
Genus-Wide Characterization of Nuclear Mitochondrial DNAs in Bumblebee (Hymenoptera: Apidae) Genomes


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Genus-Wide Mitogenome Resources for Bumblebees
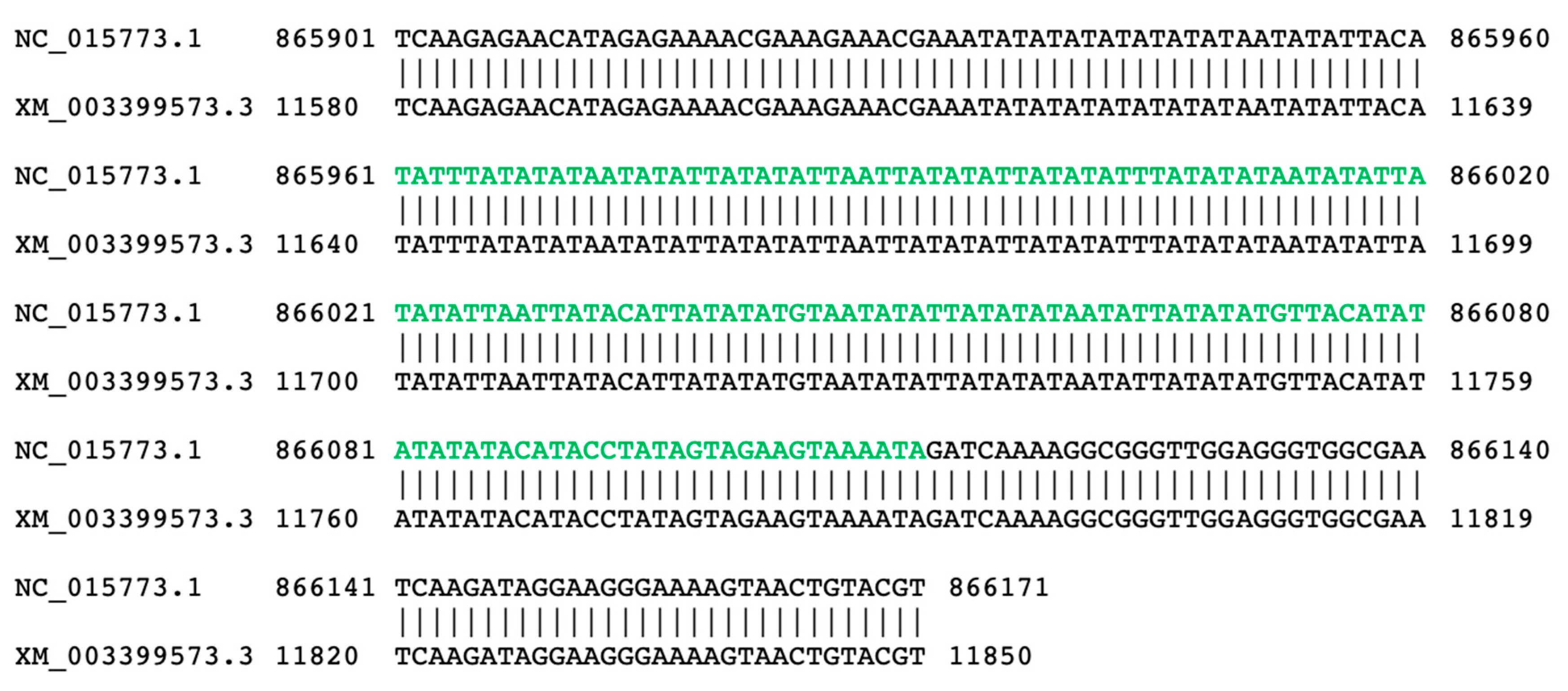
3.2. Molecular Features of NUMTs in Bumblebee Nuclear Genomes
3.3. Contribution of NUMTs to Bumblebee Genome Innovation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Timmis, J.N.; Ayliffe, M.A.; Huang, C.Y.; Martin, W.F. Endosymbiotic gene transfer: Organelle genomes forge eukaryotic chromosomes. Nat. Rev. Genet. 2004, 5, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Kleine, T.; Maier, U.G.; Leister, D. DNA Transfer from Organelles to the Nucleus: The Idiosyncratic Genetics of Endosymbiosis. Annu. Rev. Plant Biol. 2009, 60, 115–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazkani-Covo, E.; Zeller, R.M.; Martin, W. Molecular Poltergeists: Mitochondrial DNA Copies (numts) in Sequenced Nuclear Genomes. PLoS Genet. 2010, 6, e1000834. [Google Scholar] [CrossRef]
- Ricchetti, M.; Fairhead, C.; Dujon, B. Mitochondrial DNA repairs double-strand breaks in yeast chromosomes. Nat. Cell Biol. 1999, 402, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Richly, E.; Leister, D. NUMTs in sequenced eukaryotic genomes. Mol. Biol. Evol. 2004, 21, 1081–1084. [Google Scholar] [CrossRef] [Green Version]
- Noutsos, C.; Richly, E.; Leister, D. Generation and evolutionary fate of insertions of organelle DNA in the nuclear genomes of flowering plants. Genome Res. 2005, 15, 616–628. [Google Scholar] [CrossRef] [Green Version]
- Hazkani-Covo, E.; Covo, S. Numt-Mediated Double-Strand Break Repair Mitigates Deletions during Primate Genome Evolution. PLoS Genet. 2008, 4, e1000237. [Google Scholar] [CrossRef] [Green Version]
- Michalovova, M.; Vyskot, B.; Kejnovsky, E. Analysis of plastid and mitochondrial DNA insertions in the nucleus (NUPTs and NUMTs) of six plant species: Size, relative age and chromosomal localization. Heredity 2013, 111, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Turner, C.; Killoran, C.; Thomas, N.S.T.; Rosenberg, M.; Chuzhanova, N.A.; Johnston, J.; Kemel, Y.; Cooper, D.N.; Biesecker, L.G. Human genetic disease caused by de novo mitochondrial-nuclear DNA transfer. Qual. Life Res. 2003, 112, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Leister, D. Origin, evolution and genetic effects of nuclear insertions of organelle DNA. Trends Genet. 2005, 21, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Noutsos, C.; Kleine, T.; Armbruster, U.; DalCorso, G.; Leister, D. Nuclear insertions of organellar DNA can create novel patches of functional exon sequences. Trends Genet. 2007, 23, 597–601. [Google Scholar] [CrossRef]
- Zhang, D.-X.; Hewitt, G.M. Nuclear integrations: Challenges for mitochondrial DNA markers. Trends Ecol. Evol. 1996, 11, 247–251. [Google Scholar] [CrossRef]
- Williams, P. An Annotated Checklist of Bumble Bees with an Analysis of Patterns of Description (Hymenoptera: Apidae, Bombini). Bull. Nat. Hist. Mus. Entomol. 1998, 67, 79–152. [Google Scholar]
- Cameron, S.A.; Hines, H.M.; Williams, P.H. A comprehensive phylogeny of the bumble bees (Bombus). Biol. J. Linn. Soc. 2007, 91, 161–188. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.H.; Cameron, S.A.; Hines, H.M.; Cederberg, B.; Rasmont, P. A simplified subgeneric classification of the bumblebees (genus Bombus). Apidologie 2008, 39, 46–74. [Google Scholar] [CrossRef] [Green Version]
- Velthuis, H.; Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Lye, G.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Èntomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Ricardo, P.C.; Françoso, E.; Arias, M.C. Mitochondrial DNA intra-individual variation in a bumblebee species: A challenge for evolutionary studies and molecular identification. Mitochondrion 2020, 53, 243–254. [Google Scholar] [CrossRef]
- Sun, C.; Huang, J.; Wang, Y.; Zhao, X.; Su, L.; Thomas, G.W.C.; Zhao, M.; Zhang, X.; Jungreis, I.; Kellis, M.; et al. Genus-Wide Characterization of Bumblebee Genomes Provides Insights into Their Evolution and Variation in Ecological and Behavioral Traits. Mol. Biol. Evol. 2021, 38, 486–501. [Google Scholar] [CrossRef]
- Sadd, B.M.; Barribeau, S.M.; Bloch, G.; De Graaf, D.C.; Dearden, P.; Elsik, C.G.; Gadau, J.; Grimmelikhuijzen, C.J.P.; Hasselmann, M.; Lozier, J.D.; et al. The genomes of two key bumblebee species with primitive eusocial organization. Genome Biol. 2015, 16, 76. [Google Scholar] [CrossRef] [Green Version]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [PubMed] [Green Version]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; Depamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Behura, S.K. Analysis of Nuclear Copies of Mitochondrial Sequences in Honeybee (Apis mellifera) Genome. Mol. Biol. Evol. 2007, 24, 1492–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Cha, S.Y.; Yoon, H.J.; Lee, E.M.; Yoon, M.H.; Hwang, J.S.; Jin, B.R.; Han, Y.S.; Kim, I. The complete nucleotide sequence and gene organization of the mitochondrial genome of the bumblebee, Bombus ignitus (Hymenoptera: Apidae). Gene 2007, 392, 206–220. [Google Scholar] [CrossRef]
- Du, Q.; Bi, G.; Zhao, E.; Yang, J.; Zhang, Z.; Liu, G. Complete mitochondrial genome of Bombus terrestris (Hymenoptera: Apidae). Mitochondrial DNA Part A 2015, 27, 4455–4456. [Google Scholar] [CrossRef]
- Zhao, X.; Huang, J.; Sun, C.; An, J. Complete mitochondrial genome of Bombus consobrinus (Hymenoptera: Apidae). Mitochondrial DNA Part B 2017, 2, 770–772. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, Z.; Huang, J.; Liang, C.; An, J.; Sun, C. Complete mitochondrial genome of Bombus breviceps (Hymenoptera: Apidae). Mitochondrial DNA Part B 2017, 2, 604–606. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Yan, J.; Jiang, K.; Huang, Z.; Lin, G. Nearly complete mitochondrial genomes of four bumblebee species (Hymenoptera: Apidae: Bombus). Mitochondrial DNA Part B 2019, 4, 183–184. [Google Scholar] [CrossRef] [Green Version]
- Pamilo, P.; Viljakainen, L.; Vihavainen, A. Exceptionally High Density of NUMTs in the Honeybee Genome. Mol. Biol. Evol. 2007, 24, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Lenglez, S.; Hermand, D.; Decottignies, A. Genome-wide mapping of nuclear mitochondrial DNA sequences links DNA replication origins to chromosomal double-strand break formation in Schizosaccharomyces pombe. Genome Res. 2010, 20, 1250–1261. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-X.; Liu, J.; Miao, Y.-H.; Huang, D.-W.; Xiao, J.-H. Tracking the Distribution and Burst of Nuclear Mitochondrial DNA Sequences (NUMTs) in Fig Wasp Genomes. Insects 2020, 11, 680. [Google Scholar] [CrossRef] [PubMed]
- Storchova, Z.; Breneman, A.; Cande, J.; Dunn, J.; Burbank, K.S.; O’Toole, E.; Pellman, D. Genome-wide genetic analysis of polyploidy in yeast. Nat. Cell Biol. 2006, 443, 541–547. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Genomic Shotgun Reads | Published Mitogenomes | RNA-Seq |
|---|---|---|---|
| B. breviceps | MF478986 | SRR12527963 | |
| B. confusus | SRR12527987 | ||
| B. consobrinus | MF995069 | SRR12527965 | |
| B. cullumanus | SRR12528003 | ||
| B. difficillimus | SRR12527947 | SRR12527962 | |
| B. haemorrhoidalis | SRR12527992 | SRR12527929 | |
| B. ignitus | DQ870926 | SRR12527935 | |
| B. opulentus | SRR12527970 | SRR12527964 | |
| B. picipes | SRR12528031 | SRR12527936 | |
| B. polaris | SRR12527998 | ||
| B. pyrosoma | MH998260 | SRR12527934 | |
| B. sibiricus | SRR12527981 | SRR12527966 | |
| B. soroeensis | SRR12528009 | ||
| B. superbus | SRR12528014 | SRR12527932 | |
| B. turneri | SRR12528020 | SRR12527933 | |
| B. waltoni | MK252702 | SRR12527961 | |
| B. terrestris | KT368150 |
| Species | Total Length (bp) | Protein-Coding Gene | rRNA Gene | tRNA Gene |
|---|---|---|---|---|
| B. confusus | 19,550 | 13 | 2 | 19 |
| B. cullumanus | 16,418 | 13 | 2 | 20 |
| B. difficillimus | 19,401 | 13 | 2 | 22 |
| B. haemorrhoidalis | 15,358 | 13 | 2 | 22 |
| B. opulentus | 17,293 | 13 | 1 | 20 |
| B. picipes | 18,381 | 13 | 2 | 21 |
| B. polaris | 16,785 | 13 | 2 | 22 |
| B. sibiricus | 29,809 | 13 | 2 | 21 |
| B. soroeensis | 17,138 | 13 | 2 | 22 |
| B. superbus | 15,792 | 13 | 2 | 22 |
| B. turneri | 15,029 | 13 | 2 | 19 |
| Species | Number of NUMTs Fused with Their Flanking Sequences |
|---|---|
| B. breviceps | 18 |
| B. consobrinus | 8 |
| B. difficillimus | 9 |
| B. haemorrhoidalis | 9 |
| B. ignitus | 7 |
| B. opulentus | 7 |
| B. picipes | 8 |
| B. pyrosoma | 5 |
| B. sibiricus | 7 |
| B. superbus | 9 |
| B. turneri | 6 |
| B. waltoni | 12 |
| B. terrestris | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, L.; Sang, H.; Sun, C. Genus-Wide Characterization of Nuclear Mitochondrial DNAs in Bumblebee (Hymenoptera: Apidae) Genomes. Insects 2021, 12, 963. https://doi.org/10.3390/insects12110963
Ding L, Sang H, Sun C. Genus-Wide Characterization of Nuclear Mitochondrial DNAs in Bumblebee (Hymenoptera: Apidae) Genomes. Insects. 2021; 12(11):963. https://doi.org/10.3390/insects12110963
Chicago/Turabian StyleDing, Lele, Huiling Sang, and Cheng Sun. 2021. "Genus-Wide Characterization of Nuclear Mitochondrial DNAs in Bumblebee (Hymenoptera: Apidae) Genomes" Insects 12, no. 11: 963. https://doi.org/10.3390/insects12110963
APA StyleDing, L., Sang, H., & Sun, C. (2021). Genus-Wide Characterization of Nuclear Mitochondrial DNAs in Bumblebee (Hymenoptera: Apidae) Genomes. Insects, 12(11), 963. https://doi.org/10.3390/insects12110963