Development of a Mating Disruption Program for a Mealybug, Planococcus ficus, in Vineyards


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Replicated Field Trials—2004 and 2005
2.2. 2006 and 2007 Seasons
2.3. Dispenser Load and Release
2.4. Pheromone Purity
2.5. Statistics
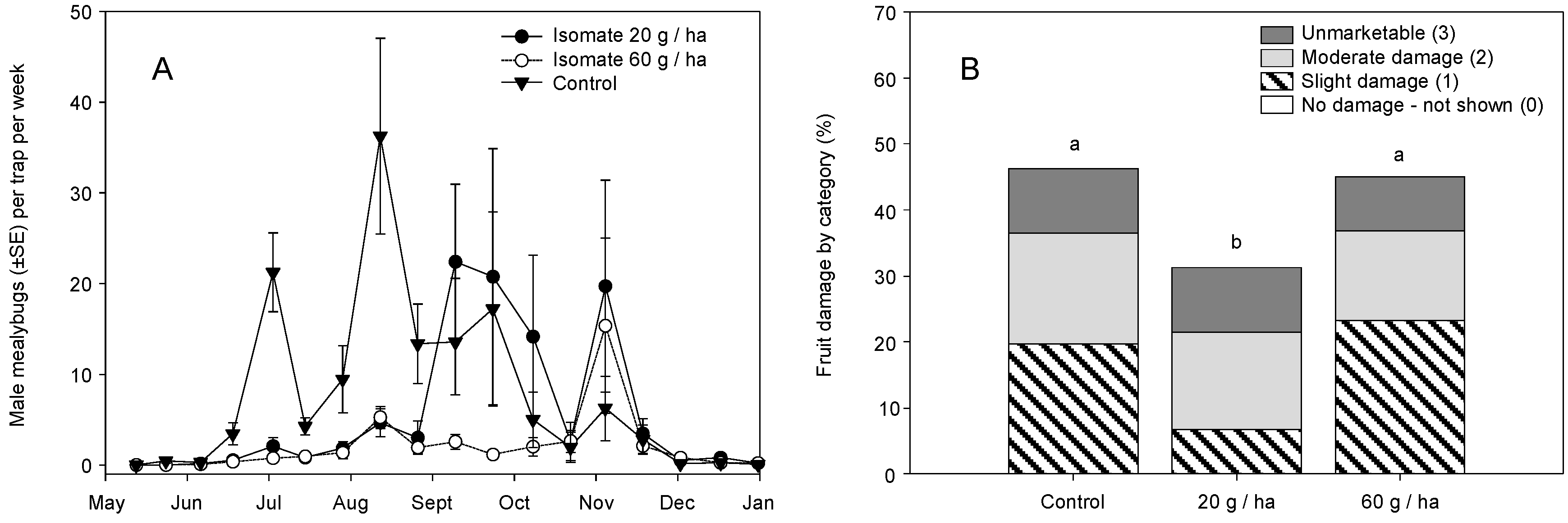
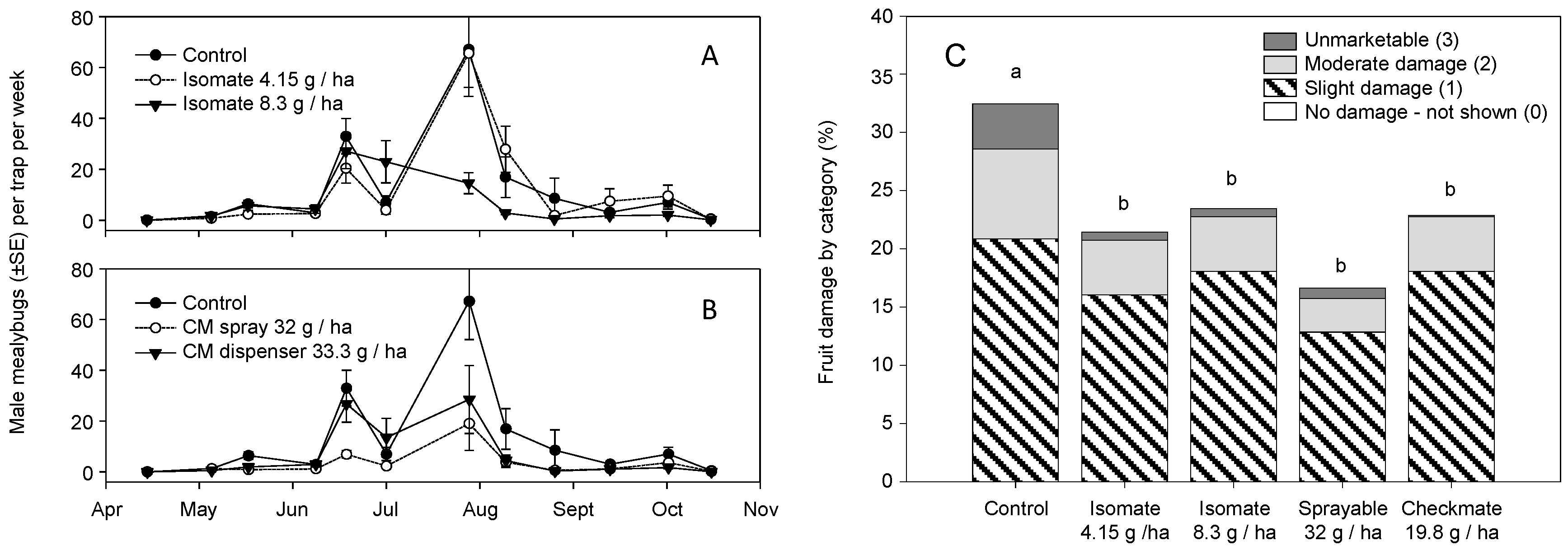
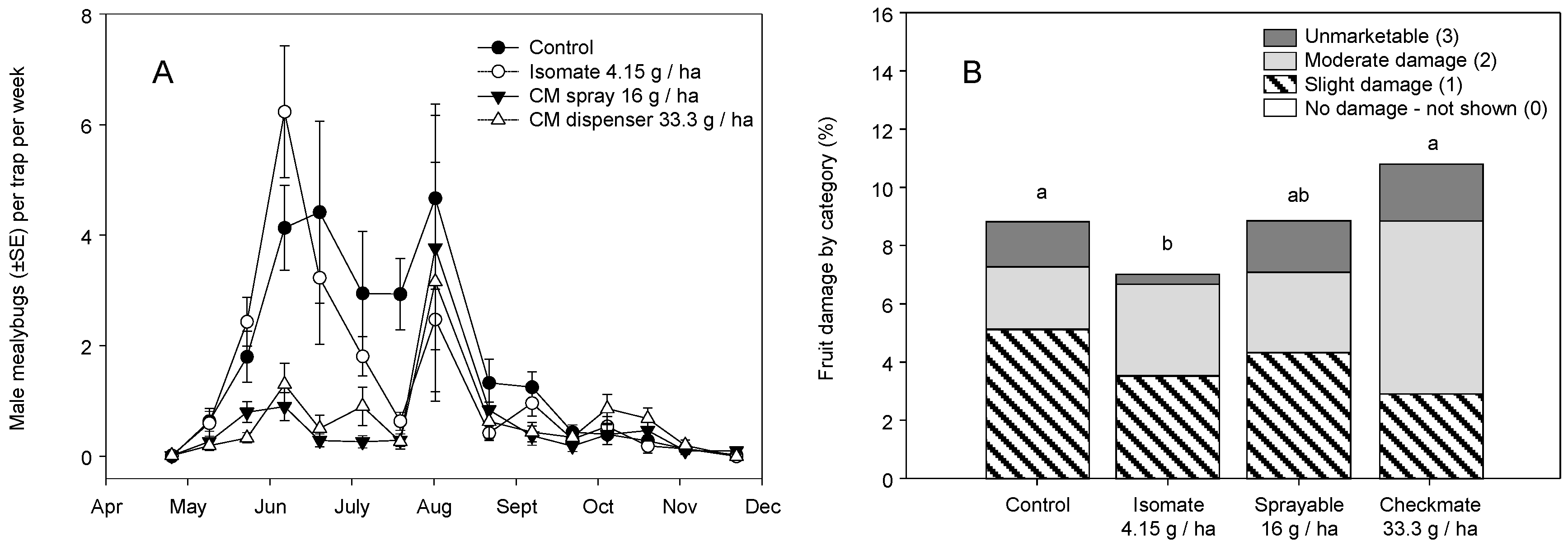
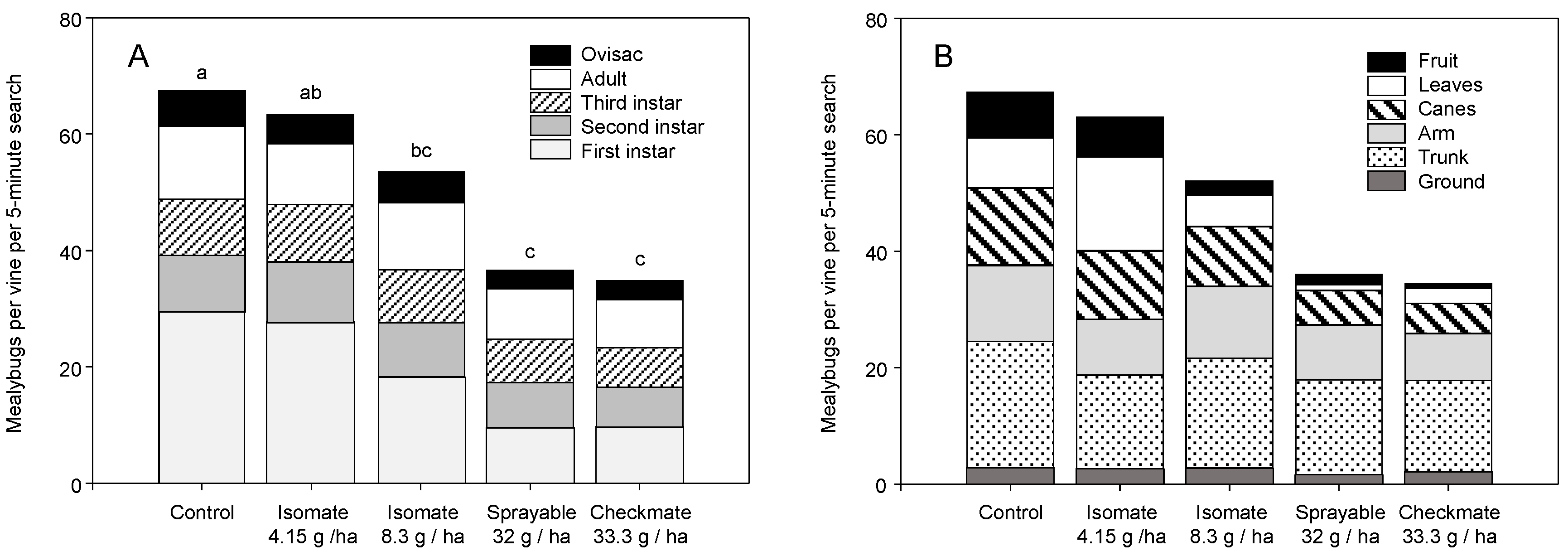
3. Results
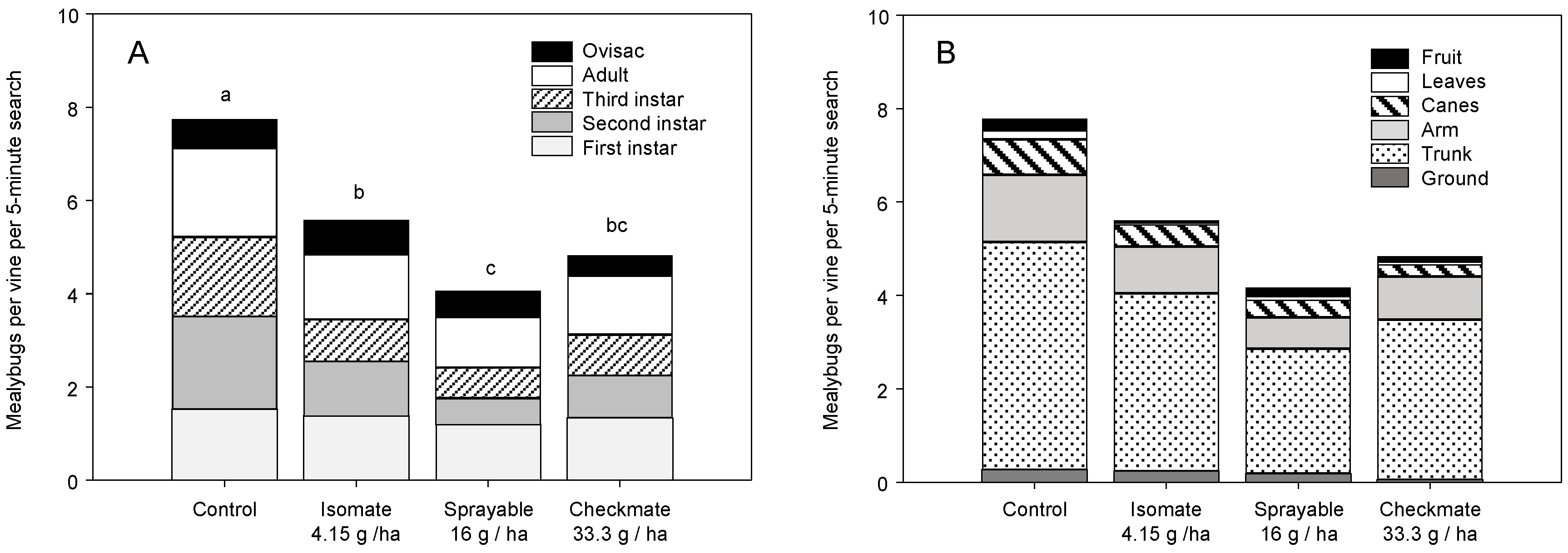
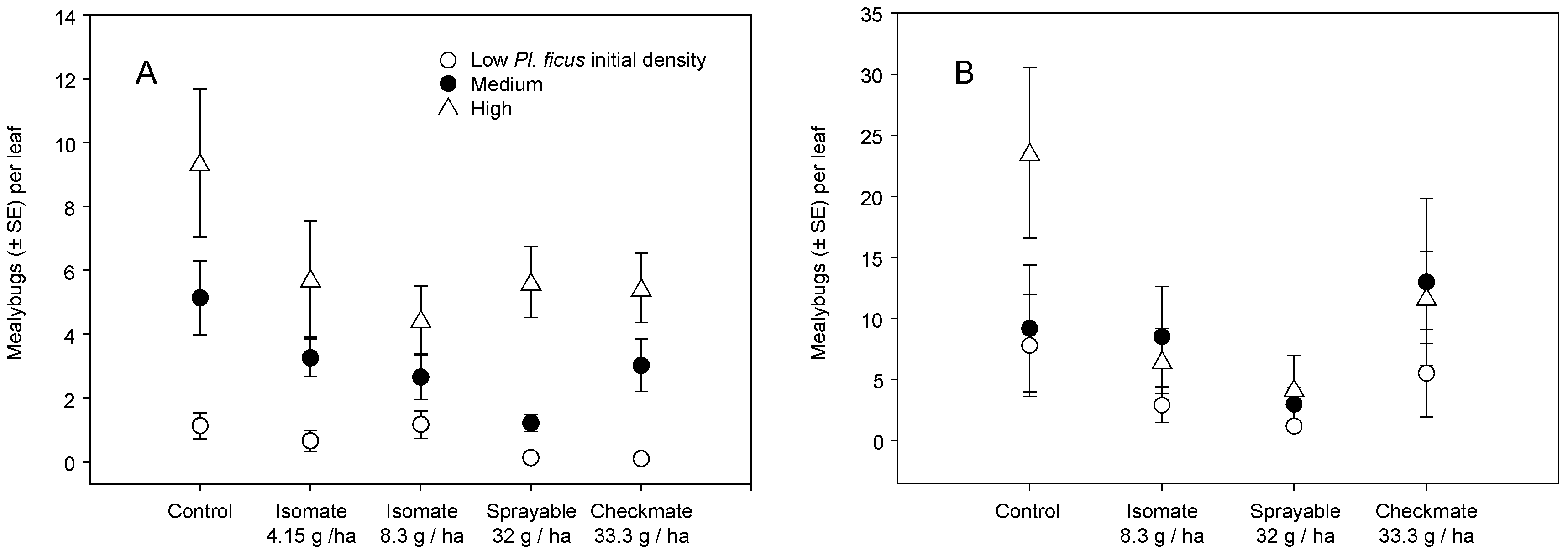
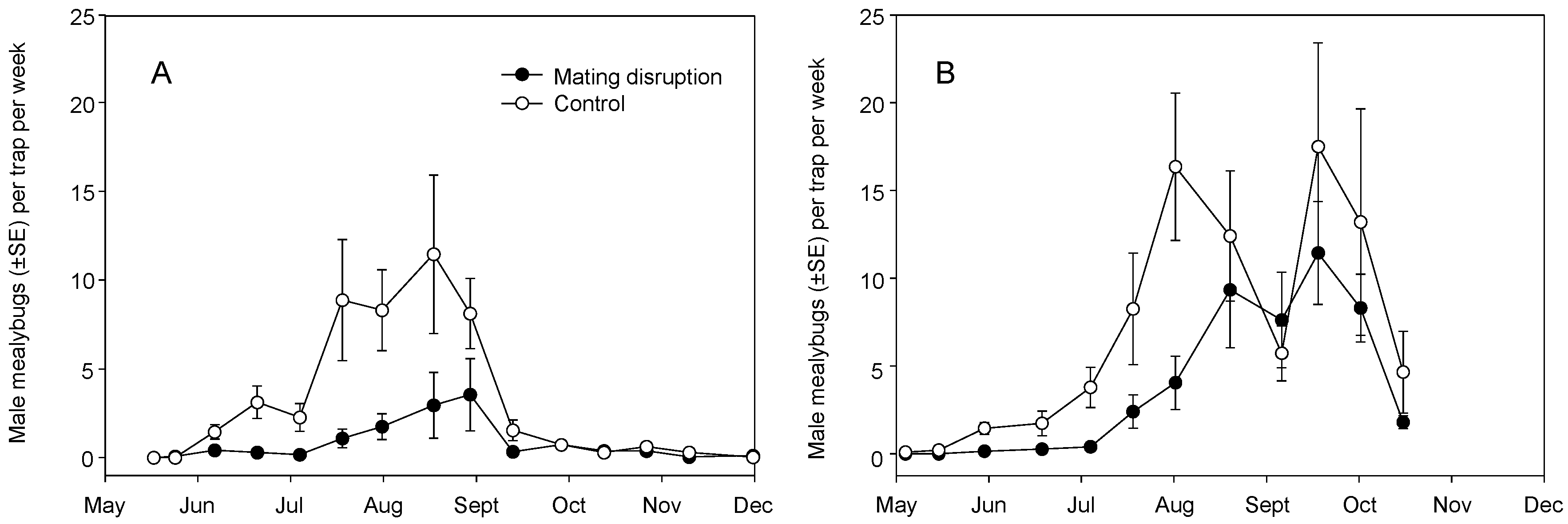
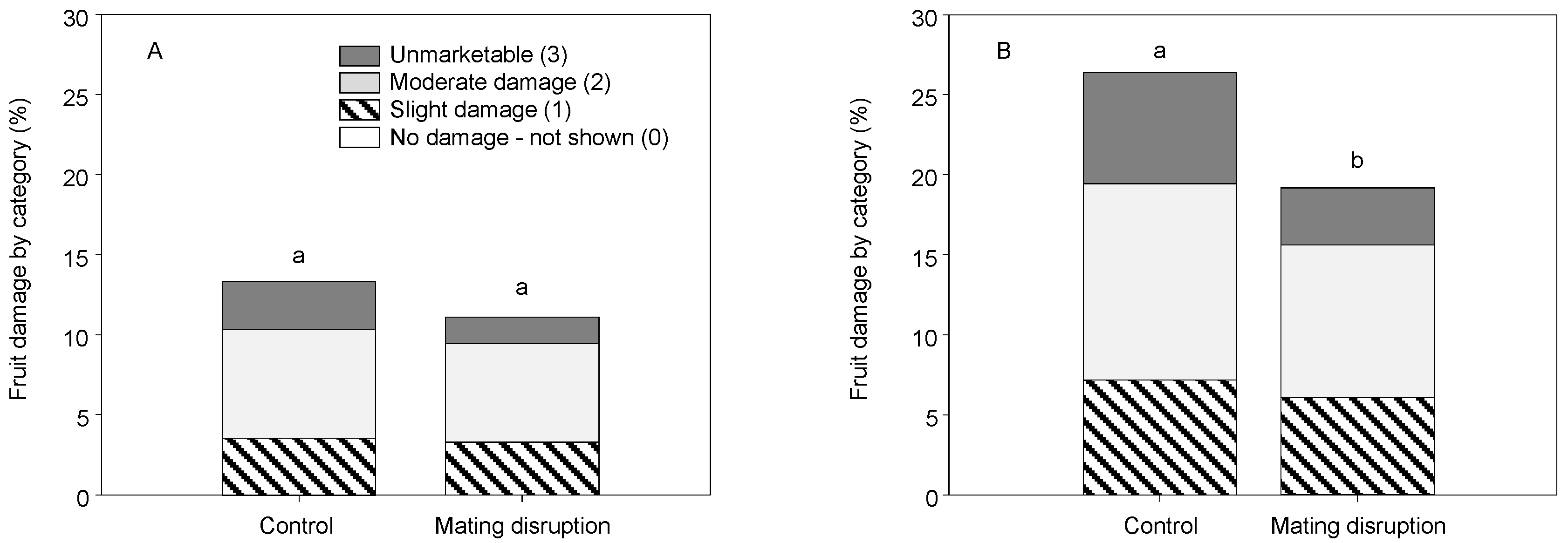
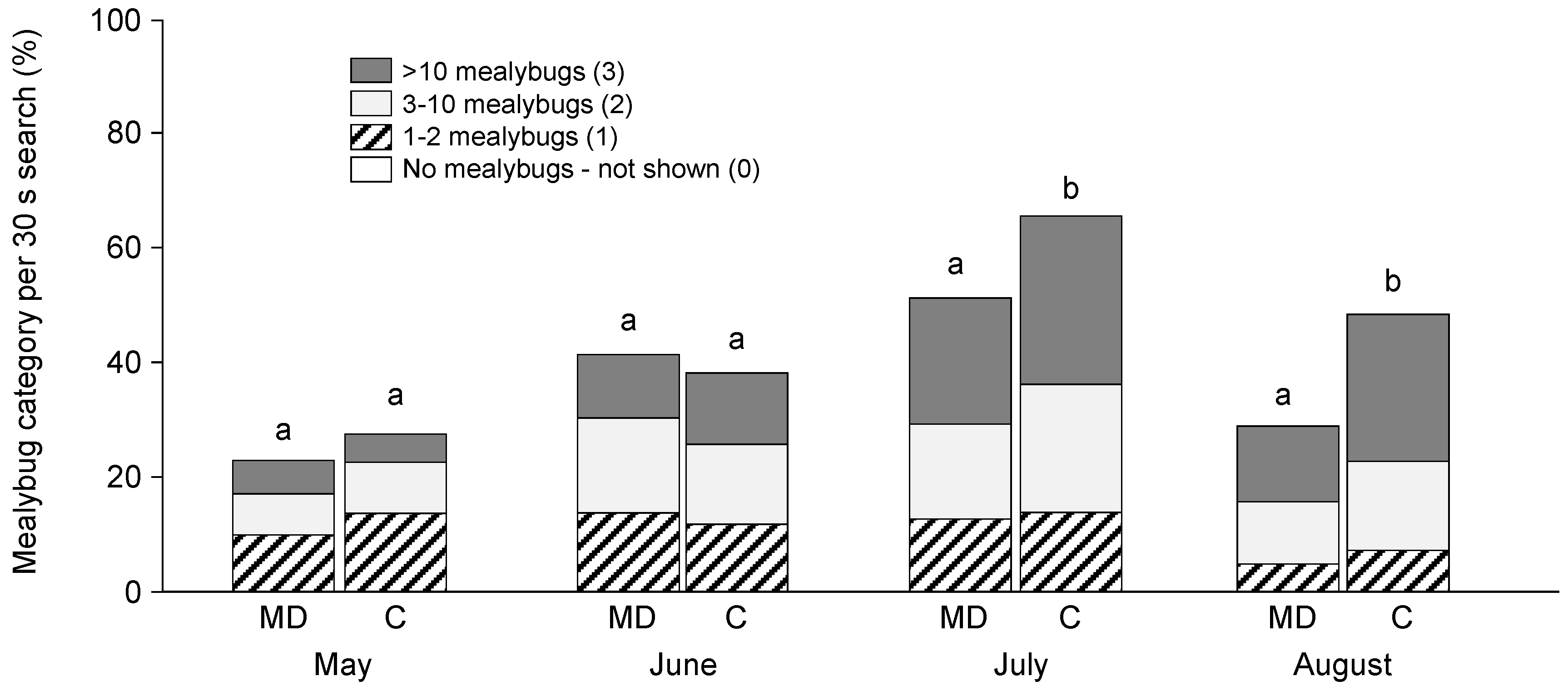
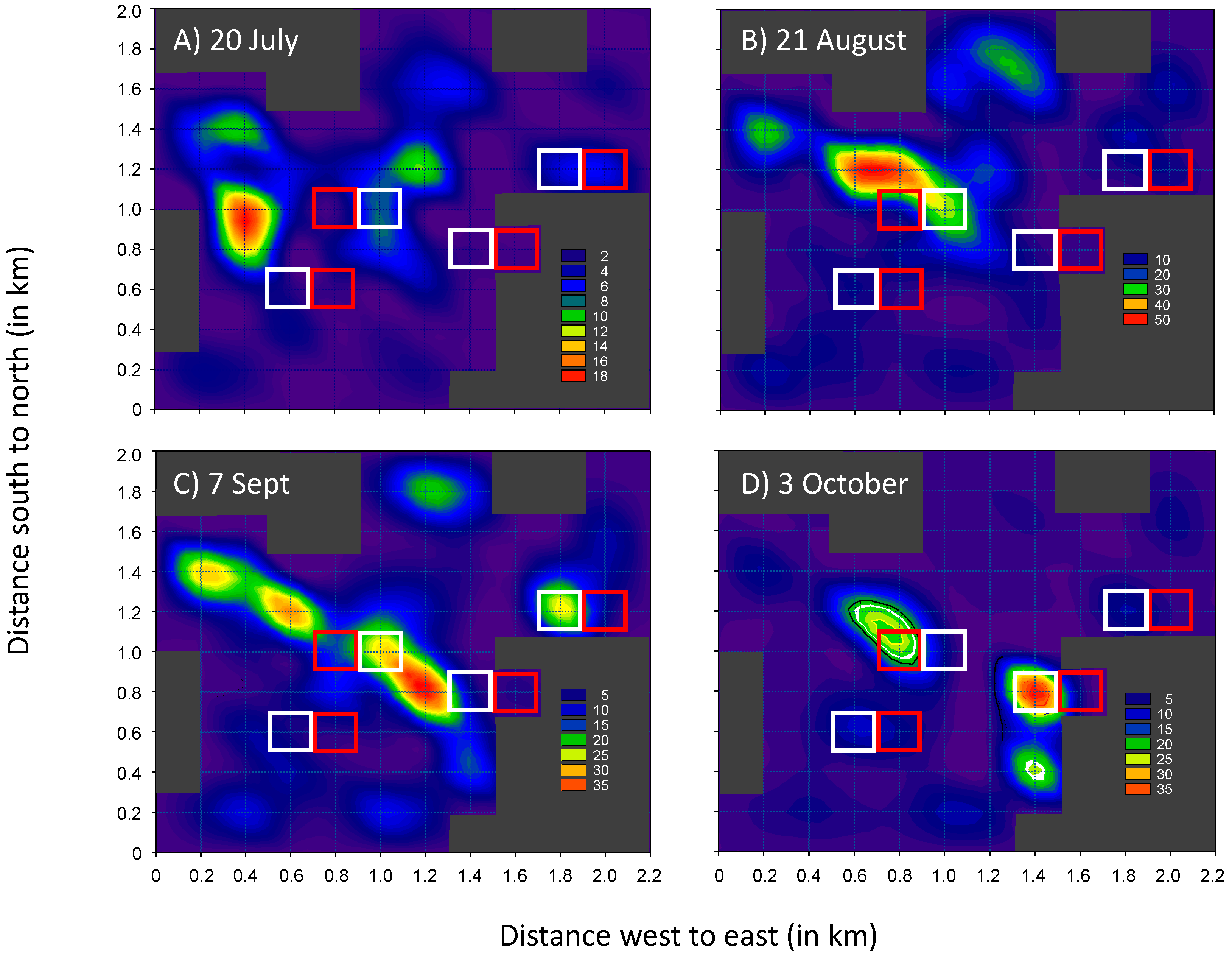
3.1. Field Trials—2004 and 2005
3.2. Field Trials—2006 and 2007
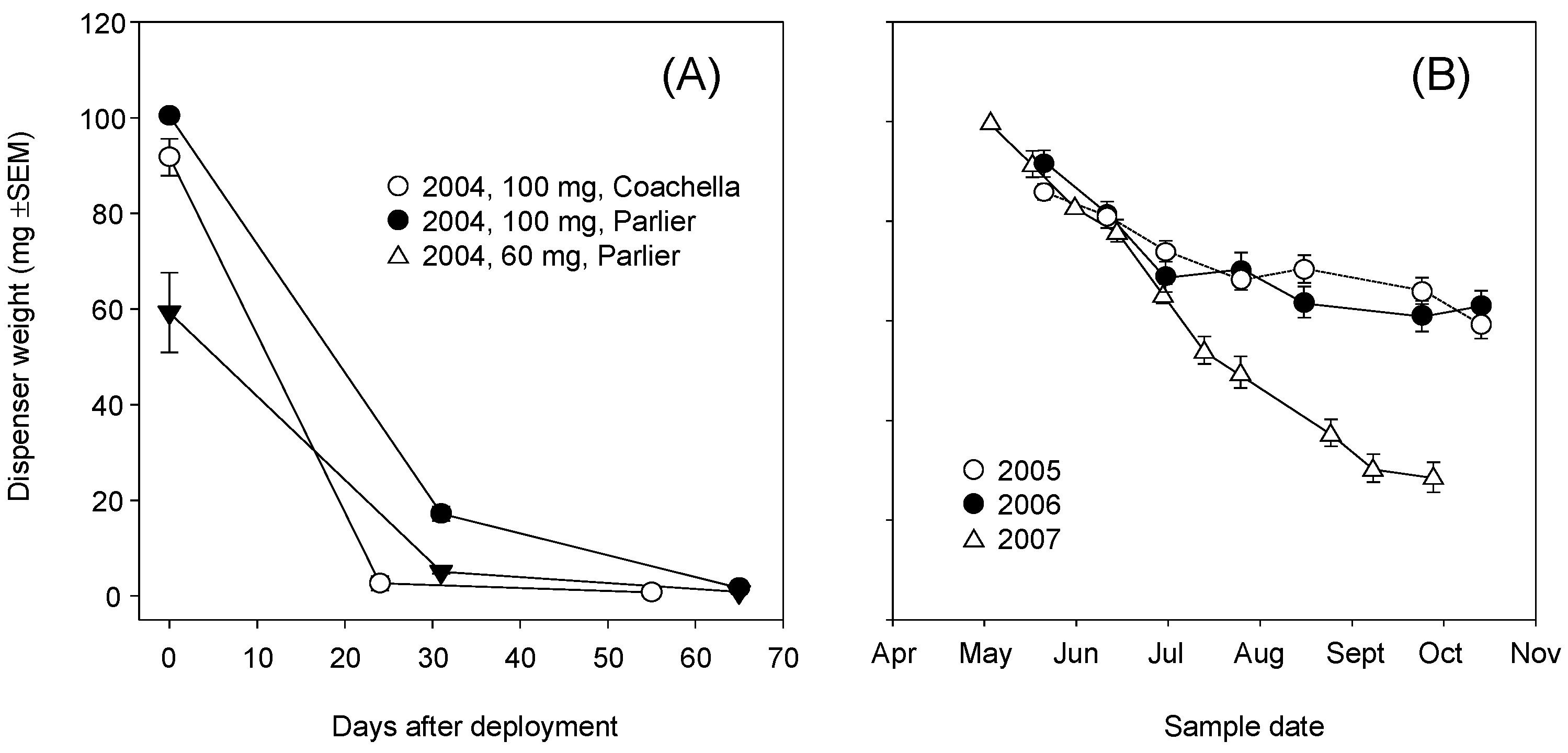
3.3. Dispenser Load and Release Rate
3.4. Pheromone Purity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daane, K.M.; Almeida, R.P.P.; Bell, V.A.; Botton, M.; Fallahzadeh, M.; Mani, M.; Miano, J.L.; Sforza, R.; Walton, V.M.; Zaveizo, T. Biology and management of mealybugs in vineyards. In Arthropod Management in Vineyards; Bostanian, N.J., Isaacs, R., Vincent, C., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 271–308. [Google Scholar] [CrossRef]
- Daane, K.M.; Bentley, W.J.; Smith, R.J.; Haviland, D.R.; Weber, E.; Gispert, C.; Battany, M.C.; Millar, J.G. Planococcus mealybugs (Vine mealybug). In Grape Pest Management, Publication 3343, 3rd ed.; Bettiga, L., Ed.; University of California, Division of Agriculture and Natural Resources: Oakland, CA, USA, 2013; pp. 246–260. [Google Scholar]
- Walton, V.M.; Pringle, K.L. Vine mealybug, Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae), a key pest in South African vineyards. A review. S. Afr. J. Enol. Vitic. 2004, 25, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Bertin, S.; Pacifico, D.; Cavalieri, V.; Marzachi, C.; Bosco, D. Transmission of Grapevine virus A and Grapevine leafroll-associated viruses 1 and 3 by Planococcus ficus and Planococcus citri fed on mixed-infected plants. Ann. Appl. Biol. 2016, 169, 53–63. [Google Scholar] [CrossRef]
- Charles, J.G.; Froud, K.J.; van den Brink, R.; Allan, D.J. Mealybugs and the spread of grapevine leafroll-associated virus 3 (GLRaV-3) in a New Zealand vineyard. Australas. Plant Pathol. 2009, 38, 576–583. [Google Scholar] [CrossRef]
- Almeida, R.P.P.; Daane, K.M.; Bell, V.A.; Blaisdell, G.A.; Cooper, M.L.; Herrbach, E.; Pietersen, G. Ecology and management of grapevine leafroll disease. Front. Microbiol. Virol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.M.; Baraff, B.; Hutchins, J.T.; Wong, M.K.; Blaisdell, G.K.; Cooper, M.L.; Daane, K.M.; Almeida, R.P.P. Relative prevalence of grapevine leafroll-associated virus species in wine grape-growing regions of California. PLoS ONE 2015, 10, e0142120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.M.; Wang, J.B.; Duffy, S.; Zhang, S.M.; Wong, M.K.; Rashed, A.; Cooper, M.L.; Daane, K.M.; Almeida, R.P.P. Occurrence of grapevine leafroll-associated virus complex in Napa Valley. PLoS ONE 2011, 6, e026227. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Lara, A.; Klaassen, V.; Stevens, K.; Sudarshana, M.R.; Rowhani, A.; Maree, H.J.; Chooi, K.M.; Blouin, A.G.; Habili, N.; Song, Y.S.; et al. Characterization of grapevine leafroll-associated virus 3 genetic variants and application towards RT-qPCR assay design. PLoS ONE 2018, 13, e0208862. [Google Scholar] [CrossRef] [Green Version]
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Golino, D.A.; Weber, E.; Sim, S.; Rowhani, A. Leafroll disease is spreading rapidly in a Napa Valley vineyard. Calif. Agric. 2008, 62, 156–160. [Google Scholar] [CrossRef] [Green Version]
- Cabaleiro, C.; Segura, A. Field transmission of grapevine leafroll associated virus 3 (GLRaV-3) by the mealybug Planococcus citri. Plant Dis. 1997, 81, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.L.; Charles, J.G. Transmission of grapevine leafroll-associated closteroviruses by Planococcus longispinus and P. calceolariae. Plant Pathol. 1997, 46, 509–515. [Google Scholar] [CrossRef]
- Sforza, R.; Boudon-Padieu, E.; Greif, C. New mealybug species vectoring grapevine leafroll-associated viruses-1 and-3 (GLRaV-1 and-3). Eur. J. Plant Pathol. 2003, 109, 975–981. [Google Scholar] [CrossRef]
- Manuel de Borbon, C.; Gracia, O.; Gomez Talquenca, G.S. Mealybugs and grapevine leafroll-associated virus 3 in vineyards of Mendoza, Argentina. Am. J. Enol. Vitic. 2004, 55, 283–285. [Google Scholar]
- Tsai, C.W.; Rowhani, A.; Golino, D.A.; Daane, K.M.; Almeida, R.P.P. Mealybug transmission of grapevine leafroll viruses: An analysis of virus-vector specificity. Phytopathology 2010, 100, 830–834. [Google Scholar] [CrossRef] [Green Version]
- Wistrom, C.M.; Blaisdell, G.K.; Wunderlich, L.R.; Almeida, R.P.P.; Daane, K.M. Ferrisia gilli (Hemiptera: Pseudococcidae) transmits grapevine leafroll-associated viruses. J. Econ. Entomol. 2016, 109, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.L.; Daugherty, M.P.; Jeske, D.R.; Almeida, R.P.P.; Daane, K.M. Incidence of grapevine leafroll disease: Effects of grape mealybug (Pseudococcus maritimus) abundance and pathogen supply. J. Econ. Entomol. 2018, 111, 1542–1550. [Google Scholar] [CrossRef]
- Daane, K.M.; Middleton, M.C.; Sforza, R.F.H.; Kamps-Hughes, N.; Watson, G.W.; Almeida, R.P.P.; Correa, M.C.G.; Downie, D.A.; Walton, V.M. Determining the geographic origin of invasive populations of the mealybug Planococcus ficus based on molecular genetic analysis. PLoS ONE 2018, 13, e0193852. [Google Scholar] [CrossRef] [Green Version]
- Walton, V.M.; Daane, K.M.; Addison, P. Biological control of arthropods and its application in vineyards. In Arthropod Management in Vineyards: Pests Approaches and Future Directions; Bostanian, N.J., Isaacs, R., Vincent, C., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 91–118. [Google Scholar]
- Charles, J.G.; Bell, V.A.; Lo, P.L.; Cole, L.M.; Chhagan, A. Mealybugs (Hemiptera: Pseudococcidae) and their natural enemies in New Zealand vineyards from 1993–2009. N. Z. Entomol. 2010, 33, 84–91. [Google Scholar] [CrossRef]
- Daane, K.M.; Malakar-Kuenen, R.D.; Walton, V.M. Temperature-dependent development of Anagyrus pseudococci (Hymenoptera:Encyrtidae) as a parasitoid of the vine mealybug, Planococcus ficus (Homoptera:Pseudococcidae). Biol. Control 2004, 31, 123–132. [Google Scholar] [CrossRef]
- Walton, V.M.; Pringle, K.L. A survey of mealybugs and associated natural enemies in vineyards in the Western Cape Province, South Africa. S. Afr. J. Enol. Vitic. 2004, 25, 23–25. [Google Scholar] [CrossRef]
- Triapitsyn, S.V.; Gonzalez, D.; Vickerman, D.B.; Noyes, J.S.; White, E.B. Morphological, biological, and molecular comparisons among the different geographical populations of Anagyrus pseudococci (Hymenoptera:Encyrtidae), parasitoids of Planococcus spp. (Hemiptera:Pseudococcidae), with notes on Anagyrus daetylopii. Biol. Control 2007, 41, 14–24. [Google Scholar] [CrossRef]
- Mahfoudhi, N.; Dhouibi, M.H. Survey of mealybugs (Hemiptera: Pseudococcidae) and their natural enemies in Tunisian vineyards. Afr. Entomol. 2009, 17, 154–160. [Google Scholar] [CrossRef]
- Daane, K.M.; Bentley, W.J.; Walton, V.M.; Malakar-Kuenen, R.; Millar, J.G.; Ingels, C.A.; Weber, E.A.; Gispert, C. New controls investigated for vine mealybug. Calif. Agric. 2006, 60, 31–38. [Google Scholar] [CrossRef]
- Sunitha, N.D.; Jagginavar, S.B.; Biradar, A.P. Bioefficacy botanicals and newer insecticides against grape vine mealybug, Maconellicoccus hirsutus (Green). Karnataka J. Agric. Sci. 2009, 22, 710–711. [Google Scholar]
- Lo, P.L.; Walker, J.T.S. Good results from a soil-applied insecticide against mealybugs. N. Z. Winegrower 2010, 14, 125–127. [Google Scholar]
- Mansour, R.; Belzunces, L.P.; Suma, P.; Zappala, L.; Mazzeo, G.; Grissa-Lebdi, K.; Russo, A.; Biondi, A. Vine and citrus mealybug pest control based on synthetic chemicals. A review. Agron. Sustain. Dev. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, T.; Prasad, K.S.; Shekhar, M.A.; Manjunath, D. Evaluation on neem based formulations vis-a-vis dichlorvos against Meconellicoccus hirsutus. Uttar Pradesh J. Zool. 2007, 27, 13–20. [Google Scholar]
- Satyanarayana, J.; Murthy, M.S.; Srinivasa, N. Impact of pesticides on grapevine mealybug predator, Cryptolaemus montrouzieri Mulsant. 1. Residual toxicity and safety of pesticidal sprays to predatory grubs. Indian J. Entomol. 1991, 53, 587–592. [Google Scholar]
- Walton, V.M.; Pringle, K.L. Effects of pesticides and fungicides used on grapevines on the mealybug predatory beetle Nephus ’boschianus’ (Coccinellidae, Scymnini). S. Afr. J. Enol. Vitic. 2001, 22, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Mgocheki, N.; Addison, P. Effect of contact pesticides on vine mealybug parasitoids, Anagyrus sp near pseudococci (Girault) and Coccidoxenoides perminutus (Timberlake) (Hymenoptera: Encyrtidae). S. Afr. J. Enol. Vitic. 2009, 30, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Daane, K.M.; Vincent, C.; Isaacs, R.; Ioriatti, C. Entomological opportunities and challenges for sustainable viticulture in a global market. Annu. Rev. Entomol. 2018, 63, 193–214. [Google Scholar] [CrossRef] [PubMed]
- Wilson, H.; Daane, K.M. Review of ecologically-based pest management in California vineyards. Insects 2017, 8, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical ecology and management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entomol. 2011, 104, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Bagnoli, B.; Cooper, M.; Ioriatti, C.; Varela, L. The successful use of sex pheromones to monitor and disrupt mating of Lobesia botrana in California. IOBC/WPRS Bull. 2014, 99, 45–48. [Google Scholar]
- Rotundo, G.; Tremblay, E. Studies on a sexual pheromone of Planococcus citri (Risso) (Homoptera, Coccoidea). I. Bollettino del Laboratorio di Entomologia Agraria ’Filippo Silvestri’ Portici 1972, 30, 217–230. [Google Scholar]
- Bierlleonhardt, B.A.; Moreno, D.S.; Schwarz, M.; Fargerlund, J.; Plimmer, J.R. Isolation, identification and synthesis of the sex pheromone of the citrus mealybug, Planococcus citri (Risso). Tetrahedron Lett. 1981, 22, 389–392. [Google Scholar] [CrossRef]
- Geiger, C.A.; Daane, K.M. Seasonal movement and distribution of the grape mealybug (Homoptera: Pseudococcidae): Developing a sampling program for San Joaquin Valley vineyards. J. Econ. Entomol. 2001, 94, 291–301. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Daane, K.M.; Ponti, L.; Walton, V.M.; Ellis, C.K. Prospective evaluation of the biological control of vine mealybug: Refuge effects and climate. J. Appl. Ecol. 2008, 45, 524–536. [Google Scholar] [CrossRef]
- Hinkens, D.M.; McElfresh, J.S.; Millar, J.G. Identification and synthesis of the sex pheromone of the vine mealybug, Planococcus ficus. Tetrahedron Lett. 2001, 42, 1619–1621. [Google Scholar] [CrossRef]
- Millar, J.G.; Midland, S.L.; McElfresh, J.S.; Daane, K.M. (2,3,4,4-Tetramethylcyclopentyl)methyl acetate, a sex pheromone from the obscure mealybug: First example of a new structural class of monoterpenes. J. Chem. Ecol. 2005, 31, 2999–3005. [Google Scholar] [CrossRef]
- Figadère, B.A.; McElfresh, J.S.; Borchardt, D.; Daane, K.M.; Bentley, W.; Millar, J.G. Trans-alpha-Necrodyl isobutyrate, the sex pheromone of the grape mealybug, Pseudococcus maritimus. Tetrahedron Lett. 2007, 48, 8434–8437. [Google Scholar] [CrossRef]
- Zou, Y.; Millar, J.G. Synthesis of the pheromone of the longtailed mealybug, a sterically congested, irregular monoterpenoid. J. Org. Chem. 2009, 74, 7207–7209. [Google Scholar] [CrossRef] [PubMed]
- Walton, V.M.; Daane, K.M.; Pringle, K.L. Monitoring Planococcus ficus in South African vineyards with sex pheromone-baited traps. Crop Protect. 2004, 23, 1089–1096. [Google Scholar] [CrossRef]
- Bahder, B.W.; Naidu, R.A.; Daane, K.M.; Millar, J.G.; Walsh, D.B. Pheromone-based monitoring of Pseudococcus maritimus (Hemiptera: Pseudococcidae) populations in Concord grape vineyards. J. Econ. Entomol. 2013, 106, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, J.G.; Daane, K.M.; McElfresh, J.S.; Moreira, J.A.; Malakar-Kuenen, R.; Guillen, M.; Bentley, W.J. Development and optimization of methods for using sex pheromone for monitoring the mealybug Planococcus ficus (Homoptera: Pseudococcidae) in California vineyards. J. Econ. Entomol. 2002, 95, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Walton, V.M.; Daane, K.M.; Bentley, W.J.; Millar, J.G.; Larsen, T.E.; Malakar-Kuenen, R. Pheromone-based mating disruption of Planococcus ficus (Hemiptera:Pseudococcidae) in California vineyards. J. Econ. Entomol. 2006, 99, 1280–1290. [Google Scholar] [CrossRef] [PubMed]
- Mansour, R.; Grissa-Lebdi, K.; Khemakhem, M.; Chaari, I.; Trabelsi, I.; Sabri, A.; Marti, S. Pheromone-mediated mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in Tunisian vineyards: Effect on insect population dynamics. Biologia 2017, 72, 333–341. [Google Scholar] [CrossRef]
- Sharon, R.; Zahavi, T.; Sokolsky, T.; Sofer-Arad, C.; Tomer, M.; Kedoshim, R.; Harari, A.R. Mating disruption method against the vine mealybug, Planococcus ficus: Effect of sequential treatment on infested vines. Entomol. Exp. Appl. 2016, 161, 65–69. [Google Scholar] [CrossRef]
- Shapira, I.; Keasar, T.; Harari, A.R.; Gavish-Regev, E.; Kishinevsky, M.; Steinitz, H.; Sofer-Arad, C.; Tomer, M.; Avraham, A.; Sharon, R. Does mating disruption of Planococcus ficus and Lobesia botrana affect the diversity, abundance and composition of natural enemies in Israeli vineyards? Pest Manag. Sci. 2018, 74, 1837–1844. [Google Scholar] [CrossRef]
- Lucchi, A.; Suma, P.; Ladurner, E.; Iodice, A.; Savino, F.; Ricciardi, R.; Cosci, F.; Marchesini, E.; Conte, G.; Benelli, G. Managing the vine mealybug, Planococcus ficus, through pheromone-mediated mating disruption. Environ. Sci. Pollut. Res. 2019, 26, 10708–10718. [Google Scholar] [CrossRef]
- Cocco, A.; Lentini, A.; Serra, G. Mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in vineyards using reservoir pheromone dispensers. J. Insect Sci. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Geiger, C.A.; Daane, K.M.; Bentley, W.J.; Yokota, G.Y.; Martin, L.A. Sampling program for grape mealybugs improves pest management. Calif. Agric. 2001, 55, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Cocco, A.; Muscas, E.; Mura, A.; Iodice, A.; Savino, F.; Lentini, A. Influence of mating disruption on the reproductive biology of the vine mealybug, Planococcus ficus (Hemiptera: Pseudococcidae), under field conditions. Pest Manag. Sci. 2018, 74, 2806–2816. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef] [PubMed]
- Kol-Maimon, H.; Levi-Zada, A.; Franco, J.C.; Dunkelblum, E.; Protasov, A.; Eliyaho, M.; Mendel, Z. Male behaviors reveal multiple pherotypes within vine mealybug Planococcus ficus (Signoret) (Hemiptera; Pseudococcidae) populations. Naturwissenschaften 2010, 97, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Miano, J.L.; Becerra, V.C.; Gonzalez, M.F. Mating disruption for Planococcus ficus Signoret: How to successfully initiate a novel sustainable control tool. Phytopathology 2011, 101, S120. [Google Scholar]
- Waterworth, R.A.; Wright, I.M.; Millar, J.G. Reproductive biology of three cosmopolitan mealybug (Hemiptera: Pseudococcidae) species, Pseudococcus longispinus, Pseudococcus viburni, and Planococcus ficus. Ann. Entomol. Soc. Am. 2011, 104, 249–260. [Google Scholar] [CrossRef]
- Franco, J.C.; Silva, E.B.; Cortegano, E.; Campos, L.; Branco, M.; Zada, A.; Mendel, Z. Kairomonal response of the parasitoid Anagyrus spec. nov near pseudococci to the sex pheromone of the vine mealybug. Entomol. Exp. Appl. 2008, 126, 122–130. [Google Scholar] [CrossRef]
- Shorey, H.H.; Sisk, C.B.; Gerber, R.G. Widely separated pheromone release sites for disruption of sex pheromone communication in two species of lepidoptera. Environ. Entomol. 1996, 25, 446–451. [Google Scholar] [CrossRef]
- Benelli, G.; Lucchi, A.; Thomson, D.; Ioriatti, C. Sex pheromone aerosol devices for mating disruption: Challenges for a brighter future. Insects 2019, 10, 308. [Google Scholar] [CrossRef] [Green Version]
- De Lame, F.M.; Epstein, D.; Gut, L.J.; Goldfarb, H.; Miller, J.R. Effect of Varying Dispenser Point Source Density on Mating Disruption of Grapholita molesta (Lepidoptera: Tortricidae). J. Econ. Entomol. 2010, 103, 1299–1305. [Google Scholar] [CrossRef] [PubMed]












© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daane, K.M.; Yokota, G.Y.; Walton, V.M.; Hogg, B.N.; Cooper, M.L.; Bentley, W.J.; Millar, J.G. Development of a Mating Disruption Program for a Mealybug, Planococcus ficus, in Vineyards. Insects 2020, 11, 635. https://doi.org/10.3390/insects11090635
Daane KM, Yokota GY, Walton VM, Hogg BN, Cooper ML, Bentley WJ, Millar JG. Development of a Mating Disruption Program for a Mealybug, Planococcus ficus, in Vineyards. Insects. 2020; 11(9):635. https://doi.org/10.3390/insects11090635
Chicago/Turabian StyleDaane, Kent M., Glenn Y. Yokota, Vaughn M. Walton, Brian N. Hogg, Monica L. Cooper, Walter J. Bentley, and Jocelyn G. Millar. 2020. "Development of a Mating Disruption Program for a Mealybug, Planococcus ficus, in Vineyards" Insects 11, no. 9: 635. https://doi.org/10.3390/insects11090635
APA StyleDaane, K. M., Yokota, G. Y., Walton, V. M., Hogg, B. N., Cooper, M. L., Bentley, W. J., & Millar, J. G. (2020). Development of a Mating Disruption Program for a Mealybug, Planococcus ficus, in Vineyards. Insects, 11(9), 635. https://doi.org/10.3390/insects11090635