
Development of a LAMP-Based Molecular Species Diagnosis Method for Four Major Agricultural Pests in the Genus Spodoptera (Lepidoptera: Noctuidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Mitochondrial Genome Sequencing
2.2. Phylogenetic Analysis and Primer Design
2.3. LAMP and PCR
3. Results
3.1. Mitochondrial Genome Sequencing, Primer Design, and Selection
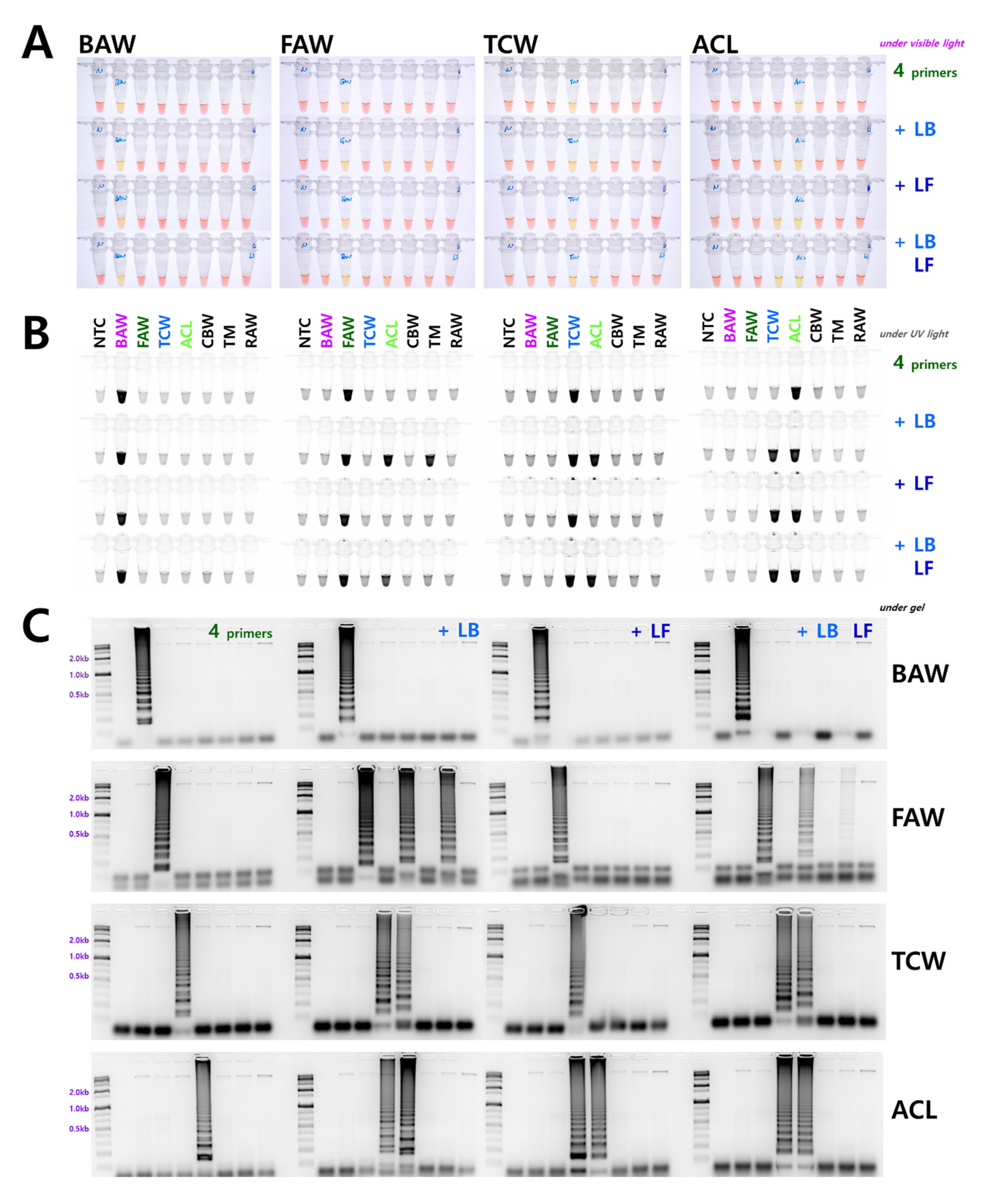
3.2. Comparison of Loop Primer Effectiveness
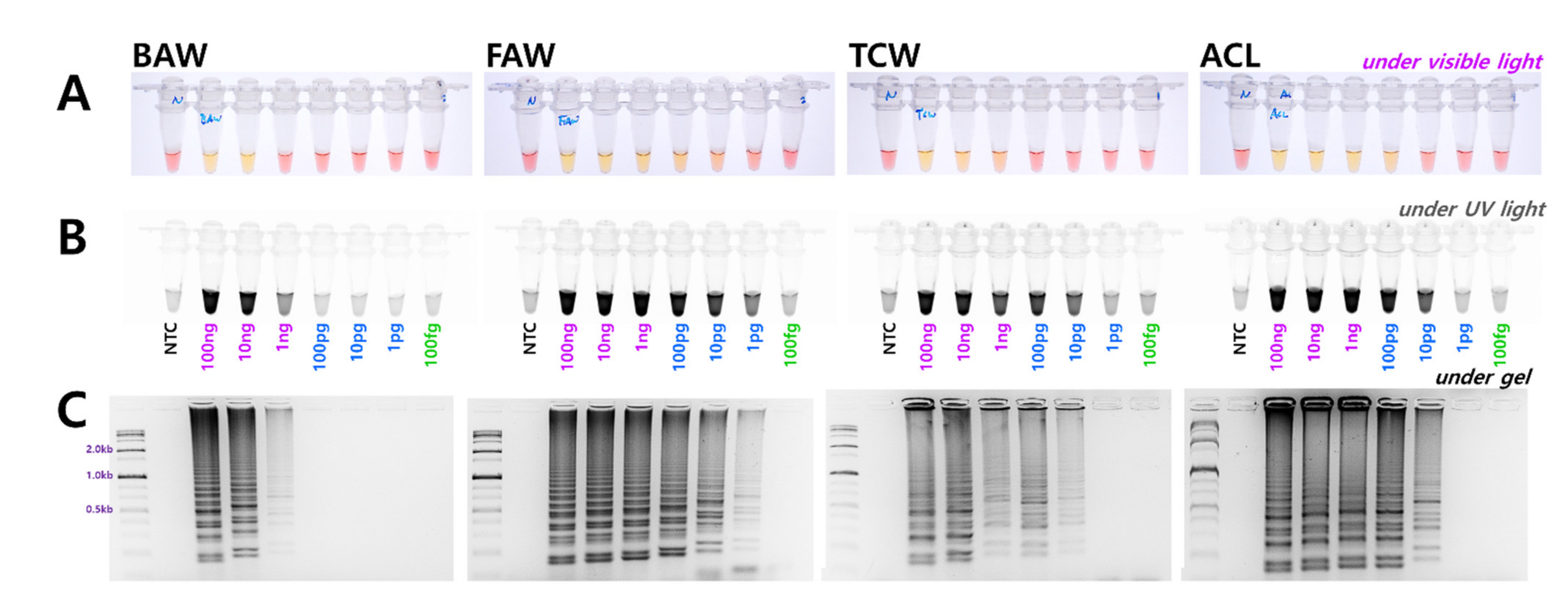
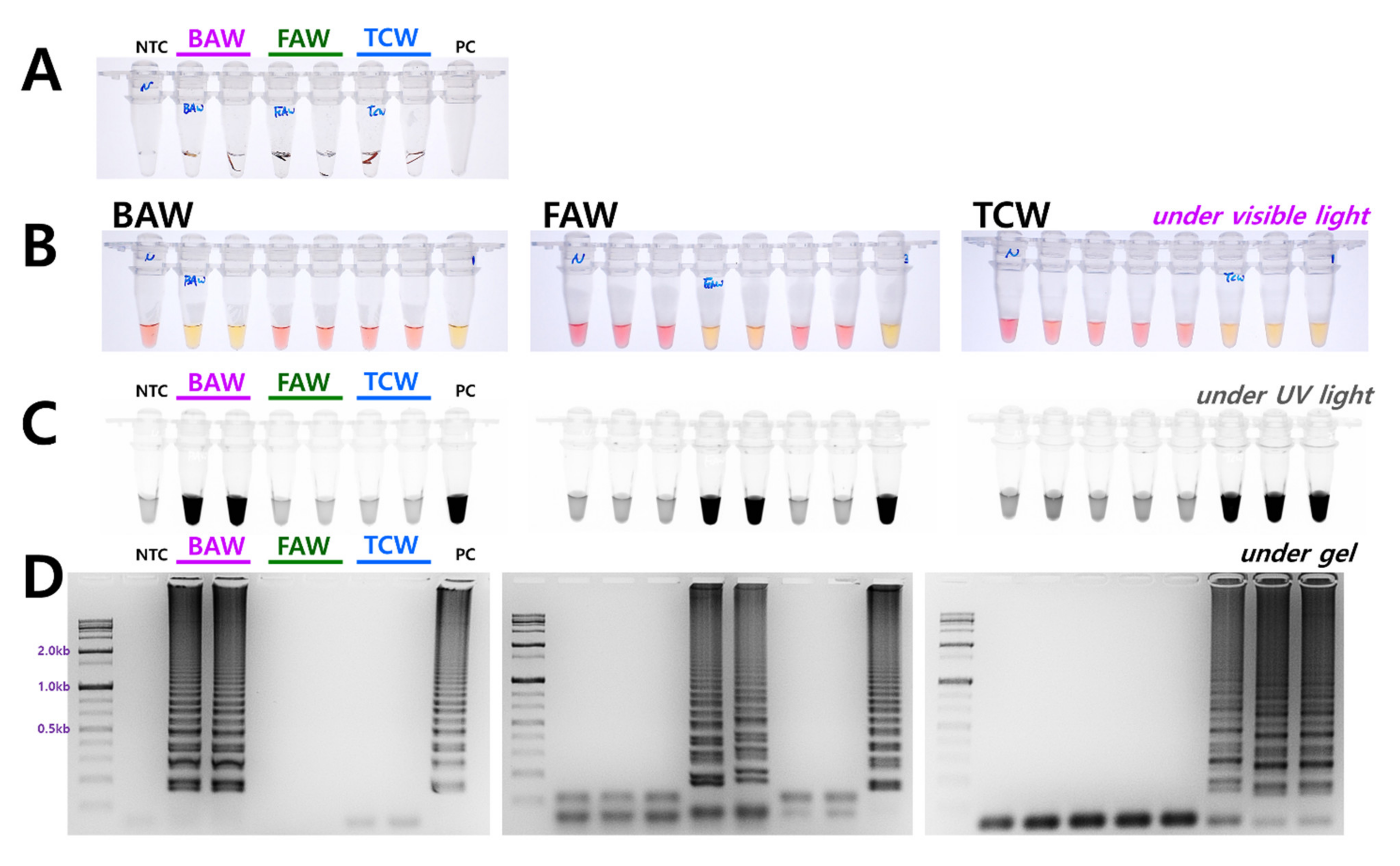
3.3. Determining the Diagnosis Limit Concentrations and Combining the DNA Release Technology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- CABI. Invasive Species Compendium. Available online: https://www.cabi.org/isc/search/index (accessed on 14 August 2020).
- Kim, J.; Nam, H.Y.; Kwon, M.; Choi, J.H.; Cho, S.R.; Kim, G.H. Novel diamide resistance-linked mutation in Korean Spodoptera exigua and a LAMP assay based on a mutation-associated intronic InDel. J. Pest Sci. 2021, 94, 1017–1029. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B.; Machial, C.M.; Miresmailli, S.; Bainard, L.D. Essential oil-based pesticides: New insights from old chemistry. In Pesticide Chemistry; Ohkawa, H., Miyagawa, H., Lee, P.W., Eds.; Wiley-VCH: Weinheim, Germany, 2007; pp. 201–209. [Google Scholar]
- Ahmad, M.; Mehmood, R. Monitoring of resistance to new chemistry insecticides in Spodoptera litura (Lepidoptera: Noctuidae) in Pakistan. J. Econ. Entomol. 2015, 108, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- EPPO. PM 7/124 (1) Spodoptera littoralis, Spodoptera litura, Spodoptera frugiperda, Spodoptera eridania. EPPO Bull. 2015, 45, 410–444. [Google Scholar] [CrossRef]
- Singab, M.; El-Hefny, A.; El-Hadek, M.K. Resistance ratio assessment to several bioinsecticides in cotton leafworm, Spodoptera littoralis at different governorates in Egypt. J. Plant Prot. Pathol. 2014, 5, 605–611. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafa, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Nam, H.Y.; Kwon, M.; Kim, H.J.; Yi, H.J.; Haenniger, S.; Unbehend, M.; Heckel, D.G. Development of a simple and accurate molecular tool for Spodoptera frugiperda species identification using LAMP. Pest Manag. Sci. 2021, 77, 3145–3153. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kwon, M.; Park, K.J.; Maharjanm, R. Monitoring of four major lepidopteran pests in Korean cornfields and management of Helicoverpa armigera. Entomol. Res. 2018, 48, 308–316. [Google Scholar] [CrossRef]
- Nam, H.Y.; Kwon, M.; Kim, H.J.; Kim, J. Development of a species diagnostic molecular tool for an invasive pest, Mythimna loreyi, using LAMP. Insects 2020, 11, 817. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.; Kent, W.J.; Smit, A.; Zhang, Z.; Baertsch, R.; Hardison, R.C.; Haussler, D.; Miller, W. Human-mouse alignments with BLASTZ. Genome Res. 2003, 13, 103–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, M.J.; Wojciechowski, M.F. Improved bootstrap confidence limits in large-scale phylogenies, with an example from Neo-Astragalus (Leguminosae). Syst. Biol. 2000, 49, 671–685. [Google Scholar] [CrossRef] [Green Version]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Mayor, C.; Brudno, M.; Schwartz, J.R.; Poliakov, A.; Rubin, E.M.; Frazer, K.A.; Patcher, L.S.; Dubchak, I. VISTA: Visualizing global DNA sequence alignments of arbitrary length. Bioinformatics 2000, 16, 1046–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badolo, A.; Okado, K.; Guelbeogo, W.M.; Aonuma, H.; Bando, H.; Fukumoto, S.; Sagnon, N.F.; Kanuka, H. Development of an allele-specific, loop-mediated, isothermal amplification method (AS-LAMP) to detect the L1014F kdr-w mutation in Anopheles gambiae s. l. Malar. J. 2012, 11, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badolo, A.; Bando, H.; Traoré, A.; Ko-Ketsu, M.; Guelbeogo, W.M.; Kanuka, H.; Ranson, H.; Fukumoto, S. Detection of G119S ace-1 R mutation in field-collected Anopheles gambiae mosquitoes using allele-specific loop-mediated isothermal amplification (AS-LAMP) method. Malar. J. 2015, 14, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purpose | Primers | Sequence (5’ → 3’) |
|---|---|---|
| for Spodoptera exigua (beet armyworm, BAW) | ||
| BAW_5F3 | GGAGCTATATGACCCCCTGTC 1 | |
| BAW_5B3 | TCCTGTTGCTATAAAGAATGTAGAC | |
| BAW_5FIP | GCTCAAGTAACTGATACTCCTGATCTAATTTAATCCATTTCAAATTCCTCT | |
| BAW_5BIP | ATTCCCAAATAACTCAAGGACTATTCTTCAAAATATTCATATGCTTGT | |
| BAW_5LF | CTGATCTAATTAAAATAATAGTATTAAG | |
| BAW_5LB | TTACAATTATTTTAGGAATTTATTTTAC | |
| for Spodoptera frugiperda (fall armyworm, FAW) | ||
| FAW_1F3 | TTCTAGTAGGATATTTAATATCAAGCT | |
| FAW_1B3 | CGAAAATTATTACTTGATATATCATTTGTA | |
| FAW_1FIP | CTGGTAAAATAGTTCAAATTAATAATATATTAATCGATTTTTATTAGAAGGTC | |
| FAW_1BIP | CCTTCACTACGATTATTATATTTATTAGAACTTCAATATCATTGATGACCA | |
| FAW_1LF | GTAAAATAGTTCAAATTAATTCAATTATTT | |
| FAW_1LB | CTTAATAATCCTTTAATTACATTAAAATC | |
| for Spodoptera litura (tobacco cutworm, TCW) | ||
| TCW_3F3 | GTTTCTTTACCTTTATGATTAAGTTTC 2 | |
| TCW_3B3 | ATAATCTACTAAATGGAAAGGATGG | |
| TCW_3FIP | GGTATTAAAATAGTGGGTGTTCCTTGATTAAATAACTCTCAACATATATTCATTCA | |
| TCW_3BIP | TTGAAACTATTAGAAATATTATTCGACCGACCTGCAATCATATTAGCTGTT | |
| TCW_3LF | GGGTGTTCCTTGAGGAATTATA | |
| TCW_3LB | GGGAACATTAGCAGTTCGATT | |
| for Spodoptera littoralis (African cotton leafworm, ACL) | ||
| ACL_2F3 | CCTTACCATTATGATTAAGCTTTATG | |
| ACL_2B3 | GACTATTCCTCTAACTAAAACTATTGT | |
| ACL_2FIP | AGTTATTAATAAATGACCTGCGATTATATTAATATTATTCGACCAGGAACAT | |
| ACL_2BIP | CTGGACCTTCTATACCAAATTATTTTATATAACCGCAACTGCTGAT | |
| ACL_2LF | AGCTGTTAATCGAACTGCTA | |
| ACL_2LB | TAATTCAAATTTTATTATTAATCTTAGA | |
| for PCR | ||
| LCO1490 | GGTCAACAAATCATAAAGATATTGG | |
| HCO2198 | TAAACTTCAGGCTGACCAAAAAATCA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, H.Y.; Kim, J.H.; Lee, S.H.; Heckel, D.G.; Kim, J. Development of a LAMP-Based Molecular Species Diagnosis Method for Four Major Agricultural Pests in the Genus Spodoptera (Lepidoptera: Noctuidae). Insects 2021, 12, 883. https://doi.org/10.3390/insects12100883
Nam HY, Kim JH, Lee SH, Heckel DG, Kim J. Development of a LAMP-Based Molecular Species Diagnosis Method for Four Major Agricultural Pests in the Genus Spodoptera (Lepidoptera: Noctuidae). Insects. 2021; 12(10):883. https://doi.org/10.3390/insects12100883
Chicago/Turabian StyleNam, Hwa Yeun, Ju Hyeon Kim, Si Hyeock Lee, David G. Heckel, and Juil Kim. 2021. "Development of a LAMP-Based Molecular Species Diagnosis Method for Four Major Agricultural Pests in the Genus Spodoptera (Lepidoptera: Noctuidae)" Insects 12, no. 10: 883. https://doi.org/10.3390/insects12100883
APA StyleNam, H. Y., Kim, J. H., Lee, S. H., Heckel, D. G., & Kim, J. (2021). Development of a LAMP-Based Molecular Species Diagnosis Method for Four Major Agricultural Pests in the Genus Spodoptera (Lepidoptera: Noctuidae). Insects, 12(10), 883. https://doi.org/10.3390/insects12100883