Characterization of Sodium Channel Mutations in the Dengue Vector Mosquitoes Aedes aegypti and Aedes albopictus within the Context of Ongoing Wolbachia Releases in Kuala Lumpur, Malaysia

 , , ,
, , ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
- Monitor insecticide resistance in Aedes aegypti in the vicinity of Wolbachia program release sites in Shah Alam, Selangor (30 km from Kuala Lumpur) and look for changes in the genetic constitution of resistance at those sites.
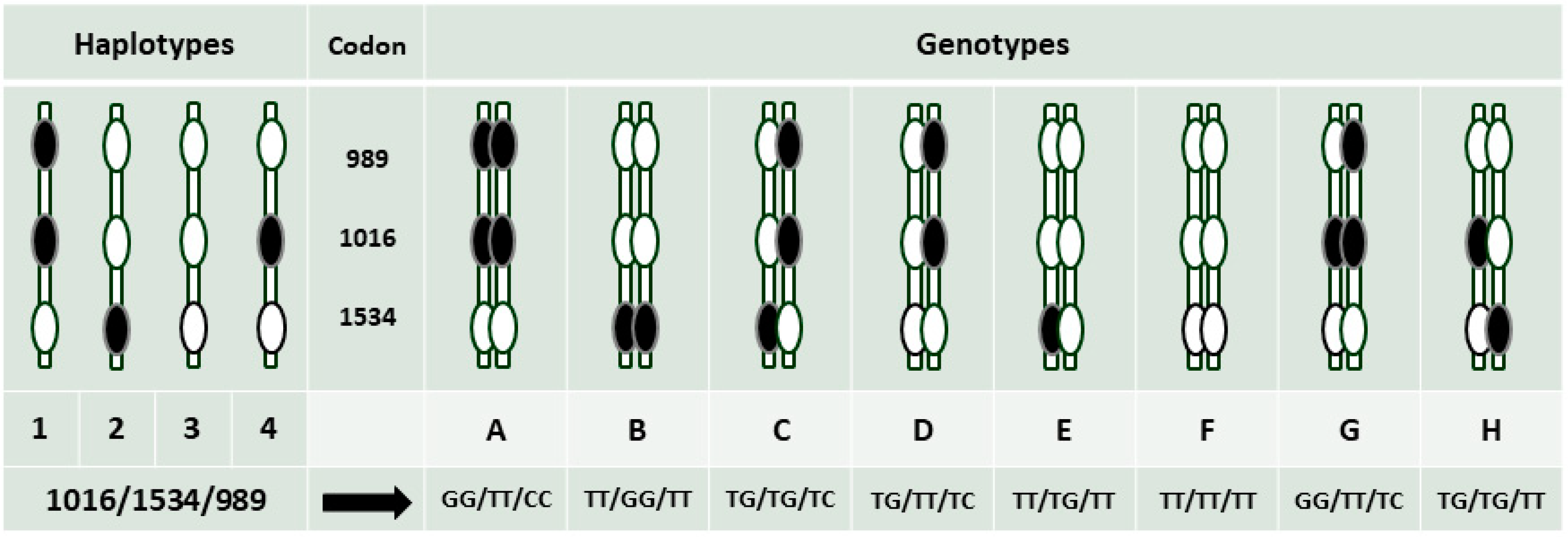
- Characterize the genotypes at three loci in the sodium channel for the V1016G, F1534C and S989P mutations and assess the association between the mutations and pyrethroid resistance in Ae. aegypti in Shah Alam.
- Assess the utility of backcrossing laboratory colony Wolbachia mosquitoes to the field population to maintain consistency in sodium channel mutations and enhance survival of the released mosquitoes.
- Sequence the region of the sodium channel gene around codon 1534 in Ae. albopictus (a second vector of dengue which may be the target of future Wolbachia release programs in Malaysia) from AU2, Keramat, Selangor (Kuala Lumpur boundary) to look for resistance mutations which have been identified in some populations, though not yet from Malaysia.
2. Materials and Methods
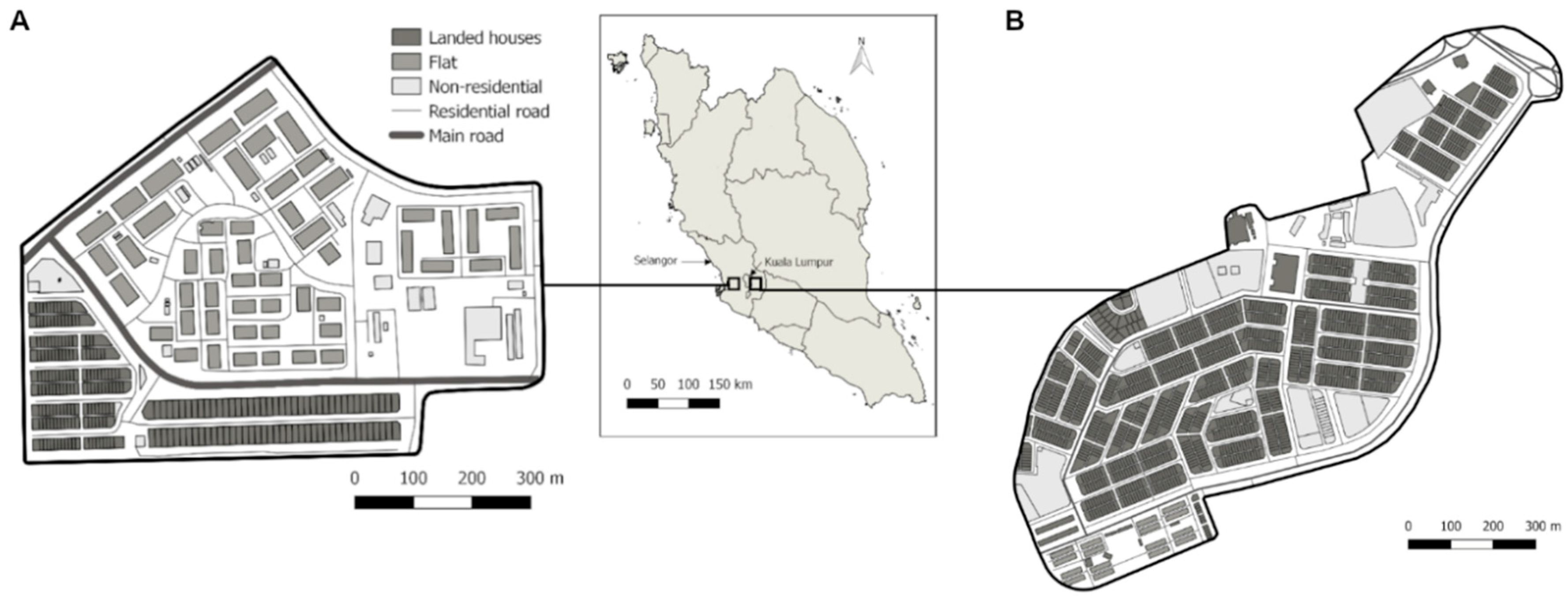
2.1. Sampling and the Wolbachia Release Program
2.2. Colony Samples
2.3. Colony Rearing Protocols
2.4. Backcrossing
2.5. Adult Bioassays
2.6. Assays for Sodium Channel Mutations Associated with Pyrethroid Resistance
2.6.1. Aedes aegypti
2.6.2. Aedes albopictus
2.7. Analyses
3. Results
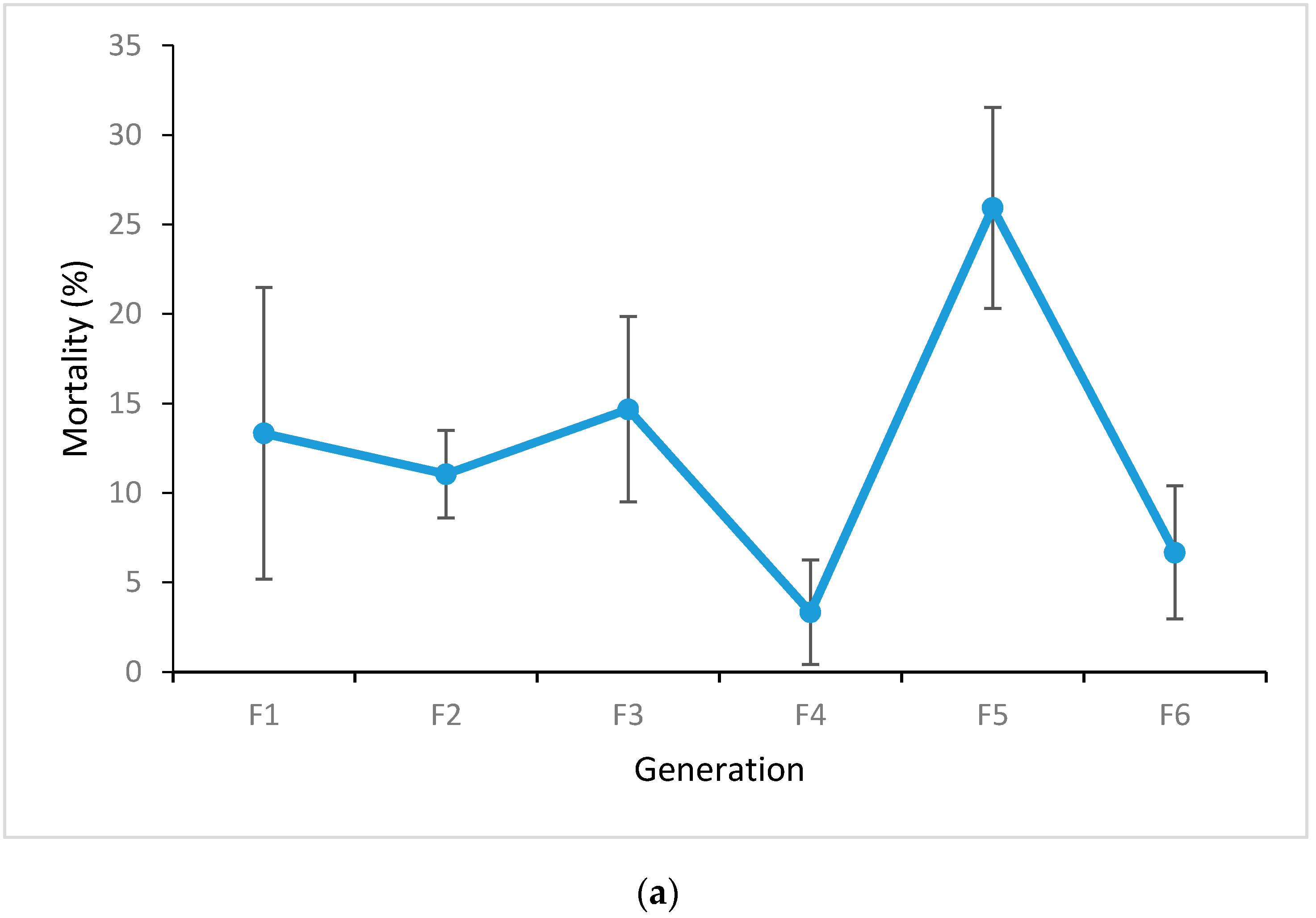
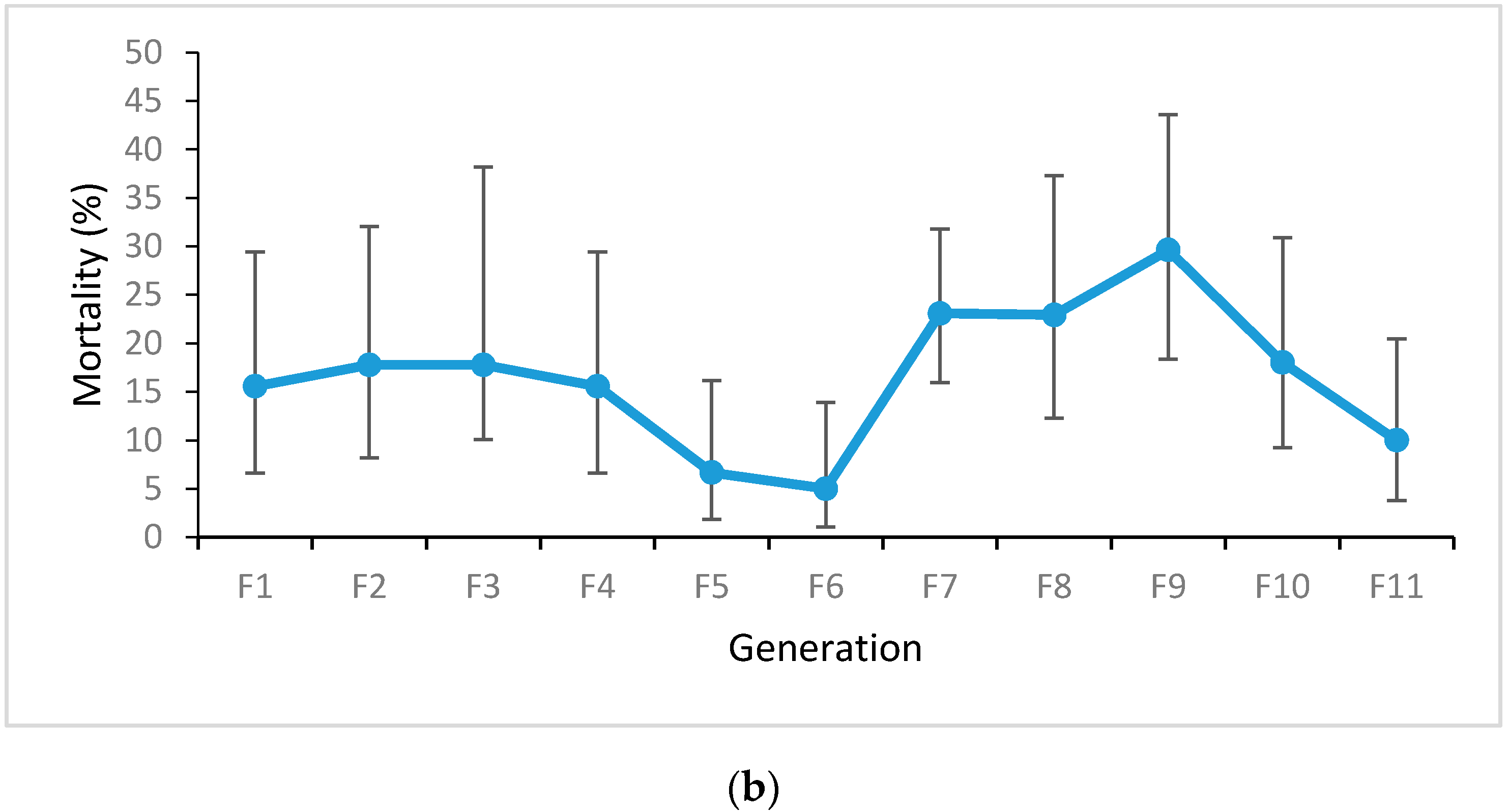
3.1. Adult Bioassays
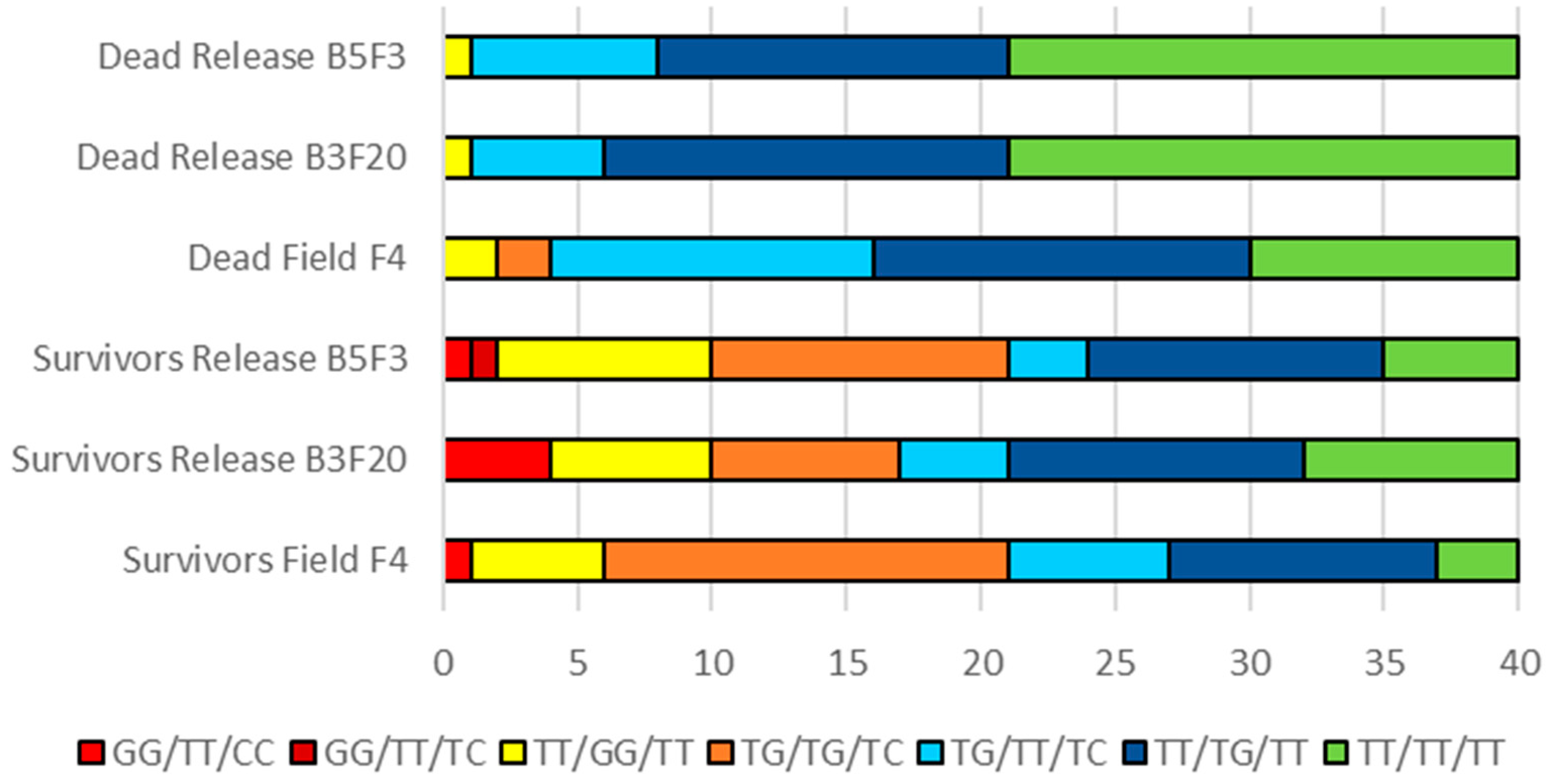
3.2. Assays for Sodium Channel Mutations Associated with Pyrethroid Resistance
3.3. Aedes albopictus
4. Discussion
4.1. Resistance in the Dengue Vector Mosquitoes in Kuala Lumpur
4.2. Comparison between Vssc Genotypes in the Release Colony and the Field
4.3. Changes in Genotype Frequencies in the Release Strain over Time: Implications for Field Survival
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ministry of Health Malaysia. 2019. Available online: http://idengue.remotesensing.gov.my/idengue/index.php (accessed on 9 August 2020).
- Loke, S.R.; Ann, A.T.W.; Nazni, W.A.; Lee, H.L.; Sofian Azirun, M. Insecticide susceptibility status of field-collected Aedes (Stegomyia) aegypti (L.) at a dengue endemic site in Shah Alam, Selangor, Malaysia. Southeast Asian J. Trop. Med. Public Health 2012, 43, 34–47. [Google Scholar]
- Wan-Norafikah, O.; Nazni, W.A.; Lee, H.; Zainol-Ariffin, P.; Sofian-Azirun, M. Permethrin resistance in Aedes aegypti (Linnaeus) collected from Kuala Lumpur, Malaysia. J. Asia Pac. Entomol. 2010, 13, 175–182. [Google Scholar] [CrossRef]
- Ishak, I.H.; Jaal, Z.; Ranson, H.; Wondji, C.S. Contrasting patterns of insecticide resistance and knockdown resistance (kdr) in the dengue vectors Aedes aegypti and Aedes albopictus from Malaysia. Parasites Vectors 2015, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidayati, H.; Sofian Azirun, M.; Nazni, W.A.; Lee, H.L. Insecticide resistance development in Culex quinquefasciatus (Say), Aedes aegypti (L.) and Aedes albopictus (Skuse) larvae against malathion, permethrin and temephos. Trop. Biomed. 2005, 22, 45–52. [Google Scholar]
- WHO. Chemistry and Specifications on Insecticides: Sixteenth Report of the WHO Expert Committee on Vector Biology and Control; WHO Technical Report Series 899; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- Chen, C.D.; Nazni, W.A.; Lee, H.L.; Sofian Azirun, M. Susceptibility of Aedes aegypti and Aedes albopictus to temephos in four study sites in Kuala Lumpur City Center and Selangor State, Malaysia. Trop. Biomed. 2005, 22, 207–216. [Google Scholar] [PubMed]
- Nazni, W.A.; Lee, H.L.; Azahari, A.H. Adult and larval insecticide susceptibility status of Culex quinquefasciatus (Say) mosquitoes in Kuala Lumpur. Trop. Biomed. 2005, 22, 63–68. [Google Scholar]
- Rohani, A.; Ismail, S.; Malinda, M.; Anuar, I.; Mohd Mazlan, I.; Salmah Maszaitun, M. Aedes larval population dynamics and risk for dengue epidemics in Malaysia. Trop. Biomed. 2011, 28, 237–248. [Google Scholar]
- El-Garj, F.M.A.; Wajidi, M.F.F.; Avicor, S.W. Allelic variants of cytochrome P450 monooxygenases: Constitutive and insecticide-mediated expression in a Malaysian strain of the dengue vector, Aedes aegypti (Diptera: Culicidae). Eur. J. Entomol. 2016, 113, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Ishak, I.H.; Kamgang, B.; Ibrahim, S.S.; Riveron, J.M.; Irving, H.; Wondji, C.S. Pyrethroid resistance in Malaysian populations of dengue vector Aedes aegypti is mediated by CYP9 family of cytochrome P450 genes. PLoS Negl. Trop. Dis. 2017, 11, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ishak, I.H.; Riveron, J.M.; Ibrahim, S.S.; Stott, R.; Longbottom, J.; Irving, H.; Wondji, C.S. The Cytochrome P450 gene CYP6P12 confers pyrethroid resistance in kdr-free Malaysian populations of the dengue vector Aedes albopictus. Sci. Rep. 2016, 6, 24707. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Nomura, Y.; Satar, G.; Hu, Z.; Nauen, R.; He, S.Y.; Zhorov, B.S.; Dong, K. Molecular evidence for dual pyrethroid-receptor sites on a mosquito sodium channel. Proc. Natl. Acad. Sci. USA 2013, 110, 11785–11790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuliandari, J.; Lee, S.; White, V.; Tantowijoyo, W.; Hoffmann, A.; Endersby-Harshman, N. Association between three mutations, F1565C, V1023G and S996P, in the voltage-sensitive sodium channel gene and knockdown resistance in Aedes aegypti from Yogyakarta, Indonesia. Insects 2015, 6, 658–685. [Google Scholar] [CrossRef] [PubMed]
- Sayono, S.; Hidayati, A.P.N.; Fahri, S.; Sumanto, D.; Dharmana, E.; Hadisaputro, S.; Asih, P.B.S.; Syafruddin, D. Distribution of Voltage-Gated Sodium Channel (Nav) alleles among the Aedes aegypti populations in Central Java Province and its association with resistance to pyrethroid insecticides. PLoS ONE 2016, 11, e0150577. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Nomura, Y.; Zhorov, B.S.; Dong, K. Sodium channel mutations and pyrethroid resistance in Aedes aegypti. Insects 2016, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Somwang, P.; Prapanthadara, L.-A. High-throughput assays for detection of the F1534C mutation in the voltage-gated sodium channel gene in permethrin-resistant Aedes aegypti and the distribution of this mutation throughout Thailand. Trop. Med. Int. Health TM IH 2011, 16, 501–509. [Google Scholar] [CrossRef]
- Kawada, H.; Higa, Y.; Komagata, O.; Kasai, S.; Tomita, T.; Nguyen Thi, Y.; Luu Lee, L.; Sánchez, R.A.P.; Takagi, M. Widespread distribution of a newly found point mutation in Voltage-Gated Sodium Channel in pyrethroid-resistant Aedes aegypti populations in Vietnam. PLoS Negl. Trop. Dis. 2009, 3, e527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, A.F.; Rajatileka, S.; Ranson, H. Pyrethroid resistance in Aedes aegypti from Grand Cayman. Am. J. Trop. Med. Hyg. 2010, 83, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Linss, B.; Gerlinde, J.; Brito, L.P.; Azambuja Garcia, G.; Saori Araki, A.; Vieira Bruno, R.; Pereira Lima, J.B.; Valle, D.; Martins, A.J. Distribution and dissemination of the Val1016Ile and Phe1534Cys kdr mutations in Aedes aegypti Brazilian natural populations. Parasites Vectors 2014, 7, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-X.; Kaufman, P.E.; Xue, R.-D.; Zhao, M.-H.; Wang, G.; Yan, T.; Guo, X.-X.; Zhang, Y.-M.; Dong, Y.-D.; Xing, D.; et al. Relationship between insecticide resistance and kdr mutations in the dengue vector Aedes aegypti in Southern China. Parasites Vectors 2015. [Google Scholar] [CrossRef] [Green Version]
- Endersby-Harshman, N.M.; Schmidt, T.L.; Chung, J.; van Rooyen, A.; Weeks, A.R.; Hoffmann, A.A. Heterogeneous genetic invasions of three insecticide resistance mutations in Indo-Pacific populations of Aedes aegypti (L.). Mol. Ecol. 2020. [Google Scholar] [CrossRef]
- Kawada, H.; Oo, S.Z.M.; Thaung, S.; Kawashima, E.; Maung, Y.N.M.; Thu, H.M.; Thant, K.Z.; Minakawa, N. Co-occurrence of point mutations in the voltage-gated sodium channel of pyrethroid-resistant Aedes aegypti populations in Myanmar. PLoS Negl. Trop Dis. 2014, 8, e3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Nazawi, A.M.; Aqili, J.; Alzahrani, M.; McCall, P.J.; Weetman, D. Combined target site (kdr) mutations play a primary role in highly pyrethroid resistant phenotypes of Aedes aegypti from Saudi Arabia. Parasites Vectors 2017, 10, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazni, W.A.; Hoffmann, A.A.; NoorAfizah, A.; Cheong, Y.L.; Mancini, M.V.; Golding, N.; Kamarul, G.M.R.; Arif, M.A.K.; Thohir, H.; NurSyamimi, H.; et al. Establishment of Wolbachia strain wAlbB in Malaysian populations of Aedes aegypti for dengue control. Curr. Biol. 2019, 29, 4241–4248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ant, T.H.; Herd, C.S.; Geoghegan, V.; Hoffmann, A.A.; Sinkins, S.P. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathog. 2018, 14, e1006815. [Google Scholar] [CrossRef] [PubMed]
- Yeap, H.L.; Mee, P.; Walker, T.; Weeks, A.R.; O’Neill, S.L.; Johnson, P.; Ritchie, S.A.; Richardson, K.M.; Doig, C.; Endersby, N.M.; et al. Dynamics of the “popcorn” Wolbachia infection in outbred Aedes aegypti informs prospects for mosquito vector control. Genetics 2011, 187, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbe-Ormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. Available online: http://www.nature.com/nature/journal/v476/n7361/abs/nature10356.html#supplementary-information (accessed on 9 August 2020). [CrossRef]
- Endersby, N.M.; Hoffmann, A.A. Effect of Wolbachia on insecticide susceptibility in lines of Aedes aegypti. Bull. Entomol. Res. 2013, 103, 269–277. [Google Scholar] [CrossRef]
- Garcia, G.A.; Hoffmann, A.A.; Maciel-de-Freitas, R.; Villela, D.A.M. Aedes aegypti insecticide resistance underlies the success (and failure) of Wolbachia population replacement. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Garcia, G.d.A.; Sylvestre, G.; Aguiar, R.; da Costa, G.B.; Martins, A.J.; Lima, J.B.P.; Petersen, M.T.; Lourenço-de-Oliveira, R.; Shadbolt, M.F.; Rašić, G.; et al. Matching the genetics of released and local Aedes aegypti populations is critical to assure Wolbachia invasion. PLoS Negl. Trop. Dis. 2019, 13, e0007023. [Google Scholar] [CrossRef]
- Tee, A.S.; Daud, A.R.; Alias, M.; Lee, H.L.; Tham, A.S. Guidelines on the Use of Ovitraps for Aedes Surveillance. Kuala Lumpur: Vector Control. Unit Vector Borne Disease Section; Ministry of Health Malaysia: Putrajaya, Malaysia, 1997.
- Noor Afizah, A.; Mahirah, M.; Azahari, A.; Khairul Asuad, M.; Nazni, W.; Lee, H. Absence of Aedes aegypti (L.) on an ecological island: Competitive Exclusion? Southeast Asian J. Trop. Med. Public Health 2015, 46, 850–856. [Google Scholar]
- WHO. Instructions for Determining the Susceptibility or Resistance of Mosquito Larvae to Insecticides; World Health Organization: Geneva, Switzerland, 1981; Available online: http://www.who.int/iris/handle/10665/69615 (accessed on 9 August 2020).
- WHO. Monitoring and Managing Insecticide Resistance in Aedes Mosquito Populations: Interim Guidance for Entomologists; World Health Organization: Geneva, Switzerland, 2016; Available online: http://www.who.int/iris/handle/10665/204588 (accessed on 9 August 2020).
- Kasai, S.; Komagata, O.; Itokawa, K.; Shono, T.; Ng, L.C.; Kobayashi, M.; Tomita, T. Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti: Target site insensitivity, penetration, and metabolism. PLoS Negl. Trop. Dis. 2014, 8, e2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, S.; Ng, L.C.; Lam-Phua, S.G.; Tang, C.S.; Itokawa, K.; Komagata, O.; Kobayashi, M.; Tomita, T. First detection of a putative knockdown resistance gene in major mosquito vector, Aedes albopictus. Jpn. J. Infect. Dis. 2011, 64, 217–221. [Google Scholar] [PubMed]
- Endersby-Harshman, N.M.; Wuliandari, J.R.; Harshman, L.G.; Frohn, V.; Johnson, B.J.; Ritchie, S.A.; Hoffmann, A.A. Pyrethroid susceptibility has been maintained in the dengue vector, Aedes aegypti (Diptera: Culicidae), in Queensland, Australia. J. Med. Entomol. 2017, 54, 1649–1658. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. Available online: http://www.nature.com/nature/journal/v476/n7361/abs/nature10355.html#supplementary-information (accessed on 9 August 2020). [CrossRef] [PubMed]
- Smith, L.B.; Kasai, S.; Scott, J.G. Pyrethroid resistance in Aedes aegypti and Aedes albopictus: Important mosquito vectors of human diseases. Pestic. Biochem. Physiol. 2016, 133, 1–12. [Google Scholar] [CrossRef]
- Su, X.; Guo, Y.; Deng, J.; Xu, J.; Zhou, G.; Zhou, T.; Li, Y.; Zhong, D.; Kong, L.; Wang, X.; et al. Fast emerging insecticide resistance in Aedes albopictus in Guangzhou, China: Alarm to the dengue epidemic. PLoS Negl. Trop. Dis. 2019, 13, e0007665. [Google Scholar] [CrossRef]
- Zhou, X.; Li, M.; Liu, N.; Qiu, X.; Tong, Y.; Yang, C.; Zeng, X. Knockdown resistance (kdr) mutations within seventeen field populations of Aedes albopictus from Beijing China: First report of a novel V1016G mutation and evolutionary origins of kdr haplotypes. Parasites Vectors 2019, 12, 180. [Google Scholar] [CrossRef]
- Marcombe, S.; Farajollahi, A.; Healy, S.P.; Clark, G.G.; Fonseca, D.M. Insecticide resistance status of United States populations of Aedes albopictus and mechanisms involved. PLoS ONE 2014, 9, e101992. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Bonizzoni, M.; Zhong, D.; Zhou, G.; Cai, S.; Li, Y.; Wang, X.; Lo, E.; Lee, R.; Sheen, R.; et al. Multi-country survey revealed prevalent and novel F1534S mutation in Voltage-Gated Sodium Channel (VGSC) gene in Aedes albopictus. PLoS Negl. Trop. Dis. 2016, 10, e0004696. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.; Zhou, Q.; Xu, Z.; Zhu, G.; Dong, K.; Zhorov, B.S.; Chen, M. Three sodium channel mutations from Aedes albopictus confer resistance to Type I, but not Type II pyrethroids. Insect Biochem. Mol. Biol. 2020. [Google Scholar] [CrossRef]
- Li, Y.; Xu, J.; Zhong, D.; Zhang, H.; Yang, W.; Zhou, G.; Su, X.; Wu, Y.; Wu, K.; Cai, S.; et al. Evidence for multiple-insecticide resistance in urban Aedes albopictus populations in southern China. Parasites Vectors 2018, 11, 4. [Google Scholar] [CrossRef] [PubMed]
- Rasli, R.; Lee, H.L.; Wasi Ahmad, N.; Fikri, S.F.F.; Ali, R.; Muhamed, K.A.; Hadi, A.A.; Liu, Q.Y.; Meng, F.X. Susceptibility status and resistance mechanisms in permethrin-selected, laboratory susceptible and field-collected Aedes aegypti from Malaysia. Insects 2018, 9, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, N.; Kwiatkowska, R.M.; Irving, H.; Diabaté, A.; Dabire, R.; Wondji, C.S. Target-site resistance mutations (kdr and RDL), but not metabolic resistance, negatively impact male mating competiveness in the malaria vector Anopheles gambiae. Heredity 2015, 115, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype Frequency (%) (n) | ||||||||
|---|---|---|---|---|---|---|---|---|
| GG/TT/CC | TT/GG/TT | TG/TG/TC | TG/TT/TC | TT/TG/TT | TT/TT/TT | TG/TG/TT | ||
| Gen | A | B | C | D | E | F | H | |
| Survivors | Field 2016 | 2.5 (1) | 30.0 (12) | 67.5 (27) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| Dead | Field 2016 | 17.5 (7) | 32.5 (13) | 50.0 (20) | 0.0 (0) | 0.0 (0) | 0.0 (0) | 0.0 (0) |
| Survivors | Field 2019 | 15.0 (6) | 35.0 (14) | 45.0 (18) | 2.5 (1) | 2.5 (1) | 0.0 (0) | 0.0 (0) |
| Dead | Field 2019 | 12.5 (5) | 37.5 (15) | 35.0 (14) | 0.0 (0) | 12.5 (5) | 2.5 (1) | 0.0 (0) |
| Survivors | Release B3F9 | 2.5 (1) | 15.0 (6) | 15.0 (6) | 5.0 (2) | 52.5 (21) | 10.0 (4) | 0.0 (0) |
| Dead | Release B3F9 | 2.5 (1) | 10.0 (4) | 10.0 (4) | 10.0 (4) | 35.0 (14) | 32.5 (13) | 0.0 (0) |
| Survivors | Release B5F0 | 2.5 (1) | 42.5 (17) | 27.5 (11) | 5.0 (2) | 22.5 (9) | 0.0 (0) | 0.0 (0) |
| Dead | Release B5F0 | 5.0 (2) | 35.0 (14) | 20.0 (8) | 0.0 (0) | 35.0 (14) | 0.0 (0) | 5.0 (2) |
| Odds of Surviving | n | OR | 95% Confidence Interval | OR Significant α = 0.05 | ||
|---|---|---|---|---|---|---|
| If: | Rather Than: | Lower | Upper | |||
| TG/TG/TC | TT/TT/TT | 108, 18 | 4.72 | 1.46 | 15.27 | * |
| TG/TT/TC | TT/TT/TT | 9, 18 | 4.38 | 0.78 | 24.47 | |
| TT/GG/TT | TT/TT/TT | 95, 18 | 3.73 | 1.14 | 12.16 | * |
| TT/TG/TT | TT/TT/TT | 64, 18 | 3.29 | 0.98 | 11.08 | |
| GG/TT/CC | TT/TT/TT | 24, 18 | 2.10 | 0.53 | 8.39 | |
| TG/TG/TC | GG/TT/CC | 108, 24 | 2.25 | 0.90 | 5.58 | |
| TG/TT/TC | GG/TT/CC | 9, 24 | 2.08 | 0.44 | 9.84 | |
| TT/GG/TT | GG/TT/CC | 95, 24 | 1.78 | 0.71 | 4.45 | |
| TT/TG/TT | GG/TT/CC | 64, 24 | 1.57 | 0.60 | 12.09 | |
| TG/TG/TC | TT/GG/TT | 108, 95 | 1.27 | 0.73 | 2.20 | |
| TT/GG/TT | TT/TG/TT | 95, 64 | 1.13 | 0.60 | 2.14 | |
| TG/TG/TC | TG/TT/TC | 108, 9 | 1.08 | 0.27 | 4.24 | |
| (a) | Genotype Frequency (%) (n) | |||||||
| GG/TT/CC | TT/GG/TT | TG/TG/TC | TG/TT/TC | TT/TG/TT | TT/TT/TT | GG/TT/TC | ||
| Generation | A | B | C | D | E | F | G | |
| Survivors | Field F4 | 2.5 (1) | 12.5 (5) | 37.5 (15) | 15.0 (6) | 25.0 (10) | 7.5 (3) | 0.0 (0) |
| Dead | 0.0 (0) | 5.0 (2) | 5.0 (2) | 30.0 (12) | 35.0 (14) | 25.0 (10) | 0.0 (0) | |
| Survivors | Release B3F20 | 10.0 (4) | 15.0 (6) | 17.5 (7) | 10.0 (4) | 27.5 (11) | 20.0 (8) | 0.0 (0) |
| Dead | 0.0 (0) | 2.5 (1) | 0.0 (0) | 12.5 (5) | 37.5 (15) | 47.5 (19) | 0.0 (0) | |
| Survivors | Release B5F3 | 2.5 (1) | 20.0 (8) | 27.5 (11) | 7.5 (3) | 27.5 (11) | 12.5 (5) | 2.5 (1) |
| Dead | 0.0 (0) | 2.5 (1) | 0.0 (0) | 17.5 (7) | 32.5 (13) | 47.5 (19) | 0.0 (0) | |
| (b) | Odds of Surviving | n | OR | 95% Confidence Interval | OR Significant α = 0.05 | |||
| if: | rather than: | Lower | Upper | |||||
| TT/GG/TT | TG/TG/TC | 23, 35 | 0.29 | 0.05 | 1.72 | |||
| TT/GG/TT | TG/TT/TC | 23, 37 | 8.77 | 2.46 | 31.29 | * | ||
| TT/GG/TT | TT/TG/TT | 23, 74 | 6.23 | 1.93 | 20.13 | * | ||
| TT/GG/TT | TT/TT/TT | 23, 64 | 14.25 | 4.22 | 48.16 | * | ||
| TG/TG/TC | TG/TT/TC | 35, 37 | 30.46 | 6.28 | 147.72 | * | ||
| TG/TG/TC | TT/TG/TT | 35, 74 | 21.66 | 4.83 | 97.02 | * | ||
| TG/TG/TC | TT/TT/TT | 35, 64 | 49.50 | 10.66 | 229.83 | * | ||
| TG/TT/TC | TT/TG/TT | 37, 74 | 0.71 | 0.31 | 1.61 | |||
| TG/TT/TC | TT/TT/TT | 37, 64 | 1.63 | 0.67 | 3.92 | |||
| TT/TG//TT | TT/TT/TT | 74, 64 | 2.29 | 1.10 | 4.74 | * | ||
| Genotype Frequency (%) (n) | ||||||
|---|---|---|---|---|---|---|
| GG/TT/CC | TT/GG/TT | TG/TG/TC | TG/TT/TC | TT/TG/TT | n | |
| A * | B * | C * | D | E | ||
| Field Female | 15.00 (6) | 20.00 (8) | 60.00 (24) | 0.0 (0) | 5.00 (2) | 40 |
| Field Male | 12.50 (5) | 45.00 (18) | 37.50 (15) | 0.0 (0) | 5.00 (2) | 40 |
| Total | 13.75 (11) | 32.50 (26) | 48.75 (39) | 0.0 (0) | 5.00 (4) | 80 |
| Colony Female | 15.00 (6) | 5.00 (2) | 47.50 (19) | 7.50 (3) | 25.00 (10) | 40 |
| Colony Male | 10.00 (4) | 45.00 (18) | 27.50 (11) | 7.50 (3) | 10.00 (4) | 40 |
| Total | 12.50 (10) | 25.00 (20) | 37.50 (30) | 7.50 (6) | 17.50 (14) | 80 |
| Genotype Frequency (%) (n) | |||||||
|---|---|---|---|---|---|---|---|
| Genotype | GG/TT/CC | TT/GG/TT | TG/TG/TC | TG/TT/TC | TT/TG/TT | TT/TT/TT | n |
| A | B | C | D | E | F | ||
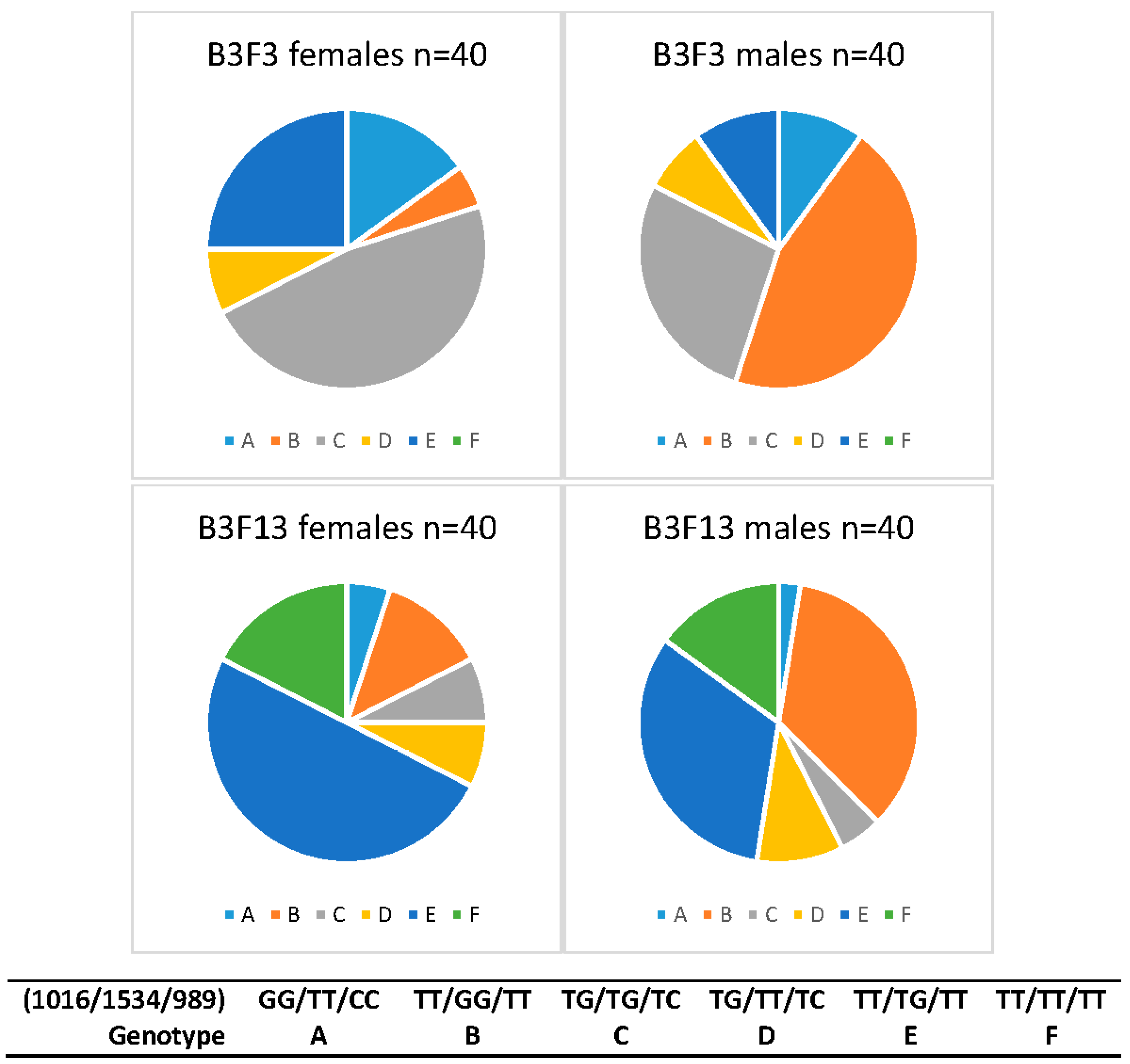
| B3F3 Female | 15.0 (6) | 5.0 (2) | 47.5 (19) | 7.5 (3) | 25.0 (10) | 0.0 (0) | 40 |
| B3F3 Male | 10.0 (4) | 45.0 (18) | 27.5 (11) | 7.5 (3) | 10.0 (4) | 0.0 (0) | 40 |
| B3F8 Female | 0 (0) | 13.3 (4) | 20.0 (6) | 13.3 (4) | 43.3 (13) | 10.0 (3) | 30 |
| B3F8 Male | 7.1 (2) | 46.4 (13) | 3.6 (1) | 7.1 (2) | 21.4 (6) | 14.3 (4) | 28 |
| B3F13 Female | 5.0 (2) | 12.5 (5) | 7.5 (3) | 7.5 (3) | 50.0 (20) | 17.5 (7) | 40 |
| B3F13 Male | 2.5 (1) | 35.0 (14) | 5.0 (2) | 10.0 (4) | 32.5 (13) | 15.0 (6) | 40 |
| B5F0 Female | 7.5 (3) | 25.0 (10) | 37.5 (15) | 7.5 (3) | 22.5 (9) | 0.0 (0) | 40 |
| B5F0 Male | 0.0 (0) | 27.5 (11) | 37.5 (15) | 22.5 (9) | 12.5 (5) | 0.0 (0) | 40 |
| Genotype Frequency (%) (n) | ||||||||
|---|---|---|---|---|---|---|---|---|
| GG/TT/CC | TT/GG/TT | TG/TG/TC | TG/TT/TC | TT/TG/TT | TT/TT/TT | n | ||
| A | B | C | D | E | F | |||
| Field Female | 2016 | 15.00 (6) | 20.00 (8) | 60.00 (24) | 0.0 (0) | 5.00 (2) | 0.00 (0) | 40 |
| Field Male | 12.50 (5) | 45.00 (18) | 37.50 (15) | 0.0 (0) | 5.00 (2) | 0.00 (0) | 40 | |
| Total | 13.75 (11) | 32.50 (26) | 48.75 (39) | 0.0 (0) | 5.00 (4) | 0.00 (0) | 80 | |
| Field Female | 2019 | 15.00 (6) | 32.50 (13) | 35.00 (14) | 2.50 (1) | 12.50 (5) | 2.50 (1) | 40 |
| Field Male | 25.00 (10) | 40.00 (16) | 27.50 (11) | 0.00 (0) | 7.50 (3) | 0.00 (0) | 40 | |
| Total | 20.00 (16) | 36.25 (29) | 31.25 (25) | 1.25 (1) | 10.00 (8) | 1.25 (1) | 80 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, N.A.; Endersby-Harshman, N.M.; Mohd Mazni, N.R.; Mohd Zabari, N.Z.A.; Amran, S.N.S.; Ridhuan Ghazali, M.K.; Abdul Karim, M.A.; Cheong, Y.L.; Sinkins, S.P.; Ahmad, N.W.; et al. Characterization of Sodium Channel Mutations in the Dengue Vector Mosquitoes Aedes aegypti and Aedes albopictus within the Context of Ongoing Wolbachia Releases in Kuala Lumpur, Malaysia. Insects 2020, 11, 529. https://doi.org/10.3390/insects11080529
Ahmad NA, Endersby-Harshman NM, Mohd Mazni NR, Mohd Zabari NZA, Amran SNS, Ridhuan Ghazali MK, Abdul Karim MA, Cheong YL, Sinkins SP, Ahmad NW, et al. Characterization of Sodium Channel Mutations in the Dengue Vector Mosquitoes Aedes aegypti and Aedes albopictus within the Context of Ongoing Wolbachia Releases in Kuala Lumpur, Malaysia. Insects. 2020; 11(8):529. https://doi.org/10.3390/insects11080529
Chicago/Turabian StyleAhmad, Noor Afizah, Nancy M. Endersby-Harshman, Nur Ruqqayah Mohd Mazni, Nur Zatil Aqmar Mohd Zabari, Siti Nor Syazwani Amran, Muhammad Kamarul Ridhuan Ghazali, Mohd Arif Abdul Karim, Yoon Ling Cheong, Steven P. Sinkins, Nazni Wasi Ahmad, and et al. 2020. "Characterization of Sodium Channel Mutations in the Dengue Vector Mosquitoes Aedes aegypti and Aedes albopictus within the Context of Ongoing Wolbachia Releases in Kuala Lumpur, Malaysia" Insects 11, no. 8: 529. https://doi.org/10.3390/insects11080529
APA StyleAhmad, N. A., Endersby-Harshman, N. M., Mohd Mazni, N. R., Mohd Zabari, N. Z. A., Amran, S. N. S., Ridhuan Ghazali, M. K., Abdul Karim, M. A., Cheong, Y. L., Sinkins, S. P., Ahmad, N. W., & Hoffmann, A. A. (2020). Characterization of Sodium Channel Mutations in the Dengue Vector Mosquitoes Aedes aegypti and Aedes albopictus within the Context of Ongoing Wolbachia Releases in Kuala Lumpur, Malaysia. Insects, 11(8), 529. https://doi.org/10.3390/insects11080529