1. Introduction
Commercial orchards are among the most intensively sprayed crops. However, pest control using chemicals is environmentally costly and becomes increasingly difficult due the development of resistance to the most commonly used pesticides [
1]. Production sustainability should thus be enhanced by using alternative methods to the use of pesticides [
2,
3]. Among them, biological control tends to reduce pest pressure by using various techniques, such as the fostering of natural enemies (predators, parasitoids), spraying with natural pesticides, and establishing wild strips or companion plants. Other methods focus more on the plants, attempting to breed resistant cultivars or adapting current cultural practices (e.g., fertilization, irrigation or pruning) to reduce plant susceptibility to pests, by fostering plant traits which are unfavourable to pests (e.g., production of defence compounds, plant shaping to limit pest sheltering and dissemination within the plant, reduced access to food resources, among others). The effectiveness of cultural practices relies on the existence of cross-tolerance between biotic and abiotic stresses [
4], and on the dependence of pest and plant development [
5]. This aspect has however only been little investigated, and prior to its implementation in commercial orchards, the main determinants of plant susceptibility have to be identified, and their variations together with growing conditions, genotype, and tree life background characterized.
Of high economic importance, the peach tree–aphid system has been one of the most studied in recent years. Thus, peach (
Prunus persica L. Basch) ranks first in stone fruit production, both worldwide (25.7 million tons) [
6] and in the Mediterranean basin (3.8 million tons) [
6]. Of the numerous herbivore pests, the green peach aphid (
Myzus persicae Sulzer) represents a major threat that affects fruit production for several years and transmits viruses. Peach susceptibility to green peach aphid has been found to be positively related to shoot development (emergence of new organs) [
7,
8] and growth (increase in organ size) [
5], as well as to apex concentrations [
8,
9,
10,
11] of total N (up to a threshold level), amino acids and non-structural sugars. The polyphenol concentrations, in contrast, were negatively related to tree susceptibility [
8,
12]. Tree susceptibility is thus determined by developmental and trophic aspects, i.e., by the balance between plant or shoot development, growth, and composition. Growing conditions and tree life background affect each of these aspects differently, with possible antagonistic effects on the defence capability. To avoid producing contradictory results, our understanding of plant pest interactions has to therefore consider simultaneously all these aspects.
For practical and feasibility reasons, studies on integrated plant functioning have mostly been performed on small, young, potted trees grown under controlled, or semi-controlled, conditions. However, aging and orchard conditions affect different aspects of plant metabolism. So, on adult trees, fruits represent a principal trophic sink in competition with vegetative development [
13,
14] when aphid infestation could arise. The relative size of the nitrogen (N) and carbon (C) reservoirs (i.e., the proportion of the roots and the proportion of one-year-old wood), increase also with plant age, meaning that store mobilization could buffer limited supplies over long periods (i.e., several years) [
15]. This could, for instance, explain why defoliation or foliar damage, whatever their cause, are less harmful on adult than on young trees. More generally, tree growth modifies the balance between shoot or tree size, shape, and composition. However, not all determinants of plant susceptibility are affected similarly, thereby modifying their effects on plant-pest interactions.
The results obtained under semi-controlled conditions must therefore be adapted to productive trees grown in open fields. The present study focuses on the peach tree—aphid system. The optimal values (e.g., rates, dimensions, concentrations) of the key plant variables determined in previous studies [
3,
5,
8] will be re-evaluated under orchard conditions, and their final impact on aphid infestations analysed, considering the functional dependence between organ formation, growth, and composition. To reach this goal, an experiment was carried out under contrasting production conditions, in an organic commercial orchard regularly subject to severe aphid infestations. Reference trees were selected from two genotypes of different precocity and vigour established on adjacent plots, and then received different levels of water and nitrogen supplies. The treatments were designed to be consistent with regional agricultural practice. They were repeated over two consecutive years, during which we were reliant on natural aphid infestations, artificial ones not being feasible in commercial orchards. This represented a major constraint, which led us to consider not only the green peach aphid but also to include the two additional species present, (i) the leaf-curl plum aphid (
Myzus varians Davidson) and (ii) the mealy plum aphid (
Hyalopterus pruni Geoffer). Thus, we assumed that the determinisms of plant susceptibility to green peach aphids and to both additional species were mostly similar, since they developed at the same times and on the same organs as the green peach aphid, sometimes in mixed colonies. Another assumption behind this work is that the external environmental aphid pressure was similar for all selected trees, meaning that aphids settled on the ones best able to foster their development.
This study aims first to discuss the effects of treatment, year, and variety on severity of infestation, defined as the proportion of infested shoots, and then to explain these effects, comparing the overall condition of infested and non-infested shoots. Indeed, aphid dispersion within the canopy is related to the number of shoots whose functional balance enables the establishment of new colonies, which may vary with cultural practices, genotype, or pedo-climate. Infested and non-infested (i.e., control) shoots were therefore compared for their growth, analysed from an architectural point of view, as well as their composition in non-structural carbohydrates (NSC), total N, amino acids, and polyphenol content. At shoot level, this extensive screening allowed (i) the defining of conditions which favoured (or prevented) aphid infestation, (ii) the determining to what extent these conditions depended on cultural practices (irrigation and fertilization), genotype or year, and (iii) the identifying of differences from those previously established in controlled or semi-controlled conditions. At tree level, this study also gives insight into how infestation severity may be manipulated by routine cultural practices.
2. Materials and Methods
2.1. Experimental Design
The study was carried out over two consecutive years (2018 and 2019), in a 1.5 ha organic commercial orchard located near Avignon (Southeast France, Lat: 43.89°, Long: 4.77°), established in 2008, at a density of 555 trees/ha (6 m and 3 m distances between rows and between trees, respectively), trained in a “gobelet” or open-centre shape. The rows, oriented E-W, were protected from dominant north winds by cypress hedges. The orchard is composed of two varieties of peach trees, both susceptible to aphids: Ivoire (vigorous, early ripening white peach) and Conquise (less vigorous, middle early ripening, white peach), grafted onto Cadaman-Avimag. This rootstock is adapted to the deep, clay-silty, and occasionally (especially in winter) anoxic soil. The aphicide treatment consisted of a single spray of mineral oil during winter. It was, similar to all cultural operations (pruning, thinning, harvesting, interrow mowing, prophylactic treatments), coherent with regional standards, prescribed (date, dose or intensity) and achieved by the orchard owner. The experimental treatments were applied to preselected trees. This treatment consisted of three modalities: the water and N inputs were either (i) unrestricted, i.e., those applied by the orchard owner (control or HN treatment), (ii) restricted only in water (hN treatment), or (iii) restricted in both water and N (hn treatment). The water supply was limited after commencement of irrigation (June 19 in 2018, June 1 in 2019) by reducing (30 L h
−1 vs. 45 L h
−1) the flow rate of the irrigation drippers. For N, both the 2018 and 2019 inputs were reduced. As a result, each HN tree received, from irrigation start to July 25 (i.e., until the end of harvest), 22 L of water in 2018 and 23 L in 2019, which had to be added to the rainfall (86 and 48 mm between April 1 and July 25 in 2018 and 2019, respectively). The N supplies were of 84 and 83 g N in 2018 and 2019, respectively. For the restricted treatments, the inputs were reduced by 30% as detailed in
Table 1.
The 54 selected trees were distributed in 18 groups (nine per variety) of three adjacent individuals, chosen for their homogeneity in terms of vigour, i.e., trunk diameter (53 cm ± 1.2 SE—standard error—for Ivoire, 58 cm ± 0.9 for Conquise) and number of branches 113 ± 4 for Ivoire and 115 9 ± 4 for Conquise. The 18 groups were located along six rows (i.e., three rows per variety, three groups per row). Each treatment modality was applied to three groups (i.e., nine trees) per variety: one per row, and the position of each modality along the row was varied depending on the row (e.g., from the nearest tree group on the first row, to the median group on the second row, and then to the furthest one on the third row). Each group received the same treatment in 2018 and 2019.
2.2. In Situ Measurements
The air temperature, rainfall and soil humidity were recorded every 15 min with an automated weather station and four capacitive sensors positioned at 40 cm soil depth, to regulate the irrigation and to record any possible extreme climatic events (frosts, etc.).
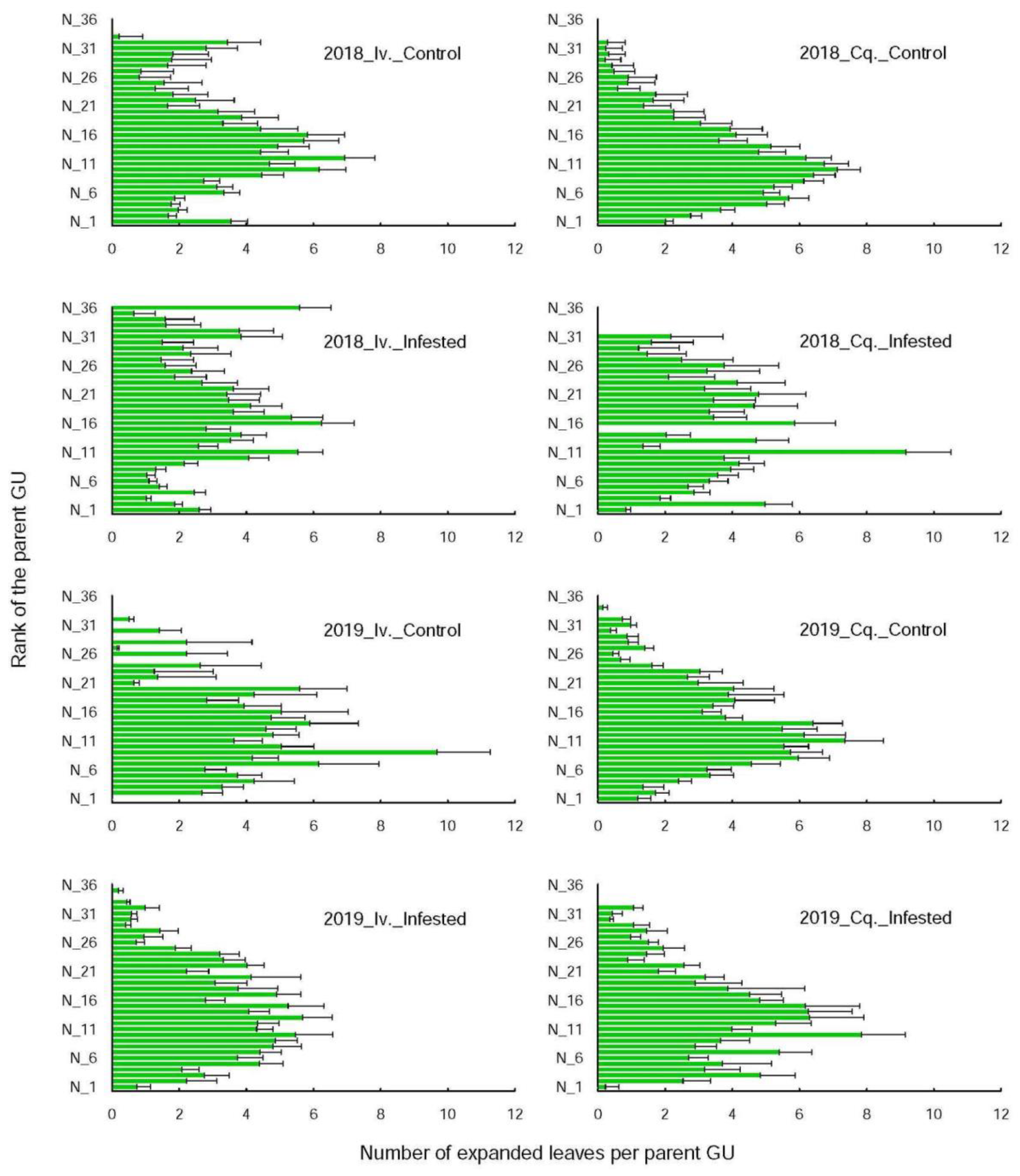
For each tree, we selected five (2018) and seven (2019) unbranched and one-year-old shoots at infestation start (i.e., before 3 May in 2018 and 9 May in 2019). Two of them were infested when selected, and the remaining (three in 2018 and five in 2019) were aphid-free controls of similar size and shape to the infested ones. Their growth was then monitored on an architectural basis, each shoot being considered as a collection of growth units (or GU) comprising an internode and its upper node with the attached leaf and axillary bud [
16]. Each axillary bud present on the selected shoots can, or cannot, give rise to a long daughter axis. The number of their expanded leaves corresponds to the number of their fully developed GUs. They were counted on the selected shoot stems and on each of their long daughter axes, previously positioned along the parent shoots by the rank of their father’s GU. Therefore, the number of leaves on a selected shoot GU was correlated to the length of the daughter to which it gave rise, if any, and was equal to one (the shoot father’s GU leaf) in absence of ramification. The repartition of the leaves along the shoot stem was thus representative of the ramification process (in terms of numbers, positions, and lengths). The measurements were taken at shoot selection, and then after fruit harvest during the period of shoot growth rest (see discussion), i.e., on 25 July 2018, and 11 July 2019.
2.3. Aphid Pressure in the Orchard
The same observer monitored the aphid infestation fortnightly in 2018 (i.e., between 30 April and 3 July) and weekly in 2019 (i.e., between 7 May and 30 June). The aphid species present on each selected tree and shoot was identified and the infestation status evaluated at tree and shoot level. More precisely, a class was assigned to each tree as follows: S0 (no aphid), S1 (less than 15% infested shoots), S2 (between 15 and 50%) and S3 (more than 50%). The shoots were classified as infested or controls according to the presence or absence of reproducing aphids. Indeed, single adults could be found on a shoot on one date but did not settle, i.e., were no longer present at the following count, thus indicating that aphids choose their hosting shoots by roaming within the canopy until they find a suitable site. The control shoots which became infested during the observation period were excluded from the statistical analyses.
Most trees were infested by more than one species. The dominant one was designated considering its abundance and harmfulness (green peach aphid > mealy plum aphid > leaf curl plum aphid). More precisely, a species was dominant at tree level if (i) it was more harmful and abundant, or (ii) it was more harmful and less abundant, but with a difference of only one class (e.g., S2 instead of S3, or S1 instead of S2). This rule was established following discussion with the orchard owner, considering his perception of the damage and economic losses due to aphid infestation. Different species could also be observed on the same shoot, all of them being passive phloem feeders which select their hosting shoot according to the same criteria.
2.4. Biochemical Determinations
The shoots were sampled on 16 May 2018, and 13 June 2019, i.e., during the period of rapid increase in the populations of the dominant aphid species. As far as possible, two samples were collected per tree, each of them ideally being composed of four to six growing apices (comprising the terminal meristems and the smallest unfolded leaves) of just emerging daughter axes inserted either on infested or on control shoots. It should be noted that on infested shoots, it was sometimes difficult to find apices young enough not to be colonized. The samples were immediately immersed in liquid N, and then stored at −80 °C until freeze-dried and ground in ball mills (MM301, Retsch, Germany) cooled with liquid N. The biochemical determinations were performed as described precisely in Jordan et al. [
8]: (i) the total N using an elemental analyser (Flash EA 1112, Thermo Finnigan, Milan, Italy), (ii) starch and soluble sugars using an enzymatic method, (iii) amino acids by HPLC (Cortecs C18 column 4.6 mm × 150 mm, particle size 2.7 μm; Waters) in line with a 2475 multiwavelength fluorescence detector, and (iv) the polyphenols by HPLC (Uptisphere HDO column; Interchim, Montluçon, France) in line with a DAD UV–visible detector (Surveyor, Thermo Fisher Scientific, River Oaks Pkwy, SJ, USA). For the polyphenols, the measurements were made at two wavelengths (280 or 330 nm) and eight compounds (listed in
Table 2) were identified by their standards. Four unidentified significant polyphenols peaks, resembling hydroxycinnamate derivatives, were also quantified using the 5CQA (
Table 2) calibration curve.
2.5. Data Analysis
The statistical analyses were focused on shoot level and were performed using R software (R Core Team 2018, Vienna, Austria) [
17].
Shoot growth was approximated by the leaf expansion rate (i.e., the increase in the number of fully developed GUs on the daughter axes and the selected shoot terminal bud). This rate was calculated for each shoot and corresponded to the daily increase in the number of expanded leaves during the observation period, divided by the length of the selected shoot stem (in meters). The C pools were quantified by considering either only the soluble sugars (glucose, sucrose, fructose, and sorbitol), i.e., the immediately available and transportable C forms, and the NSC pool, which includes also starch, i.e., the long-term storage form. The amino acids and polyphenols were summed to build the polyphenol and amino acid pools, but since each compound plays a specific role in plant metabolism, they were also considered separately in the analyses.
The leaf expansion rates, and the apex compositions were compared using non-parametric permutation tests performed at a 5% level, for which the empirical distributions of each variable effect were derived from 2500 random assignments of the tested dataset. The impact of treatments on each subgroup of shoots of similar infestation status, variety, and year was evaluated first. The absence of effect allowed the merging of shoots whatever their treatment, and to obtain pairwise comparisons between these newly formed groups, to assess, depending on the comparison, the effects of infestation status, year, or variety.
Kolmogorov–Smirnov tests were used following the previously described protocol, to evaluate how far the shoots differed: (i) in the distribution of their expanded leaves (i.e., of their long daughter axes) along the selected shoot stems, and (ii) in their apex’s amino acid, soluble sugar, and phenol profiles, i.e., in the respective contribution of each component to the cited pools.
3. Results
3.1. Aphid Pressure at Tree Level Varies More with Genotype and Year Than with Treatment
Aphid pressure in the orchard was assessed, evaluating for each selected tree, the dominant aphid species, and its abundance at infestation peak, i.e., the highest infestation severity recorded throughout the observation period (
Table 3). These effects were lower in the trees of Conquise than of Ivoire for all treatments and years.
The effects of treatment varied over time. In 2018, the hn treatment, limited in water and N, comprised the lowest number of trees infested by the most harmful species, i.e., by the green peach aphid (5 and 4 for Ivoire and Conquise, respectively, vs. 8 and 5 for the HN treatment), and the highest number of aphid-free trees for both varieties (3 and 2 for Ivoire and Conquise, respectively, vs. 0 and 1 for the HN treatment). The aphid abundances were also the lowest on these trees. The differences in aphid species and abundances between the HN and hN treatments were small but the hN trees seemed to be slightly more susceptible to aphids for both varieties. In 2019, Conquise followed almost the same pattern as in 2018, since the leaf-curl plum aphid, i.e., the less harmful species, was overrepresented on the hN and hn trees (6 and 7 trees, respectively), subjected to deficit irrigation, compared to the HN ones (3 trees). This was especially true for the hn trees, which also had the smallest proportion of infested shoots (since all trees had fewer than 15% infested shoots vs. 7 trees for the HN treatment). It was again difficult to rank the two well fertilized treatments, since two of the nine HN trees were aphid-free and two others hosted the green peach aphid, vs. 0/9 and 1/9 hN trees, respectively. For Ivoire, one in three HN and hn trees were infested by leaf curl plum aphid. In contrast, the green peach aphid was overrepresented in the hN trees (7/9 trees), showing this treatment to be the most susceptible one.
Whatever the treatment and variety, the infestations were less detrimental in 2019 than in 2018. Indeed, the trees were more tolerant to the species which developed in the second year in almost all situations. This higher tolerance compensated widely for the slightly greater aphid abundances observed in 2019.
In short, the hN treatment was suspected to be the most favourable to aphids, but this assertion was evident only for Ivoire_2019. In contrast, the hn treatment was the less susceptible in all cases, except for Ivoire_2019. The effectiveness of treatment was however small compared to those of variety and year.
3.2. The Interactions between Shoot Growth and Infestation Depend on Year
Treatment and tree infestation severity affected neither shoot growth rates (i.e., the daily increase in the number of expanded leaves per shoot including those inserted on the long daughter axes), nor shoot shape (i.e., the repartition of those leaves along the shoot stem that depend on the number, positions, and lengths of the daughter axes). The growth rates (
Table 4) on the infested and control shoots were however significantly (i.e., between 1.5 and 5.5 times) lower (i) in 2019 than in 2018 for both varieties. Conquise, known to be less vigorous, also exhibited growth rates whose values were lower by 25 to 65% than those of Ivoire. In contrast, the shoot shapes (
Figure 1) were affected neither by variety, nor by year with one exception. For Ivoire, the leaves were more numerous on the upper growth units on the infested shoots in 2018 than in 2019. That meant that the daughter axes on those Ivoire infested shoots were not only more numerous (1.07 ± 0.3 vs. 0.96 ± 0.06) but also longer (18.5 ± 3.3 vs. 15.3 ± 1.1 leaves) in 2018 than in 2019.
The interactions between aphid development, shoot vigour and shape were year dependent. In 2018, the infested and control shoots had similar growth rates (
Table 3), but the infested shoots bore more leaves on their upper growth units than the control ones. The difference was however significant only for Conquise, since the proportion of leaves inserted on the parent axes upper third were, respectively 14% and 4% for the infested and control shoots. In 2019, by contrast, the shoot growth rates were 1.7 (Ivoire) and 3 (Conquise) times higher for the infested shoots than for the controls, without any effect on shoot shape. The newly formed leaves (i.e., the leaves which expanded on the daughter axes during the aphid infestation period) were thus similarly distributed along the control and the infested shoots, despite being more numerous on the infested shoots.
3.3. The Relation between Infestation and Apex Composition Varied with Variety and Year
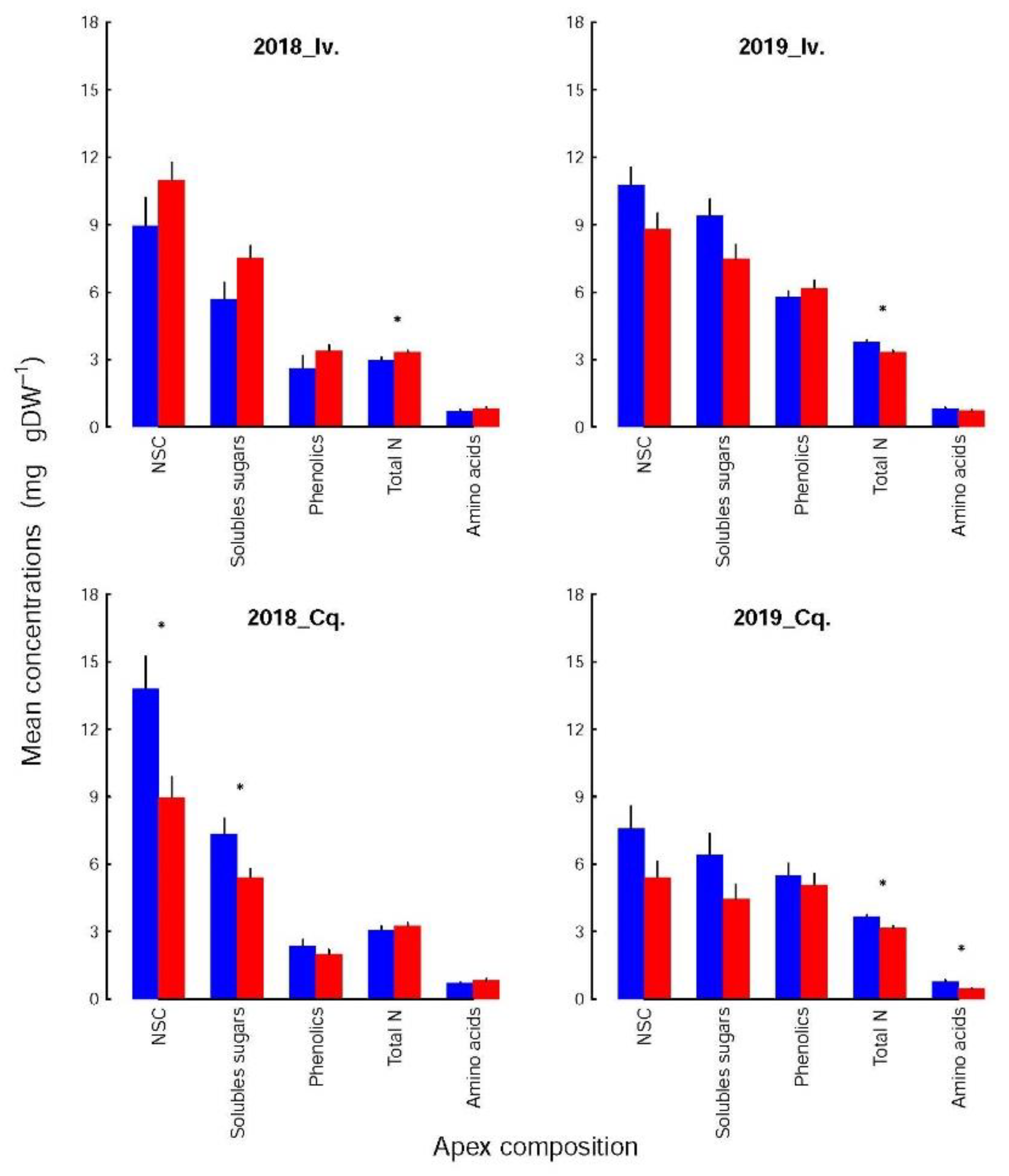
Each biochemical family (
Figure 2) was not affected to the same extent by variety and shoot infestation status. For the soluble sugars and NSC, the concentrations were higher in the control than in the infested shoots, except for Ivoire_2018. The differences were significant only for Conquise_2018 (13.8 ± 1.4 vs. 8.9 ± 0.9% DW for the NSC and 7.3 ± 0.71 vs. 5.4 ± 0.40%DW for the soluble sugars). The specific behaviour of Ivoire_2018 was related to the low concentration of soluble sugars in the controls (5.7% DW vs. more than 7.5% DW for the other groups of Ivoire shoots). These control shoots of Ivoire_2018 were subsequently the only group whose soluble sugar concentrations (i) were not significantly higher for Ivoire than for Conquise and (ii) significantly lower in 2018 than in 2019. The concentrations of soluble sugars (SolS
conc) were also proportional to those of NSC (NSC
conc), and the relations between both compounds (Equations (1) and (2), with R
2adj significance level: *** = 0.001) were affected neither by variety nor infestation status, but by year. These interannual fluctuations were due to a dramatic decrease (by 60% at least) in starch concentrations in 2019 compared to 2018.
For amino acids and total N, the concentrations (AA
conc and N
conc) were lower in the infested shoots than in the controls in 2018, and higher in 2019. For total N, they varied between 3.16 and 3.32% DW in the infested shoots, whatever the variety and year. The concentrations in the control shoots were beyond this range in 2018, and above it in 2019. Significant differences could thus be evidenced (i) between the infested and control shoots except for Conquise_2018, (ii) between years for the control shoots but not for the infested ones, but (iii) not between varieties. The amino acids followed the same patterns, since they are linked to the total N by the following polynomial relation (Equation (3) with R
2adj significance level: ** = 0.01).
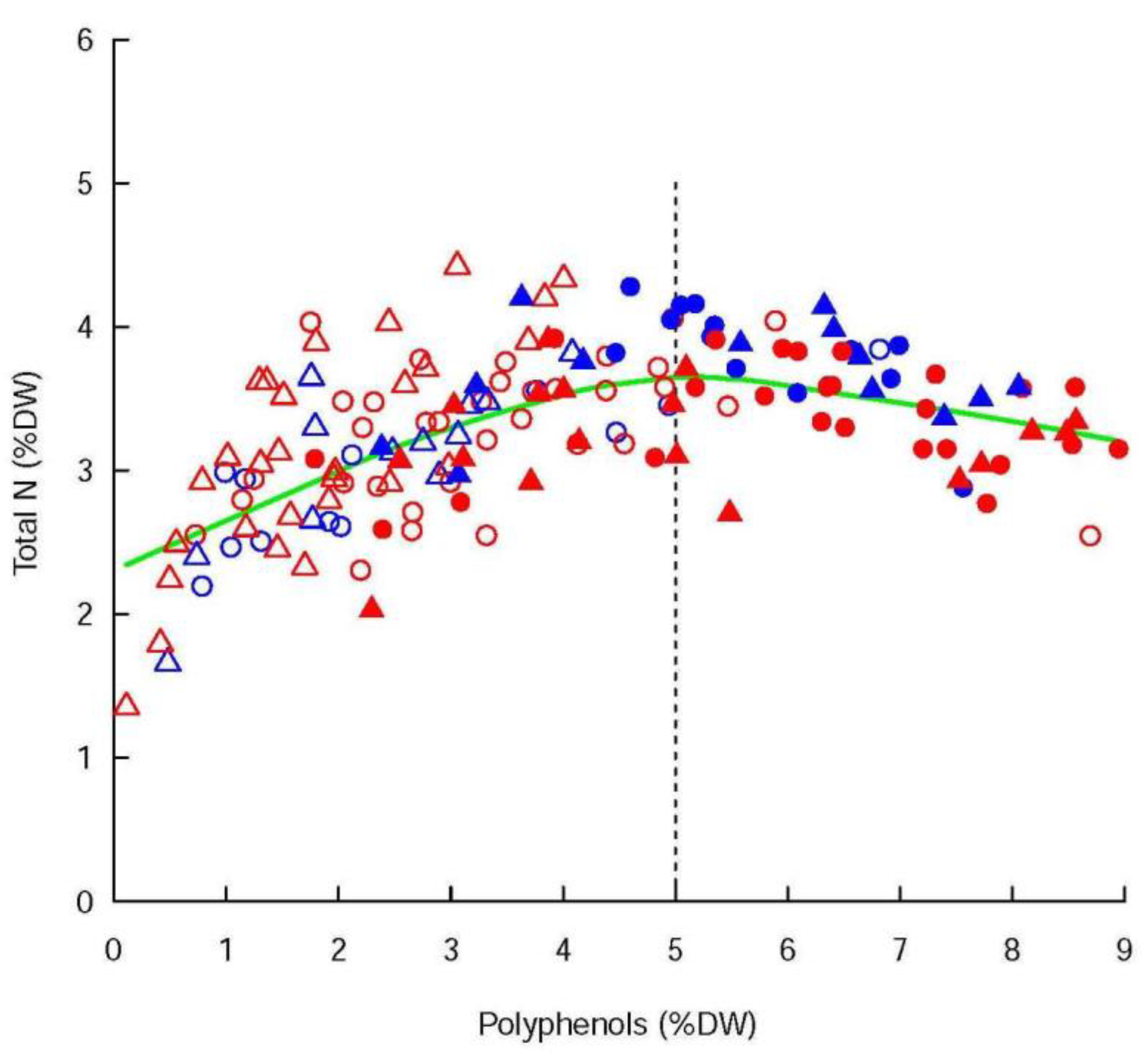
The polyphenols were not significantly affected by the infestation status, even if the concentrations were higher in the infested shoots than in the controls for Ivoire, and lower for Conquise. The difference among varieties were thus limited to the infested shoot. Indeed, in 2018 the concentrations of the infested shoots were of 3.38 ± 0.26% DW and 2.0 ± 0.20% DW for Ivoire and Conquise, respectively. In 2019 those values jumped to 6.16 ± 0.36% DW and 5.08 ± 0.49% DW, respectively. The greatest variation range was observed between years, since the concentrations, at least doubled between 2018 and 2019. Furthermore, the concentrations of polyphenols (Phe
conc) and total N were linked (
Figure 3): Phe
conc increased along with N
conc until a threshold corresponding to Phe
conc = 5% DW, then decreased slightly. Most of the values were below this threshold in 2018 and above it in 2019, and both families were consequently related positively in 2018 but negatively in 2019.
3.4. The Composition of the Different Families Differed Only by a Few Components
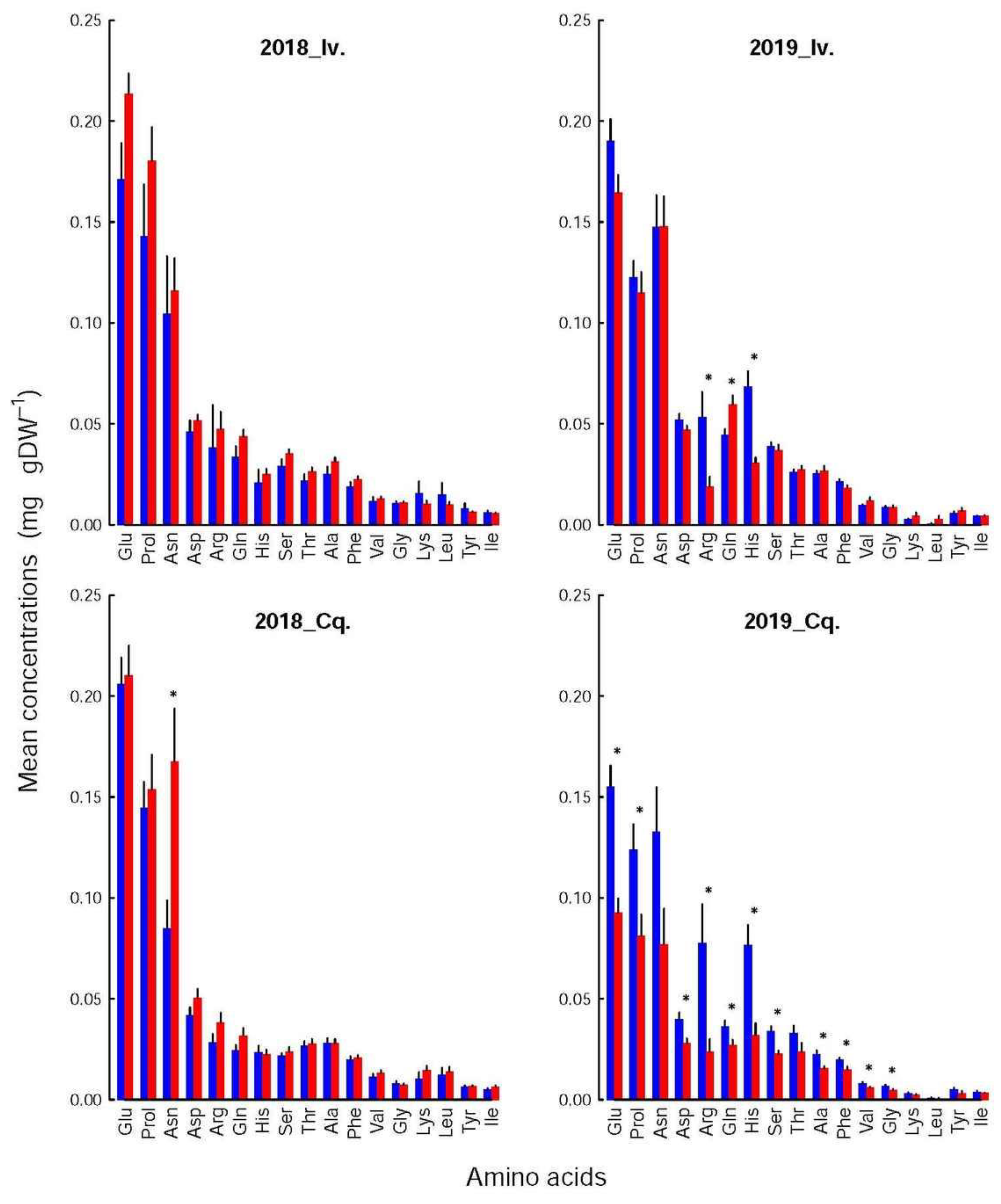
The composition (i.e., the relative contribution of each constituent) of the amino acids and the polyphenol pools were not altered by the shoot infestation status, year, or variety (Kolmogorov-Smirnoff tests), although the concentrations of several of their constituents could vary significantly. These variations were, however, too small to significantly affect the pool compositions. For instance, the relative contributions of each amino acid or polyphenol (
Figure 4 and
Figure 5) to its respective pool varied at most by 5% with shoot infestation status. The mean variation was 1.23 ± 0.12% but reached 2.08 ± 0.33% when the calculations were restricted to the compounds whose concentrations differed significantly between both groups. The greatest difference was observed for the NSC, more precisely for starch in Conquise_2018, whose contribution to the pool varied by 12%. That again did not affect the composition, but solely the concentrations of the NSC pool (
Figure 3).
The differences between infested and control shoots depended more on variety and year. Thus, among the amino acids (
Figure 4), only one compound was significantly affected in 2018 (Asn for Conquise), while in 2019, their number increased to three for Ivoire (His, Gln and Arg) and to 11 for Conquise (all acids but Asn, Thr, lys, Leu, Tyr, Ile). Concerning the polyphenols (
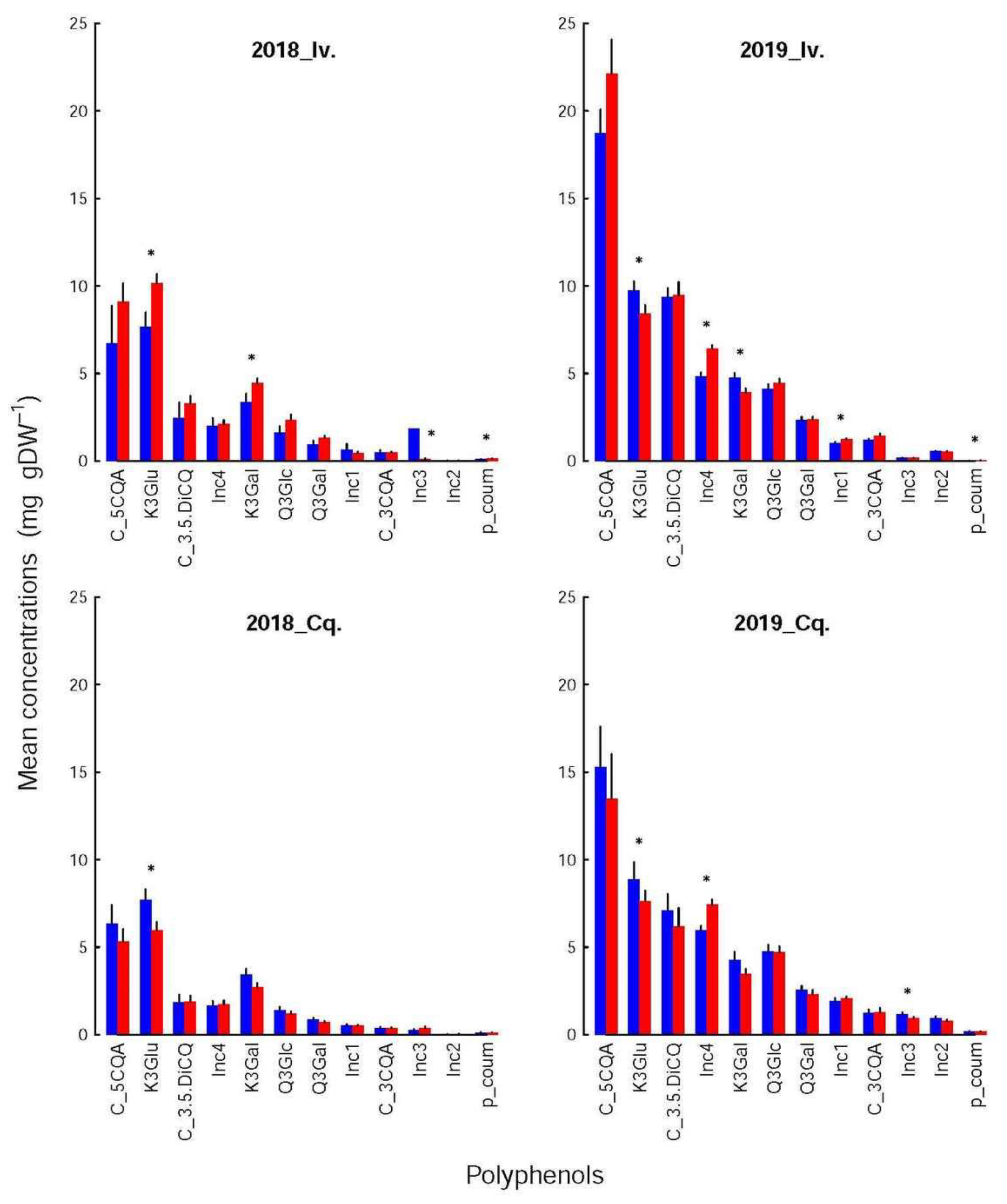
Figure 5), the differences could either be found in all shoot groups (K3Glu), be restricted to Ivoire (p-coum), or be erratic (Inc3 and K3Gal for Ivoire_2018, Inc1 and Inc2 for Ivoire_2019, and Inc3 and Inc4 for Conquise_2019).
If dependent on variety (
Table 4), the concentrations were higher for Ivoire than for Conquise for all compounds, but only some polyphenols. These exceptions were more restricted to the infested shoots and concerned only five compounds: p-coum, Inc1, Inc2, Inc3 and Inc4. All of them except Inc4 were minor compounds, i.e., contributed only little to the pool. All variations (increase or decrease) were in most cases limited to around 25%. They were around 40% for a few amino acids and reached 80% for two minor compounds: Leu and Inc3.
Furthermore, the concentrations of all the polyphenols, except for K3Gal, K3Glu and p-coum, were higher in 2019 than in 2018, thus explaining the huge interannual variation of the pool. Indeed, the concentrations increased by a factor of at least two to three for almost all compounds with two exceptions. For K3Glu and 3Gal they were limited to around 20%. The situation was more varied for the amino acids. The concentrations were higher by around 25% in 2019 than in 2018 for Ser in the control shoots for both varieties, Gln and Arg in the control shoots of Conquise, and Gln in the infested shoots of Ivoire. For His, the increase in the control shoots was higher: they doubled. In all other situations, except for Lys and Leu, the concentrations remained stable and decreased by at least 20% over the years. This decrease was higher for Lys and Leu (two minor acids): it was more important, being around 90%. The inter-annual difference therefore was small, and significant only for the infested shoots of Conquise.
5. Conclusions
Our results confirmed that lowering simultaneously the water and N inputs reduces the severity of aphid infestation in commercial orchards. These practices could reasonably be implemented in commercial orchards since they do not alter fruit production: 150 and 121 fruits per tree in 2018 and 2019, respectively, with no effects of treatment on fruit size, i.e., on the commercial value (undetailed data). Our results are also consistent with previous ones obtained in young potted trees [
3,
8] in two aspects: whatever the culture conditions, aphid abundance increases with N supply and are lowest on the trees which are limited in both N and water. The N concentrations in the apices of the infested shoots are also similar in potted and orchard trees (i.e., around 3.2%), thus indicating this variable as a major determinant of shoot susceptibility. Adult and young potted trees differ greatly nonetheless when considering the effects of treatments and infestation status on shoot development and apex concentrations in polyphenols and NSC. The differences are related to the mean values and their variations with the shoot infestation status, being in most cases higher and more significant in young potted trees.
In orchards, the stress intensities during the infestation period are moderate, being buffered, firstly, by the soil water content and pluviometry which contribute greatly and in an unpredictable way to water supply, and secondly, by the high N inputs usually found at bud burst. The decrease in infestation severity induced by a limited and transient trophic stress could however not be sufficient, meaning that other alternatives have to be implemented at the same time to control efficiently aphid infestations. Among them, pruning, which could reduce the number of susceptible (vigorous) shoots by increasing their number, or changes in ground cover, have to be taken into consideration.
Our results were obtained from, and are therefore specific to, our localized conditions. To ensure their robustness and generality, they will now be confirmed across a wider range of genotypes and climatic conditions, by regular screenings performed over a large geographical area, which will probably be more informative than continuing our experiment beyond two years.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}