New Genera and Species of the Family Throscidae (Coleoptera: Elateroidea) in Mid-Cretaceous Burmese Amber




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
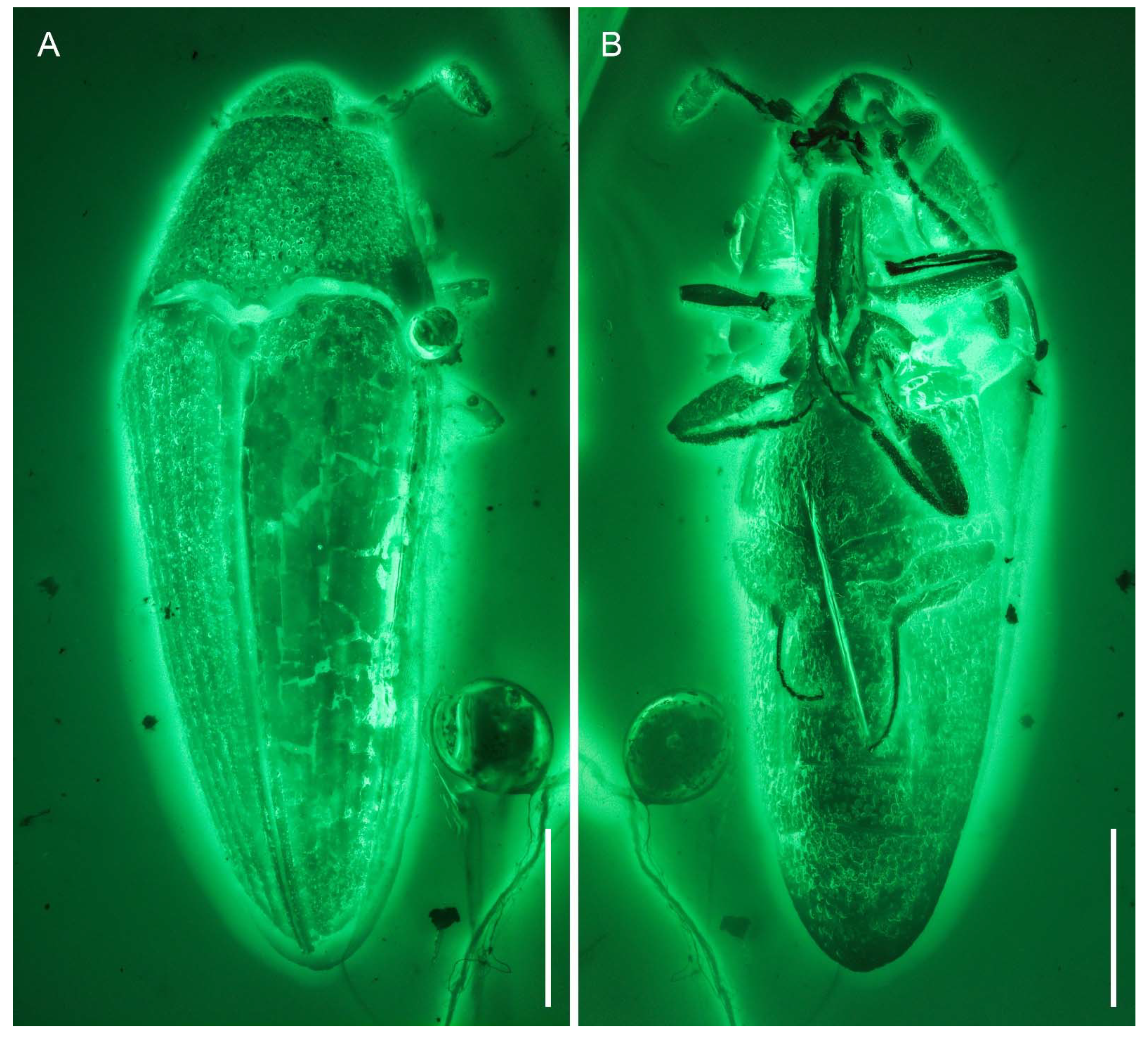
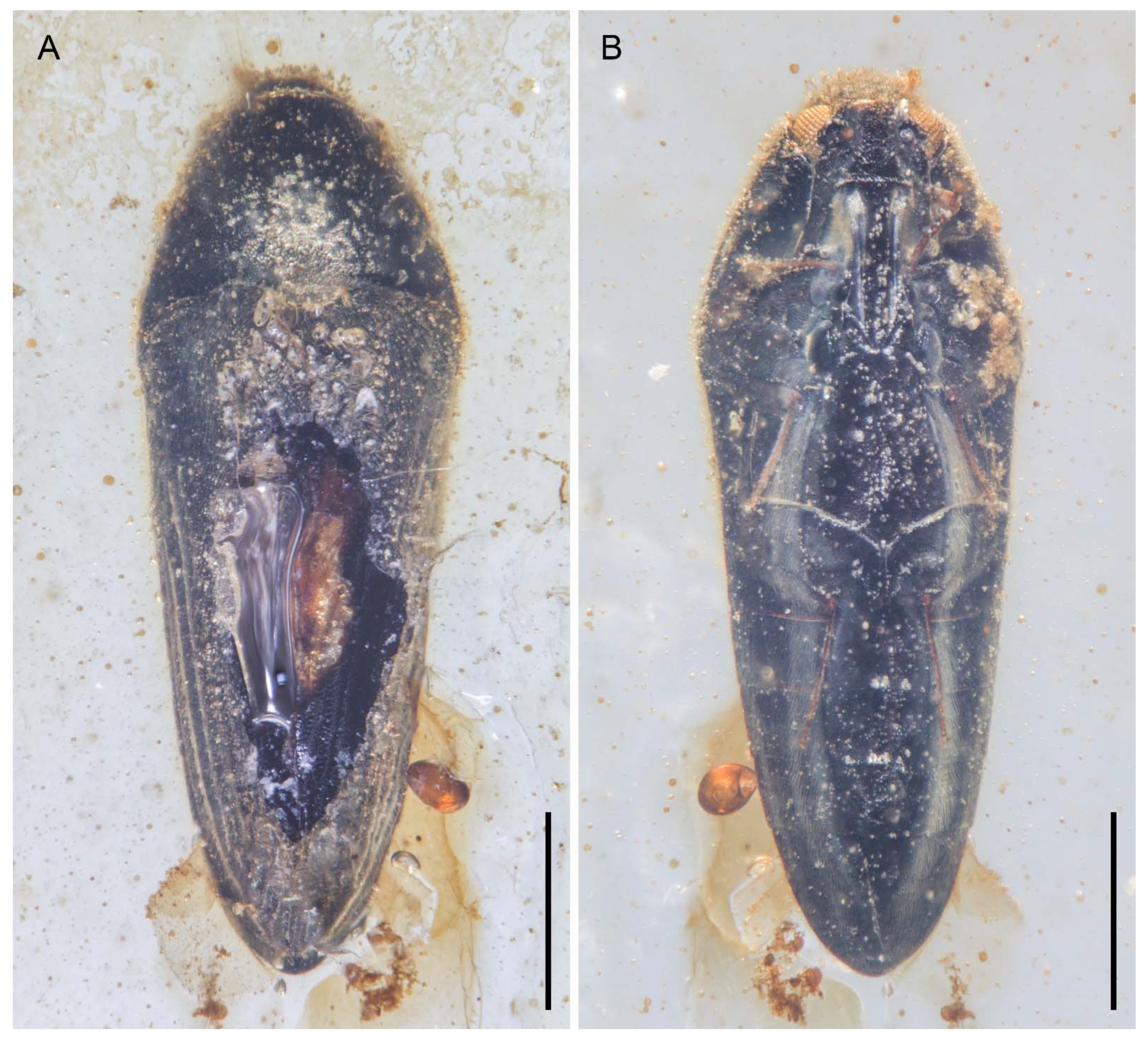
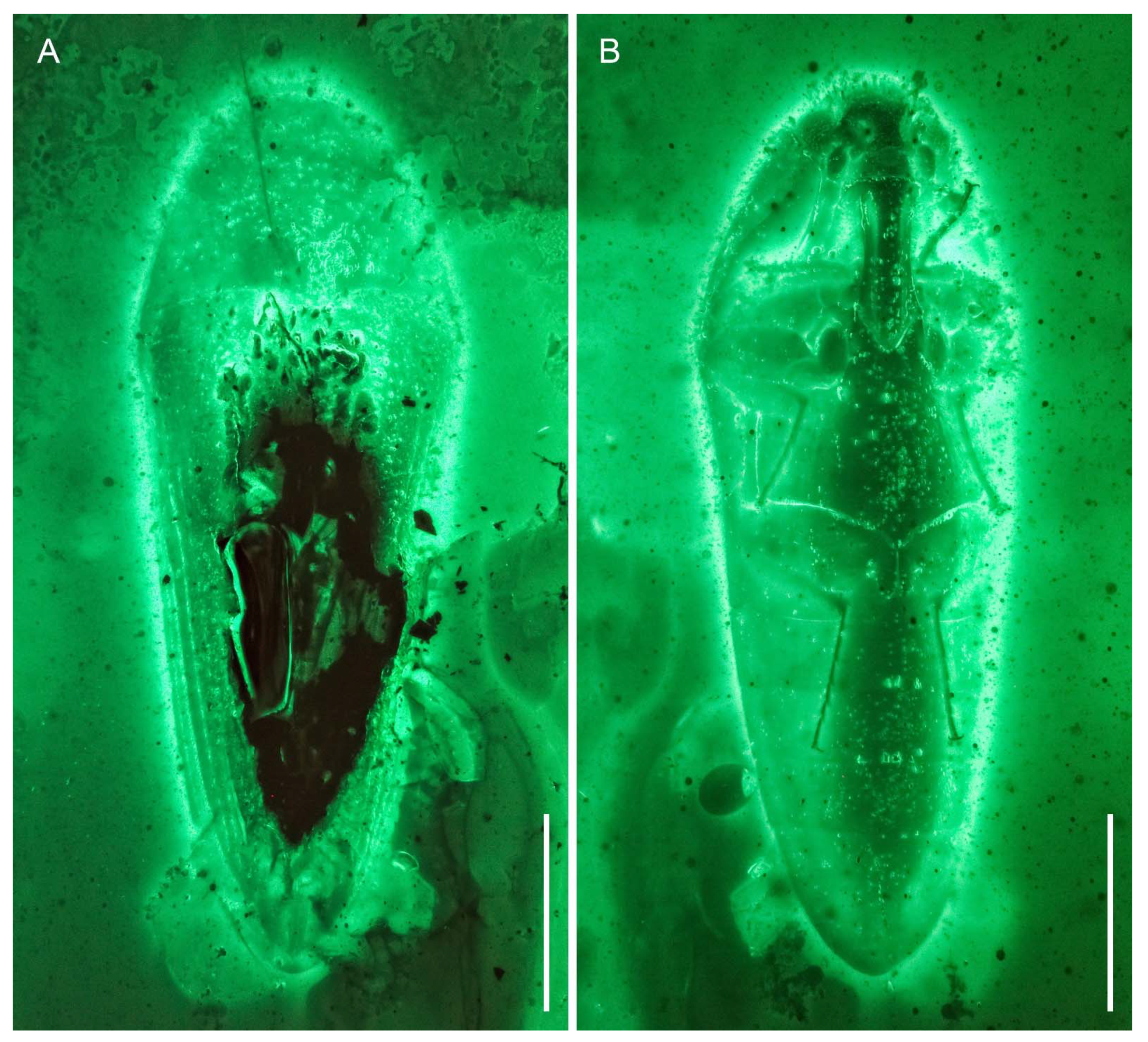
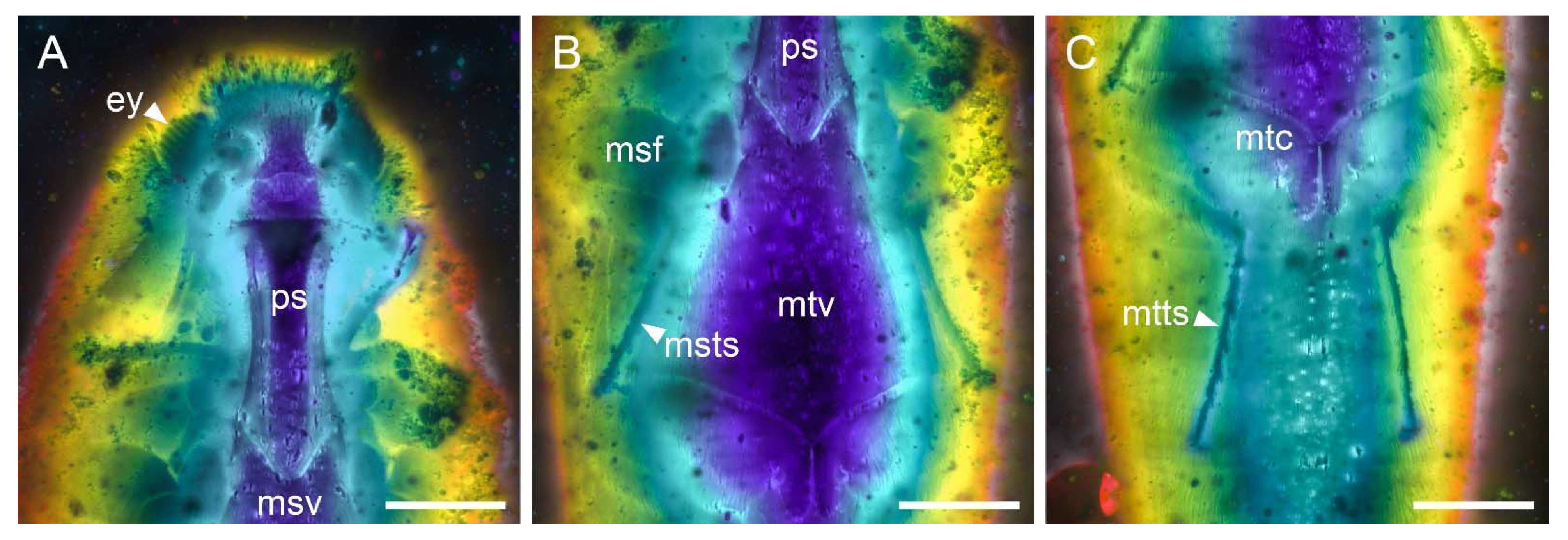
3. Systematic Palaeontology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Key to Genera of Throscidae (Modified from Muona, 2019)
| 1. Metathorax and abdomen without tarsal grooves; body elongate | 2 |
| 1′. At least metathorax with tarsal grooves | 5 |
| 2. Head without median carina | 3 |
| 2′. Head with median carina | 4 |
| 3. Prosternum with nonparallel prosternal carinae | Pseudothroscus Muona |
| 3′. Prosternum with subparallel prosternal carinae | Electrothroscus gen. nov. |
| 4. Antennae with 4 enlarged apical antennomeres | Jaira Muona |
| 4′. Antennae with 3 enlarged apical antennomeres | Potergosoma Kovalev & Kirejtshuk |
| 5. Abdomen with tarsal grooves | 6 |
| 5′. Abdomen at most with slight impressions for tarsi | 11 |
| 6. Head with median carina | 7 |
| 6′. Head without median carina | 8 |
| 7. Metaventrite simple | Potergus Bonvouloir |
| 7′. Metaventrite medially widely keeled, anteriorly deeply bifid | Rhomboaspis Kovalev & Kirejtshuk |
| 8. Supraocular ridges strong, well developed | Tyrannothroscus Muona |
| 8′. Supraocular ridges not present | 9 |
| 9. Frons with a pair of deep cavities | Captopus gen. nov. |
| 9′. Frons simple | 10 |
| 10. Protibiae apically enlarged, with tarsal grooves | Pactopus LeConte |
| 10′. Protibiae simple | Pseudopactopus gen. nov. |
| 11. Prosternum with nonparallel prosternal carinae | Trixagosoma Li et al. |
| 11′. Prosternum with subparallel prosternal carinae | 12 |
| 12. Metaventrite with vestigial tarsal grooves; median lobe longer than parameres | Triagus Kugelann |
| 12′. Metaventrite with deep, long tarsal grooves; median lobe shorter than parameres | Aulonothroscus Horn |
References
- Muona, J.; Lawrence, J.F.; Ślipiński, A. Throscidae Laporte, 1840. In Handbook of Zoology, Arthropoda: Insecta, Coleoptera, Beetles, Vol. 2: Morphology and Systematics (Elateroidea, Bostrichiformia, Cucujiformia Partim); Leschen, R.A.B., Beutel, R.G., Lawrence, J.F., Eds.; Walter de Gruyter: Berlin, Germany, 2010; pp. 69–74. [Google Scholar] [CrossRef]
- Kovalev, A.V.; Kirejtshuk, A.G.; Azar, D. The oldest representatives of the family Throscidae (Coleoptera: Elateriformia) from the Lower Cretaceous Lebanese amber. Cretac. Res. 2013, 44, 157–165. [Google Scholar] [CrossRef]
- Muona, J. Throscidae (Coleoptera) relationships, with descriptions of new fossil genera and species. Zootaxa 2019, 4576, 521–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobos, A. Estudios sobre Throscidae, I (Col. Sternoxia). Eos Rev. Esp. Entomol. 1963, 39, 135–176. [Google Scholar]
- Cobos, A. Estudios sobre Throscidae, II (Col. Sternoxia). Eos Rev. Esp. Entomol. 1967, 42, 311–351. [Google Scholar]
- Yensen, E. A revision of the North American species of Trixagus Kugelann (Coleoptera: Throscidae). Trans. Am. Entomol. Soc. 1975, 101, 125–166. [Google Scholar]
- Lawrence, J.F.; Newton, A.F. Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names). In Biology, Phylogeny, and Classification of Coleoptera: Papers Celebrating the 80th Birthday of Roy A. Crowson; Pakaluk, J., Ślipiński, S.A., Eds.; Muzeum i Instytut Zoologii PAN: Warsawa, Poland, 1995; pp. 779–1006. [Google Scholar]
- Lawrence, J.F.; Ślipiński, A.; Seago, A.E.; Thayer, M.K.; Newton, A.F.; Marvaldi, A.E. Phylogeny of the Coleoptera based on morphological characters of adults and larvae. Annal. Zool. 2011, 61, 1–217. [Google Scholar] [CrossRef]
- Kundrata, R.; Bocakova, M.; Bocak, L. The comprehensive phylogeny of the superfamily Elateroidea (Coleoptera: Elateriformia). Mol. Phylogenet. Evol. 2014, 76, 162–171. [Google Scholar] [CrossRef]
- Muona, J. The phylogeny of Elateroidea (Coleoptera), or which tree is best today? Cladistics 1995, 11, 317–341. [Google Scholar] [CrossRef]
- Kusy, D.; Motyka, M.; Andujar, C.; Bocek, M.; Masek, M.; Sklenarova, K.; Kokas, F.; Bocakova, M.; Vogler, A.P.; Bocak, L. Genome sequencing of Rhinorhipus Lawrence exposes an early branch of the Coleoptera. Front. Zool. 2018, 15, 21. [Google Scholar] [CrossRef]
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrich, F.; Letsch, H.; et al. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-D.; Zhao, Y.-C.; Huang, D.-Y.; Cai, C.-Y. A new throscid from mid-Cretaceous Burmese amber (Coleoptera: Elateroidea: Throscidae). Hist. Biol. 2020. [Google Scholar] [CrossRef]
- Kovalev, A.V.; Kirejtshuk, A.G.; Nel, A. New species of the genus Trixagus Kugelann, 1794 (Coleoptera: Throscidae) from the lowermost Eocene amber of Oise (France). Proc. Zool. Inst. RAS 2012, 316, 83–88. [Google Scholar]
- Muona, J. Eucnemidae and Throscidae in Baltic Amber. Entomol. Bl. 1993, 89, 15–45. [Google Scholar]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Ross, A.J. Supplement to the Burmese (Myanmar) amber checklist and bibliography, 2019. Palaeoentomology 2020, 3, 103–118. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Mao, Y.; Liang, K.; Su, Y.; Li, J.; Rao, X.; Zhang, H.; Xia, F.; Fu, Y.; Cai, C.; Huang, D. Various amberground marine animals on Burmese amber with discussions on its age. Palaeoentomology 2018, 1, 91–103. [Google Scholar] [CrossRef]
- Muona, J.; (Finnish Museum of Natural History, University of Helsinki, Helsinki, Finland). Personal communication, 2020.
- Yensen, E. A review of the genus Pactopus LeConte (Coleoptera: Throscidae). Coleopt. Bull. 1975, 29, 87–91. [Google Scholar]
- Wickham, H.F. Twenty new Coleoptera from the Florissant shales. Trans. Am. Entomol. Soc. 1914, 40, 257–270. [Google Scholar]
- Linard, B.; Crampton-Platt, A.; Moriniere, J.; Timmermans, M.J.T.N.; Andújar, C.; Arribas, P.; Miller, K.E.; Lipecki, J.; Favreau, E.; Hunter, A.; et al. The contribution of mitochondrial metagenomics to large-scale data mining and phylogenetic analysis of Coleoptera. Mol. Phylogenet. Evol. 2018, 128, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Clarke, D.J.; Limaye, A.; McKenna, D.D.; Oberprieler, R.G. The weevil fauna preserved in Burmese amber—Snapshot of a unique, extinct lineage (Coleoptera: Curculionoidea). Diversity 2019, 11, 1. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-D.; Huang, D.-Y.; Cai, C.-Y. New Genera and Species of the Family Throscidae (Coleoptera: Elateroidea) in Mid-Cretaceous Burmese Amber. Insects 2021, 12, 63. https://doi.org/10.3390/insects12010063
Li Y-D, Huang D-Y, Cai C-Y. New Genera and Species of the Family Throscidae (Coleoptera: Elateroidea) in Mid-Cretaceous Burmese Amber. Insects. 2021; 12(1):63. https://doi.org/10.3390/insects12010063
Chicago/Turabian StyleLi, Yan-Da, Di-Ying Huang, and Chen-Yang Cai. 2021. "New Genera and Species of the Family Throscidae (Coleoptera: Elateroidea) in Mid-Cretaceous Burmese Amber" Insects 12, no. 1: 63. https://doi.org/10.3390/insects12010063
APA StyleLi, Y. -D., Huang, D. -Y., & Cai, C. -Y. (2021). New Genera and Species of the Family Throscidae (Coleoptera: Elateroidea) in Mid-Cretaceous Burmese Amber. Insects, 12(1), 63. https://doi.org/10.3390/insects12010063