
Bumble Bee Foraged Pollen Analyses in Spring Time in Southern Estonia Shows Abundant Food Sources



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Environment
2.2. Hive Locations
2.3. Bumble Bees
2.4. Collection of Pollen
2.5. The Plant Species Determination of Pollen
2.5.1. Light Microscopy
2.5.2. DNA Metabarcoding
2.6. Statistical Analyses
3. Results
3.1. Taxonomic Variability
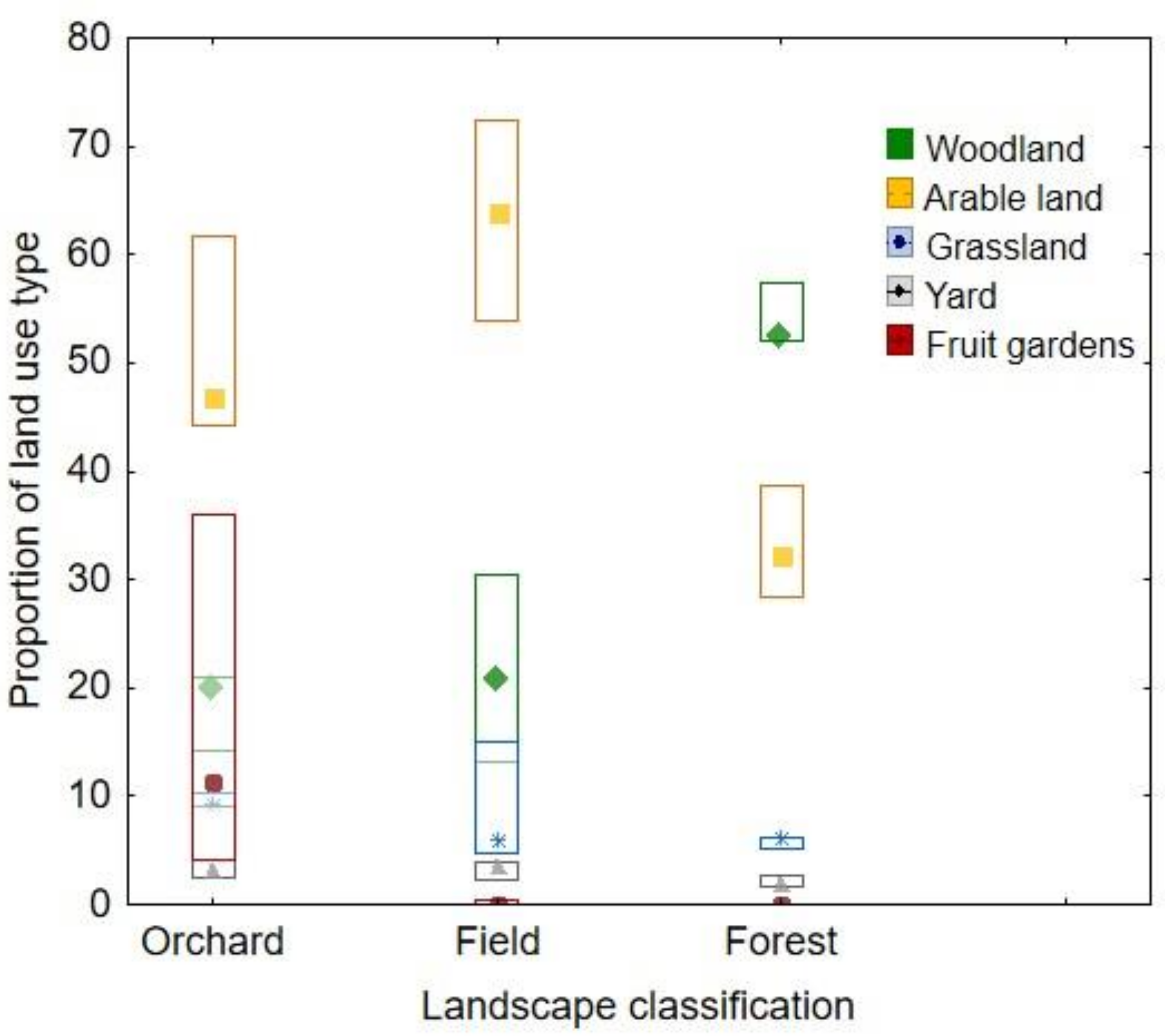
3.2. Proportional Variability of Pollens between Landscape Categories (Plant Family Level)
3.3. Correlations of Pollen Proportions and Share of Land Use Parameters (Plant Family Level)
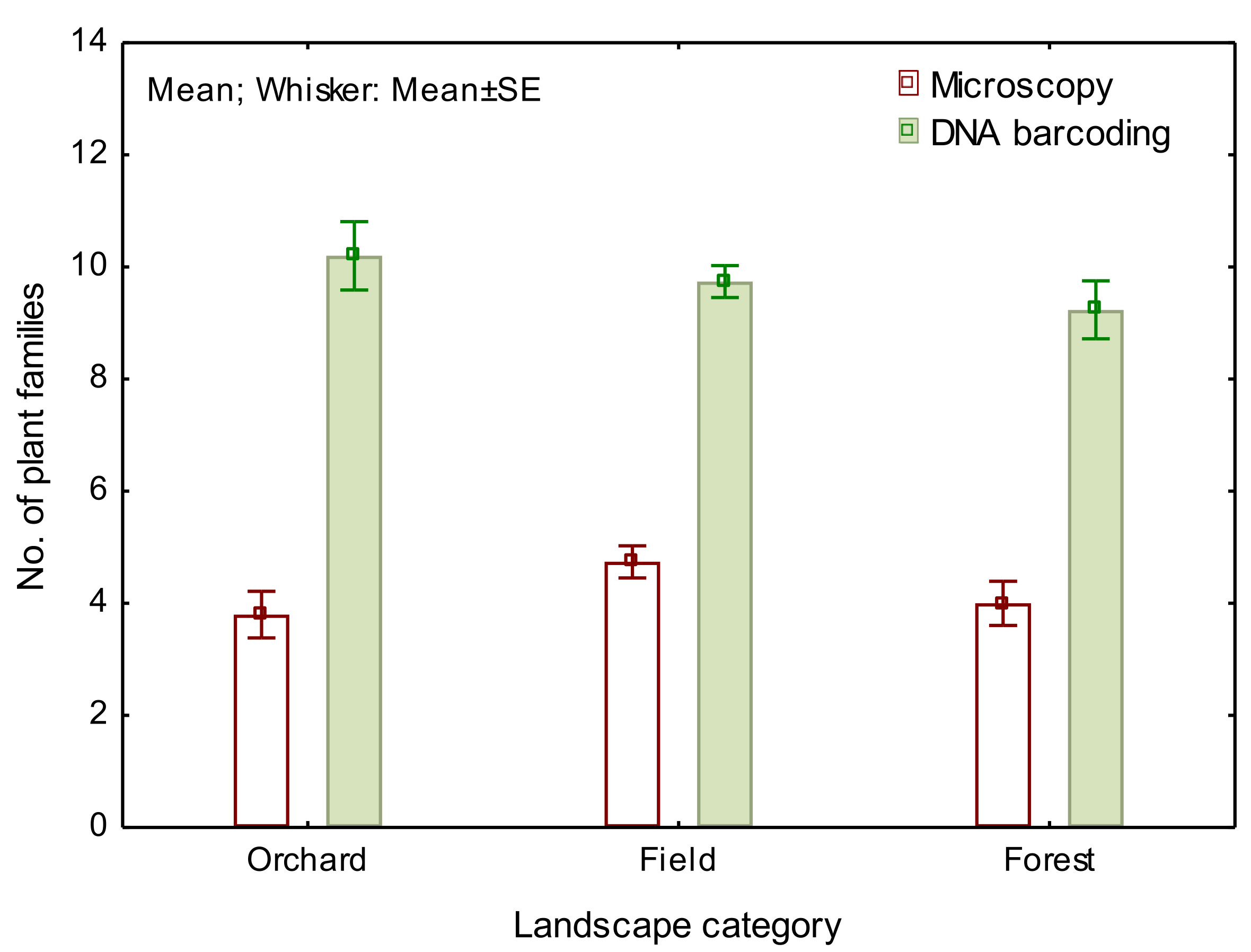
3.4. Reliability of DNA Metabarcoding Quantification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Underwood, E.; Darwin, G.; Gerritsen, E. Pollinator Initiatives in EU Member States Success Factors and Gaps. Report for European Commission under Contract for Provision of Technical Support Related to Target 2 of the EU Biodiversity Strategy to 2020—Maintaining and Restoring Ecosystems and Their Services ENV.B.2/SER/2016/0018; Institute for European Environmental Policy: Brussels, Belgium, 2017; p. 70. [Google Scholar]
- Science for Environment Policy. Pollinators: Importance for Nature and Human Well-Being, Drivers of Decline and the Need for Monitoring. Future Brief 23. Brief Produced for the European Commission DG Environment; Science Communication Unit, UWE Bristol: Bristol, UK, 2020. [Google Scholar]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.; Dicks, L.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef]
- Avarguès-Weber, A.; Lachlan, R.; Chittka, L. Bumblebee social learning can lead to suboptimal foraging choices. Anim. Behav. 2018, 135, 209–214. [Google Scholar] [CrossRef]
- Harmon-Threatt, A.N.; de Valpine, P.; Kremen, C. Estimating resource preferences of a native bumblebee: The effects of availability and use–availability models on preference estimates. Oikos 2017, 126, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Ogilvie, J.E.; Forrest, J.R. Interactions between bee foraging and floral resource phenology shape bee populations and communities. Curr. Opin. Insect. Sci. 2017, 21, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Vaudo, A.D.; Patch, H.M.; Mortensen, D.A.; Tooker, J.F.; Grozinger, C.M. Macronutrient ratios in pollen shape bumble bee (Bombus impatiens) foraging strategies and floral preferences. Proc. Natl. Acad. Sci. USA 2016, 113, 4035–4042. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Bonoan, R.E.; Gonzalez, J.; Starks, P.T. The perils of forcing a generalist to be a specialist: Lack of dietary essential amino acids impacts honey bee pollen foraging and colony growth. J. Apic. Res. 2020, 59, 95–103. [Google Scholar] [CrossRef]
- Smart, M.D.; Otto, C.R.V.; Lundgren, J.G. Nutritional status of honey bee (Apis mellifera L.) workers across an agricultural land-use gradient. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Park, M.G.; Blitzer, E.J.; Gibbs, J.; Losey, J.E.; Danforth, B.N. Negative effects of pesticides on wild bee communities can be buffered by landscape context. Proc. R. Soc. B 2015, 282, 20150299. [Google Scholar] [CrossRef] [Green Version]
- Marja, R.; Herzon, I.; Viik, E.; Elts, J.; Maend, M.; Tscharntke, T.; Batary, P. Environmentally friendly management as an intermediate strategy between organic and conventional agriculture to support biodiversity. Biol. Conserv. 2014, 178, 146–154. [Google Scholar] [CrossRef]
- Marja, R.; Kleijn, D.; Tscharntke, T.; Klein, A.-M.; Frank, T.; Batáry, P. Effectiveness of agri-environmental management on pollinators is moderated more by ecological contrast than by landscape structure or land-use intensity. Ecol. Lett. 2019, 22, 1493–1500. [Google Scholar] [CrossRef]
- Raimets, R.; Bontsutsnaja, A.; Bartkevics, V.; Pugajeva, I.; Kaart, T.; Puusepp, L.; Pihlik, P.; Keres, I.; Viinalass, H.; Mänd, M.; et al. Pesticide residues in beehive matrices are dependent on collection time and matrix type but independent of proportion of foraged oilseed rape and agricultural land in foraging territory. Chemosphere 2020, 238, 124555. [Google Scholar] [CrossRef] [PubMed]
- Westphal, C.; Steffan-Dewenter, I.; Tscharntke, T. Foraging trip duration of bumblebees in relation to landscape-wide resource availability. Ecol. Entomol. 2006, 31, 389–394. [Google Scholar] [CrossRef]
- Goulson, D.; Lepais, O.; O’Connor, S.; Osborne, J.L.; Sanderson, R.A.; Cussans, J.; Goffe, L.; Darvill, B. Effects of land use at a landscape scale on bumblebee nest density and survival. J. Appl. Ecol. 2010, 47, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Rahl, M. Microscopic identification and purity determination of pollen grains. Methods Mol. Med. 2008, 138, 263–269. [Google Scholar] [PubMed]
- Keller, A.; Danner, N.; Grimmer, G.; Ankenbrand, M.; von der Ohe, K.; Rost, S.; Härtel, S.; Steffan-Dewenter, I. Evaluating multiplexed next-generation sequencing as a method in palynology for mixed pollen samples. Plant. Biol. Stuttg. Ger. 2015, 17, 558–566. [Google Scholar] [CrossRef] [Green Version]
- Richardson, R.T.; Lin, C.-H.; Quijia, J.O.; Riusech, N.S.; Goodell, K.; Johnson, R. Rank-based characterization of pollen assemblages collected by honey bees using a multi-locus metabarcoding approach1. Appl. Plant Sci. 2015, 3, 1500043. [Google Scholar] [CrossRef]
- Sickel, W.; Ankenbrand, M.J.; Grimmer, G.; Holzschuh, A.; Härtel, S.; Lanzen, J.; Steffan-Dewenter, I.; Keller, A. Increased efficiency in identifying mixed pollen samples by meta-barcoding with a dual-indexing approach. BMC Ecol. 2015, 15, 20. [Google Scholar] [CrossRef] [Green Version]
- Smart, M.D.; Cornman, R.S.; Iwanowicz, D.D.; McDermott-Kubeczko, M.; Pettis, J.S.; Spivak, M.S.; Otto, C. A comparison of honey bee-collected pollen from working agricultural lands using light microscopy and ITS metabarcoding. Environ. Entomol. 2017, 46, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Richardson, R.T.; Lin, C.-H.; Sponsler, D.B.; Quijia, J.O.; Goodell, K.; Johnson, R.M. Application of ITS2 metabarcoding to determine the provenance of pollen collected by honey bees in an agroecosystem. Appl. Plant Sci. 2015, 3, apps.1400066. [Google Scholar] [CrossRef]
- Bell, K.L.; Burgess, K.S.; Botsch, J.C.; Dobbs, E.K.; Read, T.D.; Brosi, B.J. Quantitative and qualitative assessment of pollen DNA metabarcoding using constructed species mixtures. Mol. Ecol. 2019, 28, 431–455. [Google Scholar] [CrossRef] [PubMed]
- Loit, K.; Soonvald, L.; Astover, A.; Runno-Paurson, E.; Öpik, M.; Tedersoo, L. Temporal and cultivar-specific effects on potato root and soil fungal diversity. Agronomy 2020, 10, 1535. [Google Scholar] [CrossRef]
- Parmentier, L.; Meeus, I.; Cheroutre, L.; Mommaerts, V.; Louwye, S.; Smagghe, G. Commercial bumblebee hives to assess an anthropogenic environment for pollinator support: A case study in the region of Ghent (Belgium). Environ. Monit. Assess. 2014, 186, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Statistical Office of Estonia (Ed.). 2000 Population and Housing Census, I. Population De Facto and Usual Resident Population, Location of the Population, Population Sex and Age Structure; Statistikaamet: Tallinn, Estonia, 2001. [Google Scholar]
- Statistical Office of Estonia (Ed). 2000 Population and Housing Census. Population Settlements of Rural Municipalities; Statistikaamet: Tallinn, Estonia, 2001. [Google Scholar]
- Karise, R.; Dreyersdorff, G.; Jahani, M.; Veromann, E.; Runno-Paurson, E.; Kaart, T.; Smagghe, G.; Mänd, M. Reliability of the entomovector technology using Prestop-Mix and Bombus terrestris L. as a fungal disease biocontrol method in open field. Sci. Rep. 2016, 6, 31650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulis, E.S.J.; Goulson, D. Commercial bumble bees on soft fruit farms collect pollen mainly from wildflowers rather than the target crops. J. Apic. Res. 2014, 53, 404–407. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Carreck, N.L.; Swain, J.L.; Knight, M.E.; Goulson, D.; Hale, R.J.; Sanderson, R.A. Bumblebee flight distances in relation to the forage landscape. J. Anim. Ecol. 2008, 77, 406–415. [Google Scholar] [CrossRef]
- Diaz-Forero, I.; Kuusemets, V.; Maend, M.; Liivamaegi, A.; Kaart, T.; Luig, J. Effects of Forest Habitats on the Local Abundance of Bumblebee Species: A Landscape-scale Study. Balt. For. 2011, 17, 235–242. [Google Scholar]
- Walther-Hellwig, K.; Frankl, R. Foraging Distances of Bombus muscorum, Bombus lapidarius, and Bombus terrestris (Hymenoptera, Apidae). J. Insect Behav. 2000, 13, 239–246. [Google Scholar] [CrossRef]
- Walther-Hellwig, K.; Frankl, R. Foraging habitats and foraging distances of bumblebees, Bombus spp. (Hym., Apidae), in an agricultural landscape. J. Appl. Entomol. 2000, 124, 299–306. [Google Scholar] [CrossRef]
- Kreyer, D.; Oed, A.; Walther-Hellwig, K.; Frankl, R. Are forests potential landscape barriers for foraging bumblebees? Landscape scale experiments with Bombus terrestris agg. and Bombus pascuorum (Hymenoptera, Apidae). Biol. Conserv. 2004, 116, 111–118. [Google Scholar] [CrossRef]
- Ruedenauer, F.; Raubenheimer, D.; Kessner-Beierlein, D.; Grund-Mueller, N.; Noack, L.; Spaethe, J.; Leonhardt, S. Best be(e) on low fat: Linking nutrient perception, regulation and fitness. Ecol. Lett. 2020, 23, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, M.; Halbritter, H.; Weber, M.; Buchner, R.; Frosch-Radivo, A.; Ulrich, S.; Zetter, R. Pollen Terminology; Springer: Wien, Austria, 2009; Volume VII, p. 266. [Google Scholar] [CrossRef]
- D’Albore, R.G. An Illustrated Handbook. Mediterranean Melissopalynology; Università degli studi di Perugia, Facoltà di agraria, Istituto di entomologia agrarian: Perugia, Italy, 1998. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Koetschan, C.; Förster, F.; Keller, A.; Schleicher, T.; Ruderisch, B.; Schwarz, R.; Müller, T.; Wolf, M.; Schultz, J. The ITS2 Database III—sequences and structures for phylogeny. Nucleic Acids Res. 2010, 38, D275–D279. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D. The conservation of bumble bees. Bee World 2003, 84, 105–106. [Google Scholar] [CrossRef]
- Karise, R.; Mänd, M.; Ivask, M.; Koskor, E.; Bender, A. The effect of pollen amount and its caloric value in hybrid lucerne (Medicago x varia) on its attractiveness to bumble bees (Bombus terrestris). Agron. Res. 2006, 4, 211–216. [Google Scholar]
- Somme, L.; Vanderplanck, M.; Michez, D.; Lombaerde, I.; Moerman, R.; Wathelet, B.; Wattiez, R.; Lognay, G.; Jacquemart, A.-L. Pollen and nectar quality drive the major and minor floral choices of bumble bees. Apidologie 2014, 46, 92–106. [Google Scholar] [CrossRef]
- Evans, L.J.; Raine, N.E. Changes in learning and foraging behaviour within developing bumble bee (bombus terrestris) colonies. PLoS ONE 2014, 9, e90556. [Google Scholar] [CrossRef]
- Grüter, C.; Moore, H.; Firmin, N.; Helanterä, H.; Ratnieks, F.L.W. Flower constancy in honey bee workers (Apis mellifera) depends on ecologically realistic rewards. J. Exp. Biol. 2011, 214, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Jeon, H.; Jung, C. Foraging behaviour and preference of pollen sources by honey bee (Apis mellifera) relative to protein contents. J. Ecol. Environ. 2020, 44, 4. [Google Scholar] [CrossRef]
- Gijbels, P.; Ceulemans, T.; Van den Ende, W.; Honnay, O. Experimental fertilization increases amino acid content in floral nectar, fruit set and degree of selfing in the orchid Gymnadenia conopsea. Oecologia 2015, 179, 785–795. [Google Scholar] [CrossRef]
- Viik, E.; Mänd, M.; Karise, R.; Lääniste, P.; Williams, I.H.; Luik, A. The impact of foliar fertilization on the number of bees (Apoidea) on spring oilseed rape. Agriculture 2012, 99, 41–46. [Google Scholar]
- Hülsmann, M.; von Wehrden, H.; Klein, A.-M.; Leonhardt, S.D. Plant diversity and composition compensate for negative effects of urbanization on foraging bumble bees. Apidologie 2015, 46, 760–770. [Google Scholar] [CrossRef]
- Kleijn, D.; Raemakers, I. A retrospective analysis of pollen host plant use by stable and declining bumble bee species. Ecology 2008, 89, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.J.; Gibbs, J.; Graham, K.K.; Isaacs, R. Narrow pollen diets are associated with declining Midwestern bumble bee species. Ecology 2019, 100, e02697. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Tooker, J.F.; Patch, H.M.; Biddinger, D.J.; Coccia, M.; Crone, M.K.; Fiely, M.; Francis, J.S.; Hines, H.M.; Hodges, M.; et al. Pollen protein: Lipid macronutrient ratios may guide broad patterns of bee species floral preferences. Insects 2020, 11, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallioniemi, E.; Åström, J.; Rusch, G.M.; Dahle, S.; Åström, S.; Gjershaug, J.O. Local resources, linear elements and mass-flowering crops determine bumblebee occurrences in moderately intensified farmlands. Agric. Ecosyst. Environ. 2017, 239, 90–100. [Google Scholar] [CrossRef]
- Macukanovic, M.; Blazencic, Z. Diurnal and seasonal dynamics of nectar secretion of some species in the family Lamiaceae. Acta Veterinaria. 1999, 48, pp. 173–184. Available online: https://agris.fao.org/agris-search/search.do?recordID=YU1998000854 (accessed on 8 June 2021).
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Lye, G.; Park, K.; Osborne, J.; Holland, J.; Goulson, D. Assessing the value of Rural Stewardship schemes for providing foraging resources and nesting habitat for bumblebee queens (Hymenoptera: Apidae). Biol. Conserv. 2009, 142, 2023–2032. [Google Scholar] [CrossRef]
- Mola, J.M.; Richardson, L.L.; Spyreas, G.; Zaya, D.N.; Pearse, I.S. Long-term surveys support declines in early season forest plants used by bumblebees. J. Appl. Ecol. 2021, 58, 1431–1441. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Shortall, C.R.; Todd, A.D.; Goulson, D.; Knight, M.E.; Hale, R.J.; Sanderson, R.A. Quantifying and comparing bumblebee nest densities in gardens and countryside habitats. J. Appl. Ecol. 2008, 45, 784–792. [Google Scholar] [CrossRef]
- Gous, A.; Swanevelder, D.Z.H.; Eardley, C.D.; Willows-Munro, S. Plant–pollinator interactions over time: Pollen metabarcoding from bees in a historic collection. Evol. Appl. 2019, 12, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresty, C.E.A.; Clare, E.; Devey, D.S.; Cowan, R.S.; Csiba, L.; Malakasi, P.; Lewis, O.T.; Willis, K. Flower preferences and pollen transport networks for cavity-nesting solitary bees: Implications for the design of agri-environment schemes. Ecol. Evol. 2018, 8, 7574–7587. [Google Scholar] [CrossRef]
- Suchan, T.; Talavera, G.; Sáez, L.; Ronikier, M.; Vila, R. Pollen metabarcoding as a tool for tracking long-distance insect migrations. Mol. Ecol. Resour. 2019, 19, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bänsch, S.; Tscharntke, T.; Wünschiers, R.; Netter, L.; Brenig, B.; Gabriel, D.; Westphal, C. Using ITS2 metabarcoding and microscopy to analyse shifts in pollen diets of honey bees and bumble bees along a mass-flowering crop gradient. Mol. Ecol. 2020, 29, 5003–5018. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microscopy | DNA Metabarcoding | ||||||
|---|---|---|---|---|---|---|---|
| Taxonomic Level | Comparison Level | F | df | p | F | df | p |
| Family | Site | 1.57 | 14; 39 | 0.13 | 2.66 | 14; 39 | 0.008 |
| Landscape category | 2.05 | 2; 51 | 0.14 | 0.96 | 2; 51 | 0.39 | |
| Genera | Site | – | – | – | 1.04 | 2; 51 | 0.36 |
| Landscape category | |||||||
| Wald. Stat. | df | p | |||||
| Species | Site | 0.19 | 14; 427 | 0.65 | – | – | – |
| Landscape category | 0.06 | 14; 427 | 0.80 | – | – | – | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bontšutšnaja, A.; Karise, R.; Mänd, M.; Smagghe, G. Bumble Bee Foraged Pollen Analyses in Spring Time in Southern Estonia Shows Abundant Food Sources. Insects 2021, 12, 922. https://doi.org/10.3390/insects12100922
Bontšutšnaja A, Karise R, Mänd M, Smagghe G. Bumble Bee Foraged Pollen Analyses in Spring Time in Southern Estonia Shows Abundant Food Sources. Insects. 2021; 12(10):922. https://doi.org/10.3390/insects12100922
Chicago/Turabian StyleBontšutšnaja, Anna, Reet Karise, Marika Mänd, and Guy Smagghe. 2021. "Bumble Bee Foraged Pollen Analyses in Spring Time in Southern Estonia Shows Abundant Food Sources" Insects 12, no. 10: 922. https://doi.org/10.3390/insects12100922
APA StyleBontšutšnaja, A., Karise, R., Mänd, M., & Smagghe, G. (2021). Bumble Bee Foraged Pollen Analyses in Spring Time in Southern Estonia Shows Abundant Food Sources. Insects, 12(10), 922. https://doi.org/10.3390/insects12100922