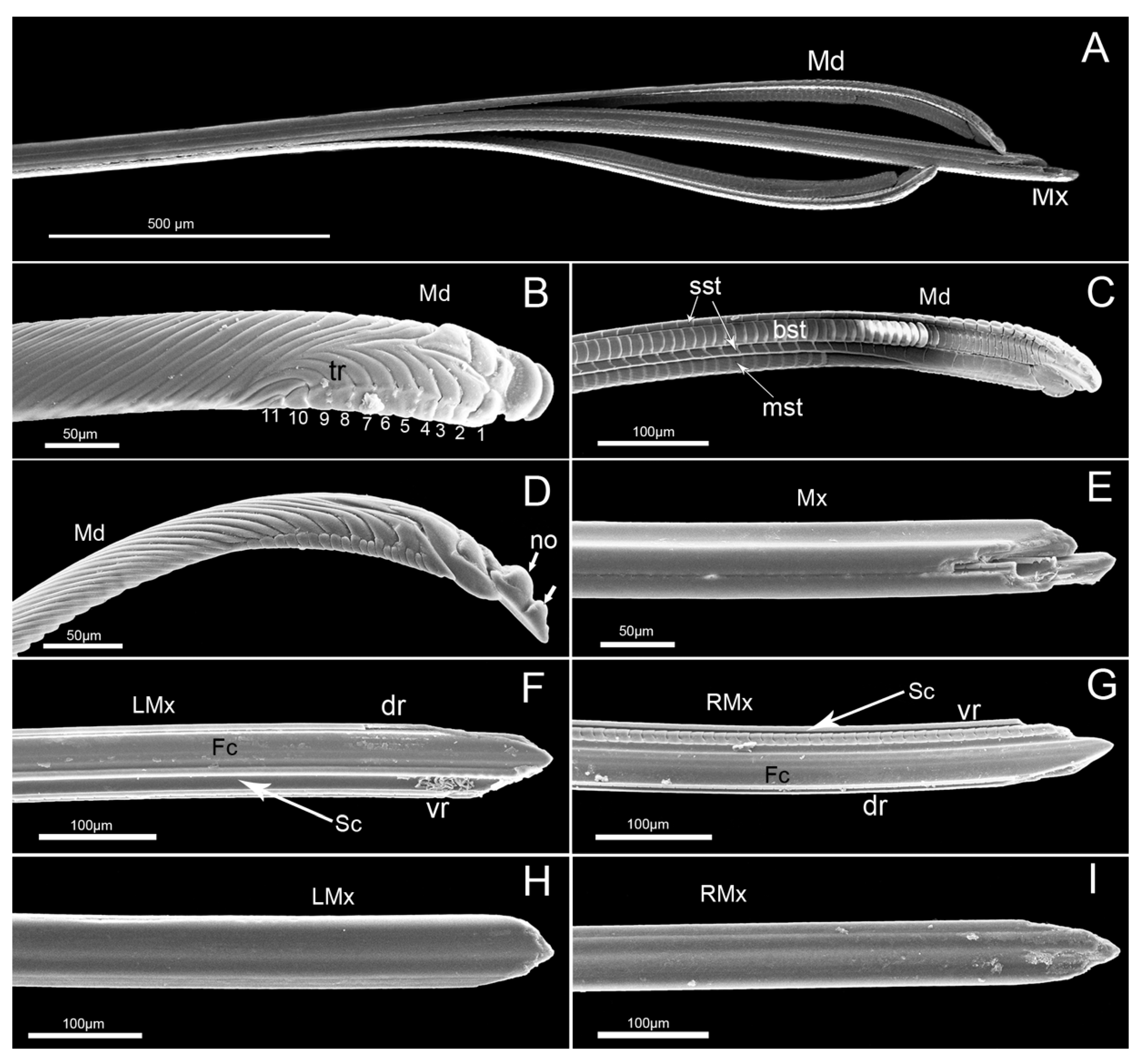
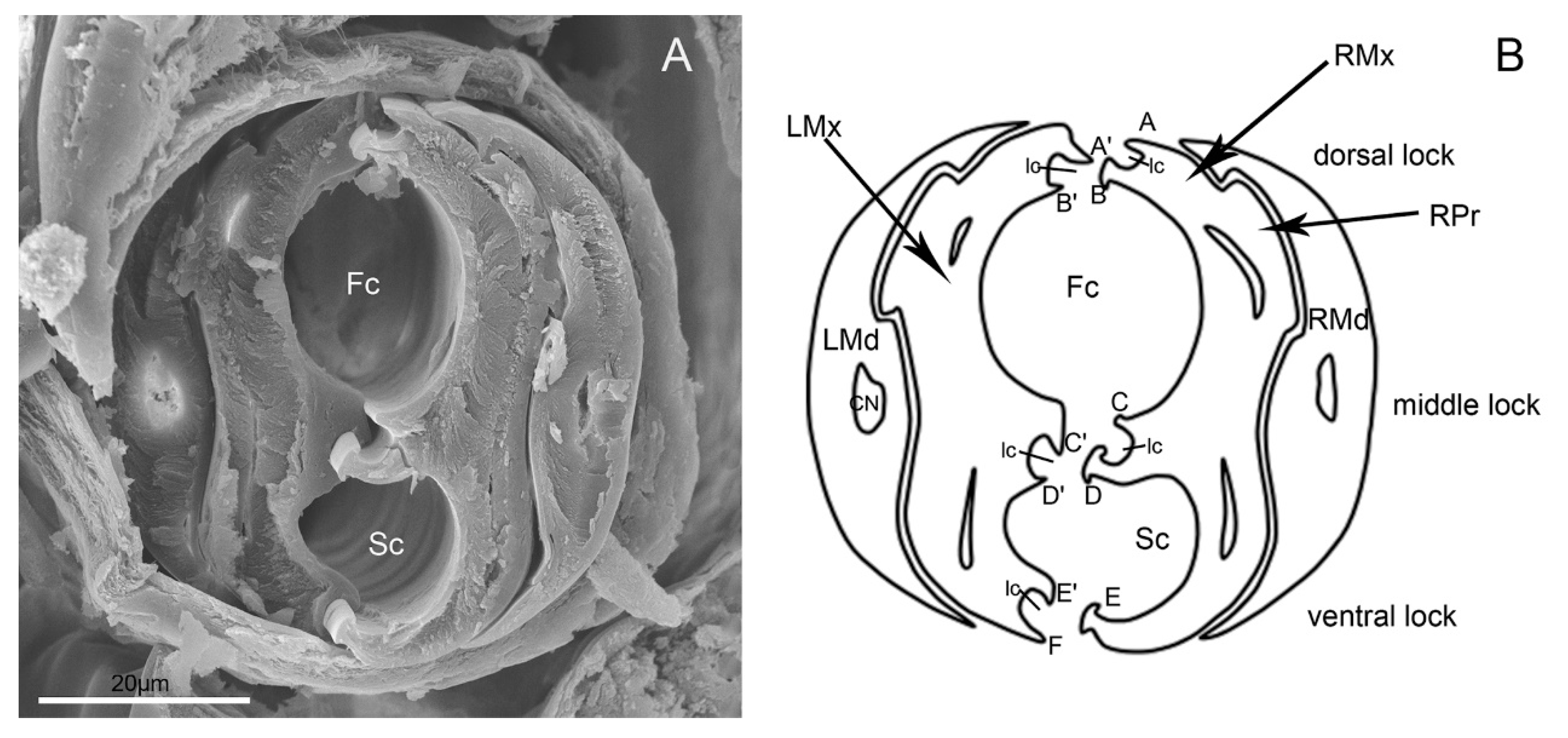
3.2. Stylet Fascicle
3.3. The Process of Feeding by E. fullo
4. Discussion
4.1. Mouthpart Morphology and Their Adaptability to Feeding
4.2. Labial Sensillar System
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marais, E.; Klok, C.J.; Terblanche, J.S.; Chown, S.L. Insect gas exchange patterns: A phylogenetic perspective. J. Exp. Biol. 2005, 208, 4495–4507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weintraub, P.G.; Beanland, L. Insect vectors of phytoplasmas. Annu. Rev. Èntomol. 2006, 51, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Brożek, J.; Bourgoin, T.; Szwedo, J. The interlocking mechanism of maxillae and mandibles in Fulgoroidea (Insecta: Hemiptera: Fulgoromorpha). Polskie Pismo Entomologiczne 2006, 75, 239–253. [Google Scholar]
- Brożek, J.; Bourgoin, T. Morphology and distribution of the external labial sensilla in Fulgoromorpha (Insecta: Hemiptera). Zoomorphology 2012, 132, 33–65. [Google Scholar] [CrossRef] [Green Version]
- Tavella, L.; Arzone, A. Comparative morphology of mouth parts of Zyginidia pullula, Empoasca vitis, and Graphocephala fennahi (Homoptera, Auchenorrhyncha). Ital. J. Zool. 1993, 60, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Leopold, R.A.; Freeman, T.P.; Buckner, J.S.; Nelson, D.R. Mouthpart morphology and stylet penetration of host plants by the glassy-winged sharpshooter, Homalodisca coagulata, (Homoptera: Cicadellidae). Arthropod Struct. Dev. 2003, 32, 189–199. [Google Scholar] [CrossRef]
- Wiesenborn, W.D. Mouth parts and alimentary canal of opsius stactogalus Fieber (Homoptera: Cicadellidae). J. Kans. Èntomol. Soc. 2004, 77, 152–155. [Google Scholar] [CrossRef]
- Zhao, L.; Dai, W.; Zhang, C.; Zhang, Y. Morphological characterization of the mouthparts of the vector leafhopper Psammotettix striatus (L.) (Hemiptera: Cicadellidae). Micron 2010, 41, 754–759. [Google Scholar] [CrossRef]
- Ammar, E.-D.; Hall, D. New and simple methods for studying hemipteran stylets, bacteriomes, and salivary sheaths in host plants. Ann. Èntomol. Soc. Am. 2012, 105, 731–739. [Google Scholar] [CrossRef]
- Pollard, D.G. Some aspects of plant penetration by Myzits persicae (Sulz.) nymphs (Homoptera, Aphididae). Bull. Entomol. Res. 1971, 61, 315–324. [Google Scholar] [CrossRef]
- Pollard, D.G. Plant penetration by feeding aphids (Hemiptera, Aphidoidea): A review. Bull. Èntomol. Res. 1973, 62, 631–714. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Mechanoreceptors of the aphid labium. Èntomol. Exp. Appl. 1978, 24, 731–737. [Google Scholar] [CrossRef]
- Uzest, M.; Gargani, D.; Dombrovsky, A.; Cazevieille, C.; Cot, D.; Blanc, S. The “acrostyle”: A newly described anatomical structure in aphid stylets. Arthropod Struct. Dev. 2010, 39, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Kaushik, S.; Ramamurthy, V.; Lakhanpaul, S.; Ramani, R.; Sharma, K.; Vidyarthi, A. Mouthparts and stylet penetration of the lac insect Kerria lacca (Kerr) (Hemiptera:Tachardiidae). Arthropod Struct. Dev. 2012, 41, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.P.; Gordh, G. The occurrence of apical labial sensilla in the Aleyrodidae and evidence for a contact chemosensory function. Èntomol. Exp. Appl. 1989, 51, 215–224. [Google Scholar] [CrossRef]
- Rosell, R.C.; Lichty, J.E.; Brown, J.K. Ultrastructure of the mouthparts of adult sweet potato whitefly, Bemisia tabaci Gennadius (Homoptera: Aleyrodidae). Int. J. Insect Morphol. Embryol. 1995, 24, 297–306. [Google Scholar] [CrossRef]
- Cobben, R.H. Evolutionary trends in Heteroptera. In Part II Mouthpart-Structures and Feeding Strategies; Mededelingen Landbouwhogeschool: Wageningen, The Netherlands, 1978; Volume 78, pp. 1–407. [Google Scholar]
- Boyd, D.W. Digestive enzymes and stylet morphology of Deraeocoris nigritulus (Uhler) (Hemiptera: Miridae) reflect adaptations for predatory habits. Ann. Èntomol. Soc. Am. 2003, 96, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Anderson, W.G.; Hengmoss, T.M.; Baxendale, F.P.; Baird, L.M.; Sarath, G.; Higley, L. Chinch bug (Hemiptera: Blissidae) mouthpart morphology, probing frequencies, and locations on resistant and susceptible germplasm. J. Econ. Entomol. 2006, 99, 212–221. [Google Scholar] [CrossRef]
- Kumar, S.M.; Sahayaraj, K. Gross morphology and histology of head and salivary apparatus of the predatory bug, Rhynocoris marginatus. J. Insect Sci. 2012, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Garzo, E.; Bonani, J.; Lopes, J.R.S.; Fereres, A. Morphological description of the mouthparts of the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Arthropod Struct. Dev. 2012, 41, 79–86. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. Morphology and distribution of antennal sense organs and diversity of mouthpart structures in Odontopus nigricornis (Stål) and Nezara viridula L. (Hemiptera). Int. J. Insect Morphol. Embryol. 1995, 24, 119–132. [Google Scholar] [CrossRef]
- Freeman, T.P.; Buckner, J.S.; Nelson, D.R. Stylet length of whitefly adults and nymphs and the mechanism of stylet insertion into the leaves of host plants. Microsc. Microanal. 2000, 6 (Suppl. 2), 876–877. [Google Scholar] [CrossRef]
- Freeman, T.P.; Buckner, J.S.; Nelson, D.R.; Chu, C.C.; Henneberry, T.J. Stylet penetration by Bemisia argentifolii (Homoptera: Aleyrodidae) into host leaf tissue. Ann. Entomol. Soc. Am. 2001, 94, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Boyd, D.W.; Cohen, A.C.; Alverson, D.R. Digestive enzymes and stylet morphology of Deraeocoris nebulosus (Hemiptera: Miridae), a predacious plant bug. Ann. Èntomol. Soc. Am. 2002, 95, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Rani, P.U. Sensillary morphology on the rostral apex and their possible role in prey location behaviour of the carnivorous stinkbug, Eocanthecona furcellata (Wolff) (Heteroptera: Pentatomidae). Acta Zool. 2009, 90, 246–253. [Google Scholar] [CrossRef]
- Esquivel, J.F. Estimating potential stylet penetration of southern green stink bug—A mathematical modeling approach. Èntomol. Exp. Appl. 2011, 140, 163–170. [Google Scholar] [CrossRef]
- Backus, E.A. Sensory systems and behaviours which mediate hemipteran plant-feeding: A taxonomic overview. J. Insect Physiol. 1988, 34, 151–165. [Google Scholar] [CrossRef]
- Strong, F.E. Physiology of Injury Caused by Lygus hesperus. J. Econ. Entomol. 1970, 63, 808–814. [Google Scholar] [CrossRef]
- Tingey, W.M.; Pillimer, E.A. Lygus Bugs: Crop Resistance and Physiological Nature of Feeding Injury. Bull. Entomol. Soc. Amer. 1977, 23, 277–287. [Google Scholar] [CrossRef]
- Hori, K. Possible causes of disease symptoms resulting from the feeding of phytophagous heteroptera, Chapter, 2. In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 11–35. [Google Scholar]
- Mitchell, P.L. Heteroptera as vectors of plant pathogens. Neotropical Èntomol. 2004, 33, 519–545. [Google Scholar] [CrossRef] [Green Version]
- Panizzi, A.R.; McPherson, J.E.; James, D.G.; Javahery, M.; McPherson, R.M. Stink bugs (Pentatomidae), Chapter, 13. In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 421–474. [Google Scholar]
- Brożek, J. Labial sensillae and the internal structure of the mouthparts of Xenophyes cascus (Bergroth 1924) (Peloridiidae: Coleorrhyncha: Hemiptera) and their significance in evolutionary studies on the Hemiptera. Aphids Hemipterous Insects 2007, 13, 35–42. [Google Scholar]
- Brożek, J. Morphology and arrangement of the labial sensilla of the water bugs. Bull. Insectol. 2008, 61, 67–168. [Google Scholar]
- Baker, G.T.; Chen, X.; Ma, P.W. Labial tip sensilla of Blissus leucopterus (Hemiptera: Blissidae): Ultrastructure and behavior. Insect Sci. 2008, 15, 271–275. [Google Scholar] [CrossRef]
- Brożek, J.; Herczek, A. Internal structure of the mouthparts of true bugs (Hemiptera: Heteroptera). Pol. J. Entomol. 2004, 73, 79–106. [Google Scholar]
- Li, Q.C.; Cheng, A.Y.; Wang, H.S.; Zhang, W.Y. Control technicians of the Halyomorpha picus (Fabricius) and Erthesina fullo (Thunberg). Plant Doctor 1998, 11, 17–18. (In Chinese) [Google Scholar]
- Mi, Q.Q.; Zhang, J.P.; Gould, E.; Chen, J.H.; Sun, Z.T.; Zhang, F. Biology, Ecology, and Management of Erthesina fullo (Hemiptera: Pentatomidae): A Review. Insects 2020, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- Song, H.W.; Wang, C.M. Studies on the harm and prevention of the Halyomorpha picus (Fabricius) and Erthesina fullo (Thunberg) to Chinese jujube. Chin. J. Appl. Entomol. 1993, 30, 225–228. (In Chinese) [Google Scholar]
- Sun, S.X. Studies on the alimentary canal of Erthesina fullo Thunberg. J. Shandong Agric. Coll. 1956, 2, 37–50. (In Chinese) [Google Scholar]
- Zhang, C.T.; Li, D.L.; Su, H.F.; Xu, G.L. A study on the biological characteristics of Halyomorpha picus and Erthesina fullo. For. Res. 1993, 6, 271–275. (In Chinese) [Google Scholar]
- Altner, H.; Prillinger, L. Ultrastructure of invertebrate chemo, thermo, and hygroreceptors and its functional significance. Int. Rev. Cytol. 1980, 67, 69–139. [Google Scholar] [CrossRef]
- Parveen, S.; Ahmad, A.; Brożek, J.; Ramamurthy, V.V. Morphological diversity of the labial sensilla of phytophagous and predatory Pentatomidae (Hemiptera: Heteroptera), with reference to their possible functions. Zootaxa 2015, 4039, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Backus, E.A. Anatomical and sensory mechanisms of leafhopper and planthopper feeding behavior. In The Leafhoppers and Planthoppers; Nault, L.R., Rodriguez, J.G., Eds.; John Wiley & Sons: New York, NY, USA, 1985; pp. 163–194. [Google Scholar]
- Feir, D.; Beck, S.D. Feeding behavior of the large milkweed bug, Oncopellus fasciatus. Ann. Èntomol. Soc. Am. 1963, 56, 224–229. [Google Scholar] [CrossRef]
- Saxena, K.N. Mode of ingestion in a heteropteran insect Dysdercus koenigii (F.) (Pyrrhocoridae). J. Insect Physiol. 1963, 9, 47–71. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. External morphology of antennal and rostral sensillae in four hemipteran insects and their possible role in host plant selection. Int. J. Trop. Insect Sci. 2005, 25, 198–207. [Google Scholar] [CrossRef]
- Depieri, R.A.; Panizzi, A.R. Rostrum length, mandible serration, and food and salivary canals areas of selected species of stink bugs (Heteroptera, Pentatomidae). Revista Brasileira De Entomologia 2010, 54, 584–587. [Google Scholar] [CrossRef] [Green Version]
- Depieri, R.A.; Siqueira, F.; Panizzi, A.R. Aging and food source effects on mandibular stylets teeth wear of Phytophagous Stink Bug (Heteroptera: Pentatomidae). Neotrop. Èntomol. 2011, 39, 952–956. [Google Scholar] [CrossRef] [Green Version]
- Barsagade, D.; Gathalkar, G. First predation record of Canthecona furcellata(Wolff.) (Hemiptera: Pentatomidae) on spinning stage silkworm Antheraea mylitta(Drury). Èntomol. Res. 2016, 46, 236–245. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. Fine structure of mouthparts and feeding performance of Pyrrhocoris sibiricus Kuschakevich with remarks on the specialization of sensilla and stylets for seed feeding. PLoS ONE 2017, 12, e0177209. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Dai, W. Fine Morphology of the mouthparts in Cheilocapsus nigrescens (Hemiptera: Heteroptera: Miridae) reflects adaptation for phytophagous habits. Insects 2019, 10, 143. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Brożek, J.; Dai, W. Sensory armature and stylets of the mouthparts of Stephanitis nashi (Hemiptera: Cimicomorpha: Tingidae), their morphology and function. Micron 2020, 132, 102840. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.; Dai, W. Morphological disparity of the mouthparts in polyphagous species of Largidae (Heteroptera: Pentatomomorpha: Pyrrhocoroidea) reveals feeding specialization. Insects 2020, 11, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, R.; Perrimon, N. The molecular genetics of head development in Drosophila melanogaster. Development 1991, 112, 899–912. [Google Scholar] [PubMed]
- Popadić, A.; Panganiban, G.; Rusch, D.; Shear, W.A.; Kaufman, T.C. Molecular evidence for the gnathobasic derivation of arthropod mandibles and for the appendicular origin of the labrum and other structures. Dev. Genes Evol. 1998, 208, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Boyan, G.; Williams, J.; Posser, S.; Bräunig, P. Morphological and molecular data argue for the labrum being non-apical, articulated, and the appendage of the intercalary segment in the locust. Arthropod Struct. Dev. 2002, 31, 65–76. [Google Scholar] [CrossRef]
- Spooner, C.S. The phylogeny of the Hemiptera based on a study of the head capsule. Illinois Biol. Monogr. 1938, 16, 1–102. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.P. A consideration of the systematic position of the Saldidae and the Mesoveliidae (Hemiptera: Heteroptera). Proc. Entomol. Soc. Wash. 1963, 65, 31–38. [Google Scholar]
- Štys, P. On the morphology of the labrum in Heteroptera. Acta Entomol. Bohemoslov. 1969, 66, 150–158. [Google Scholar]
- Forthman, M.; Weirauch, C. Phylogenetics and biogeography of the endemic Madagascan millipede assassin bugs (Hemiptera: Reduviidae: Ectrichodiinae). Mol. Ph. Evol. 2016, 100, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Forthman, M.; Weirauch, C. Millipede assassins and allies (Heteroptera: Reduviidae: Ectrichodiinae, Tribelocephalinae): Total evidence phylogeny, revised classification and evolution of sexual dimorphism. Syst. Èntomol. 2017, 42, 575–595. [Google Scholar] [CrossRef]
- Weber, H. Zur vergleichenden Physiologie der Saugorgane der Hemipteren. Zeitschrift für vergleichende Physiologie 1928, 8, 145–186. [Google Scholar] [CrossRef]
- Snodgrass, R.E. Principles of Insect Morphology; McGraw Hill Book Co.: New York, NY, USA, 1935. [Google Scholar]
- Esquivel, J.F. Stylet penetration estimates for a suite of phytophagous hemipteran pests of row crops. Environ. Èntomol. 2015, 44, 619–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faucheux, M.M. Relations entre l’ultrastructure des stylets manibulaires et maxillaires et la prise de nourriture chez les insects Hemipteres. CR Acad. Sci. Paris (Ser. D) 1975, 281, 41–44. [Google Scholar]
- Cohen, A.C. Feeding adaptations of some predaceous Hemiptera. Ann. Èntomol. Soc. Am. 1990, 83, 1215–1223. [Google Scholar] [CrossRef]
- Swart, C.C.; Felgenhauer, B.E. Structure and function of the mouthparts and salivary gland complex of the giant Waterbug, Belostoma lutarium (Stål) (Hemiptera: Belostomatidae). Ann. Èntomol. Soc. Am. 2003, 96, 870–882. [Google Scholar] [CrossRef]
- Roitberg, B.D.; Gillespie, D.R.; Quiring, D.M.J.; Alma, C.R.; Jenner, W.H.; Perry, J.; Peterson, J.H.; Salomon, M.; VanLaerhoven, S. The cost of being an omnivore: Mandible wear from plant feeding in a true bug. Naturwissenschaften 2005, 92, 431–434. [Google Scholar] [CrossRef]
- Romani, R.; Salerno, G.; Frati, F.; Conti, E.; Isidoro, N.; Bin, F. Oviposition behaviour in Lygus rugulipennis: A morpho-functional study. Èntomol. Exp. Appl. 2005, 115, 17–25. [Google Scholar] [CrossRef]
- Bérenger, J.; Pluot-Sigwalt, D. Notes sur Micrauchenus lineola (Fabricius 1787), espÈcetermitophile et termitophage (Heteroptera: Reduviidae: Harpactorinae, Apiomerini). Ann. Soc. Entomol. Fr. 2009, 45, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Sahayaraj, K.; Kanna, A.V.; Kumar, S.M. Gross morphology of feeding canal, salivary apparatus and digestive enzymes of salivary gland of Catamirus brevipennis (Serville ) (Hemiptera: Reduviidae). J. Entomol. Res. Soc. 2010, 12, 37–50. [Google Scholar]
- Brożek, J. A comparison of external and internal maxilla and mandible morphology of water bugs (Hemiptera: Heteroptera: Nepomorpha). Zootaxa 2013, 3635, 340–378. [Google Scholar] [CrossRef]
- Stubbins, F.L.; Mitchell, P.L.; Turnbull, M.W.; Reay-Jones, F.P.F.; Greene, J.K. Mouthpart morphology and feeding behavior of the invasive kudzu bug, Megacopta cribraria (Hemiptera: Plataspidae). Invertebr. Boil. 2017, 136, 309–320. [Google Scholar] [CrossRef]
- Cohen, A.C. Feeding fitness and quality of domesticated and feral predators: Effects of long-term rearing on artificial diet. Boil. Control. 2000, 17, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Miles, P.W. Interaction of plant phenols and salivary phenolases in the relationship between plants and hemiptera. Èntomol. Exp. Appl. 1969, 12, 736–744. [Google Scholar] [CrossRef]
- Miles, P. Contact chemoreception in some Heteroptera, including chemoreception internal to the stylet food canal. J. Insect Physiol. 1958, 2, 338–347. [Google Scholar] [CrossRef]
- Backus, E.A.; McLean, D.L. The sensory systems and feeding behavior of leafhoppers. I. The aster leafhopper, Macrosteles fascifronsstål (homoptera, cicadellidae). J. Morphol. 1982, 172, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.; Goodman, L.J.; Duckett, J.G. Ultrastructure of sensory receptors on the labium of the rice brown planthopper. Cell Tissue Res. 1983, 230, 353–366. [Google Scholar] [CrossRef]
- Frazier, J.L. Nervous system: Sensory system. In Fundamentals of Insect Physiology; Blum, M.S., Ed.; John Wiley & Sons: New York, NY, USA, 1985; pp. 287–356. [Google Scholar]
- Peregrine, D. Fine structure of sensilla basiconica on the labium of the cotton stainer, Dysdercus fasciatus (signoret) (Heteroptera: Pyrrhocoridae). Int. J. Insect Morphol. Embryol. 1972, 1, 241–251. [Google Scholar] [CrossRef]
- Hatfield, L.D.; Frazier, J.L. Ultrastructure of the labial tip sensilla of the tarnished plant bug, Lygus lineolaris (P. de Beauvois) (Hemiptera: Miridae). Int. J. Insect Morphol. Embryol. 1980, 9, 59–66. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Position | Length (μm) | Width (μm) | N |
|---|---|---|---|---|
| Male | Lm | 4687.3 ± 310.1 | 6 | |
| Lb | 11503.3 ± 123.9 | 6 | ||
| Lb1 | 2013.0 ± 43.4 | 586.7 ± 7.9 | 6 | |
| Lb2 | 3946.8 ± 70.7 | 282.2 ± 5.3 | 6 | |
| Lb3 | 3491.6 ± 118.0 | 384.4 ± 5.7 | 6 | |
| Lb4 | 2366.7 ± 59.1 | 290.2 ± 7.6 | 6 | |
| Female | Lb | 12952.5 ± 75.5 | 5 | |
| Lb1 | 2359.3 ± 22.6 | 629.0 ± 15.5 | 5 | |
| Lb2 | 3771.6 ± 124.4 | 286.3 ± 5.3 | 5 | |
| Lb3 | 4055.9 ± 55.0 | 410.0 ± 6.0 | 5 | |
| Lb4 | 2579.4 ± 19.5 | 315.0 ± 3.5 | 5 |
| Type | Location on Mouthparts | Length (μm) | Basal Diameter (μm) | N | Shape | Socket | Surface | Pore | Category | Function |
|---|---|---|---|---|---|---|---|---|---|---|
| St1 | Lm, Lb1–4 | 12.0 ± 1.7 | 1.6 ± 0.2 | 10 | Hair | Flexible | Smooth | No | Mechanoreceptive sensilla | Tactile |
| St2 | Lb4 | 79.8 ± 1.9 | 4.92 ± 0.2 | 7 | Hair | Flexible | Smooth | No | Mechanoreceptive sensilla | Tactile |
| St3 | Lb4 | 83.1 ± 6.3 | 4.2 ± 0.6 | 16 | Hair | Inflexible | Smooth | No | Mechanoreceptive sensilla | Tactile |
| Sb1 | Lm, Lb1–4 | 89.0 ± 15.4 | 7.4 ± 1.2 | 20 | Hair | Flexible | Grooved | No | Mechanoreceptive sensilla | Tactile |
| Sb2 | Lb2, Lb 4 | 86.3 ± 7.1 | 6.5 ± 0.7 | 6 | Peg | Flexible | Smooth | Wp (Uniporous) | Proprioceptive sensilla | Perceive the degree of flexion of the joint |
| Sb3 | Lb2 | 55.8 ± 3.3 | 6.8 ± 0.4 | 16 | Peg in pit | Inflexible | Smooth | No | Thermo-hygroreceptive sensilla | Temperature/humidity |
| Sb4 | Lb4 | 12.8 ± 0.6 | 1.9 ± 0.1 | 6 | Peg in pit | Inflexible | Smooth | No | Thermo-hygroreceptive sensilla | Temperature/humidity |
| Sb5 | SF | Longer than Sb1 | Wider than Sb1 | Peg | Flexible | Smooth | Tp | Chemoreceptive sensilla | Gustatory | |
| Sco | Lm, Lb1–4 | 2.4 ± 0.4 | 10 | Pegs in cavity | Inflexible | Smooth | No | Thermo-hygroreceptive sensilla | Temperature/humidity | |
| Sca1 | Lb1, 2, 4 | 6.7 ± 0.4 | 4 | Oval plate | Inflexible | Smooth | No | Proprioceptive sensilla | Perceive the degree of flexion of the joint | |
| Sca2 | Lb4 | 6.2 ± 1.2 | 4 | Oval plate | Inflexible | Smooth | No | Proprioceptive sensilla | Perceive the degree of flexion of the joint |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Dai, W. How Does the Intricate Mouthpart Apparatus Coordinate for Feeding in the Hemimetabolous Insect Pest Erthesina fullo? Insects 2020, 11, 503. https://doi.org/10.3390/insects11080503
Wang Y, Dai W. How Does the Intricate Mouthpart Apparatus Coordinate for Feeding in the Hemimetabolous Insect Pest Erthesina fullo? Insects. 2020; 11(8):503. https://doi.org/10.3390/insects11080503
Chicago/Turabian StyleWang, Yan, and Wu Dai. 2020. "How Does the Intricate Mouthpart Apparatus Coordinate for Feeding in the Hemimetabolous Insect Pest Erthesina fullo?" Insects 11, no. 8: 503. https://doi.org/10.3390/insects11080503
APA StyleWang, Y., & Dai, W. (2020). How Does the Intricate Mouthpart Apparatus Coordinate for Feeding in the Hemimetabolous Insect Pest Erthesina fullo? Insects, 11(8), 503. https://doi.org/10.3390/insects11080503