
Unraveling the Mode of Action of Cordyceps fumosorosea: Potential Biocontrol Agent against Plutella xylostella (Lepidoptera: Plutellidae)

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cordyceps fumosorosea
2.2. Plutella Xylostella Larvae
2.3. Inoculation of P. xylostella with C. fumosorosea
2.4. Observation of Larval Appearance under Light Microscopy (LM)
2.5. Scanning Electron Microscopy (SEM)
2.6. Transmission Electron Microscopy (TEM)
2.7. Conidial Growth Parameter Analysis on Diamondback Moth Larval Cuticle
3. Results
3.1. External Symptoms of P. xylostella Larvae Infected with C. fumosorosea
3.2. Cuticle Topography of P. xylostella Larvae
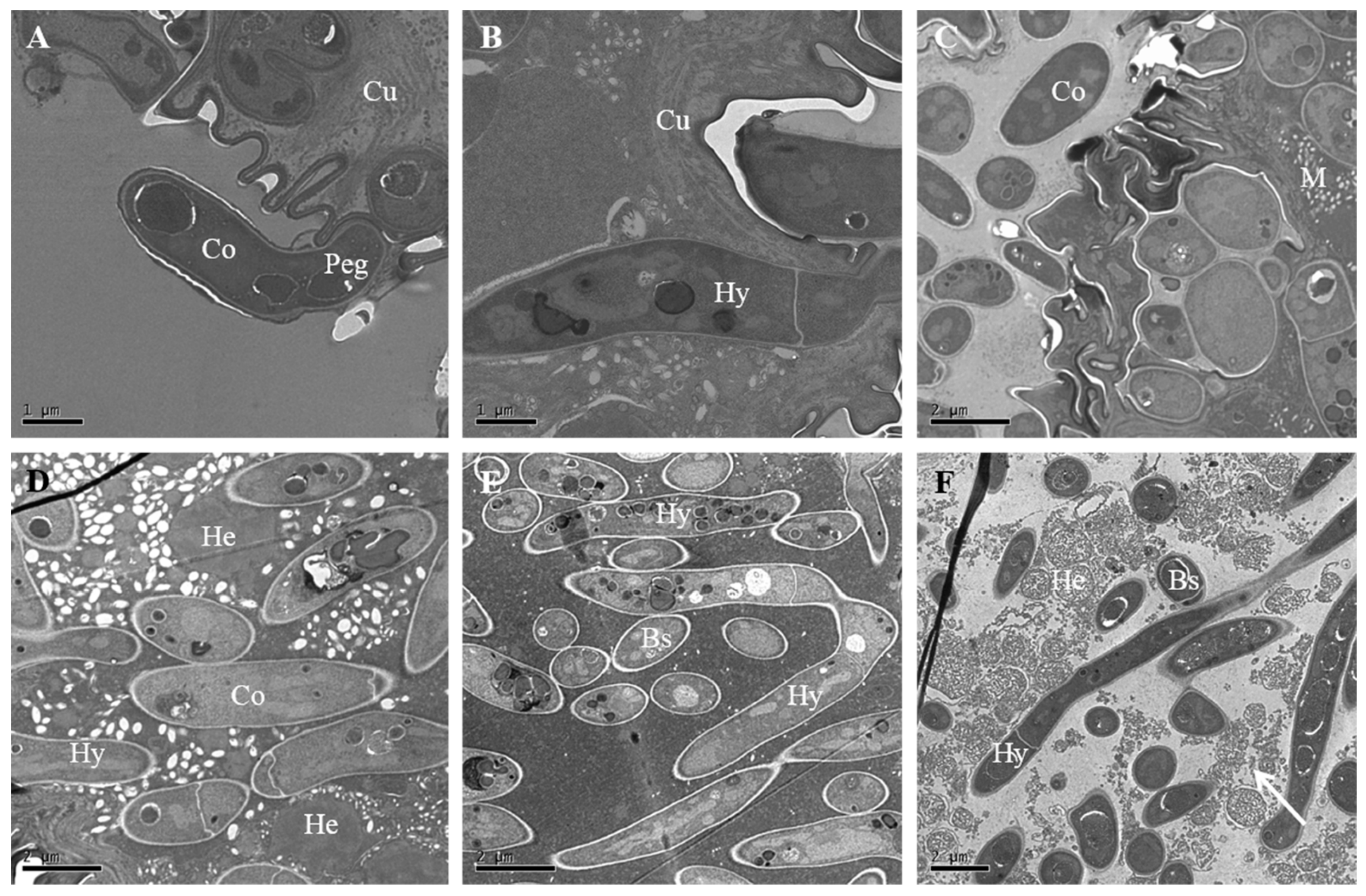
3.3. Fungal Attachment to the Insect Body
3.4. Fungal Penetration through Integument
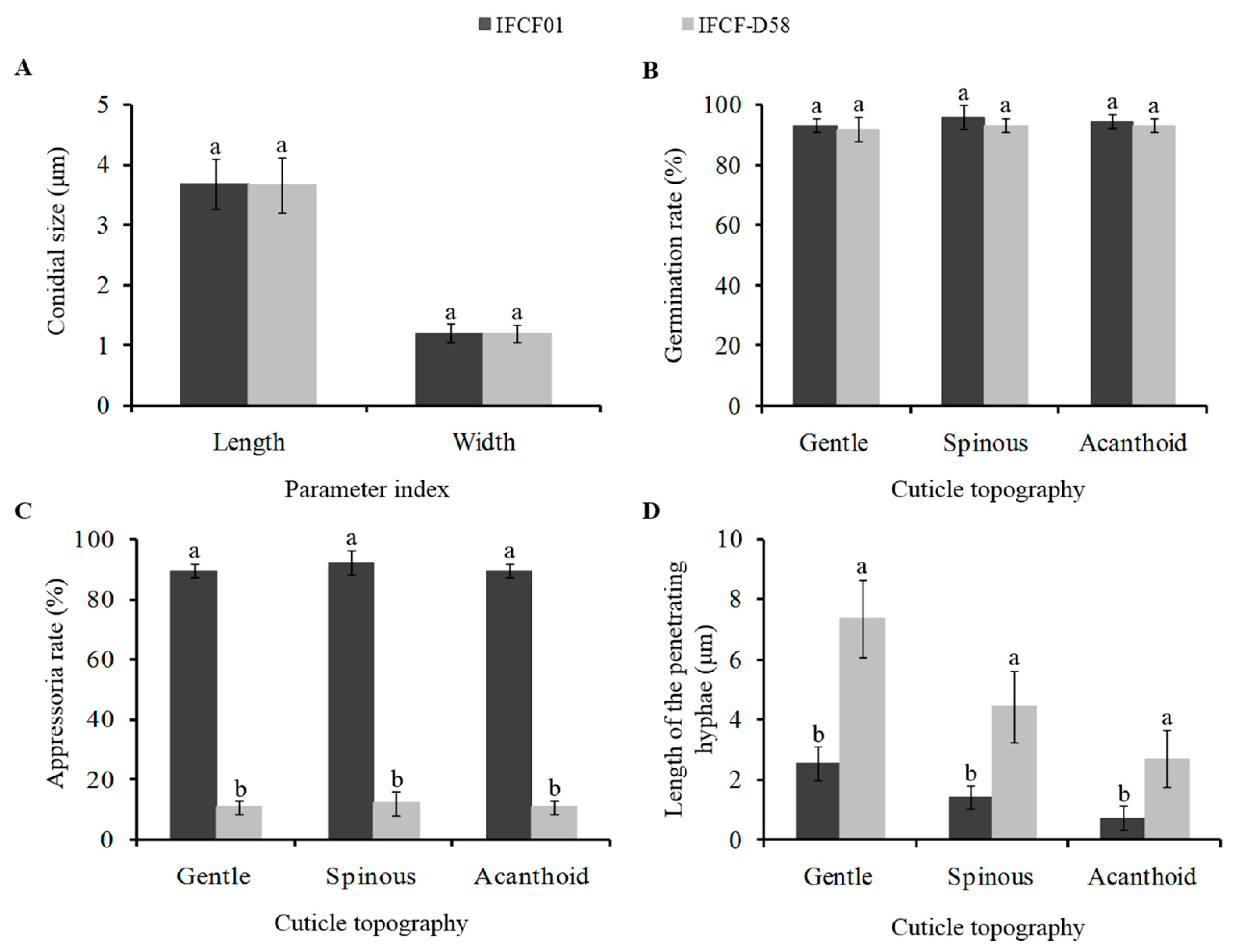
3.5. Physiological Parameters of the Two C. fumosorosea Strains with Different Pathogenicity
3.6. Colonization in the Hemocoel and Internal Tissues
3.7. Fungal Release from the Diamodback Moth Larval Cadavers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Talekar, N.S.; Shelton, A.M. Biology, Ecology, and Management of the Diamondback Moth. Annu. Rev. Entomol. 1993, 38, 275–301. [Google Scholar] [CrossRef]
- Soleymanzade, A.; Khorrami, F.; Forouzan, M. Insecticide toxicity, synergism and resistance in Plutella xylostella (Lepidoptera: Plutellidae). Acta Phytopathol. Entomol. Hungarica 2019, 54, 147–154. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, X.; Shen, J.; Mao, K.; You, H.; Li, J. Susceptibility of field populations of the diamondback moth, Plutella xylostella, to a selection of insecticides in Central China. Pestic. Biochem. Physiol. 2016, 132, 38–46. [Google Scholar] [CrossRef]
- Kang, W.J.; Koo, H.-N.; Jeong, D.-H.; Kim, H.K.; Kim, J.; Kim, G.-H. Functional and genetic characteristics of Chlorantraniliprole resistance in the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Entomol. Res. 2017, 47, 394–403. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Cao, X.; Wang, F.; Yang, Y.; Wu, S.; Wu, Y. Long-term monitoring and characterization of resistance to chlorfenapyr in Plutella xylostella (Lepidoptera: Plutellidae) from China. Pest Manag. Sci. 2019, 75, 591–597. [Google Scholar] [CrossRef]
- Xu, D.; Ali, S.; Huang, Z. Insecticidal activity influence of 20-Hydroxyecdysone on the pathogenicity of Isaria fumosorosea against Plutella xylostella. Biol. Control 2011, 56, 239–244. [Google Scholar] [CrossRef]
- Duarte, R.T.; Gonçalves, K.C.; Espinosa, D.J.L.; Moreira, L.F.; De Bortoli, S.A.; Humber, R.A.; Polanczyk, R.A. Potential of entomopathogenic fungi as biological control agents of Diamondback Moth (Lepidoptera: Plutellidae) and compatibility with chemical insecticides. J. Econ. Entomol. 2016, 109, 594–601. [Google Scholar] [CrossRef] [Green Version]
- AlJabr, A.; Hussain, A.; Rizwan-ul-haq, M. Toxin-Pathogen synergy reshaping detoxification and antioxidant defense mechanism of Oligonychus afrasiaticus (McGregor). Molecules 2018, 23, 1978. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; AlJabr, A.M. Potential synergy between spores of Metarhizium anisopliae and plant secondary metabolite, 1-chlorooctadecane for effective natural acaricide development. Molecules 2020, 25, 1900. [Google Scholar] [CrossRef] [Green Version]
- Kepler, R.M.; Luangsa-ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.-H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A phylogenetically-based nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef]
- Zimmermann, G. The entomopathogenic fungi Isaria farinosa (formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex (formerly Paecilomyces fumosoroseus): Biology, ecology and use in biological control. Biocontrol Sci. Technol. 2008, 18, 865–901. [Google Scholar] [CrossRef]
- Lu, L.H.; He, Y.R.; Wu, Y.J.; Feng, X.; Chen, H. The time–dose mortality model of a Paecilomyces fumosoroseus isolate on the diamondback moth, Plutella xylostella. Acta Entomol. Sin. 2007, 50, 567–573. [Google Scholar]
- Lei, Y.Y.; Lü, L.H.; He, Y.R.; Liang, S.Y. Correlation between biological characteristics of Isaria fumosorosea and its pathogenicity against Plutella xylostella. Acta Phytopathol. Sin. 2010, 37, 217–221. [Google Scholar]
- Xie, M.; Wang, L.; Lu, L.; Zhao, R.; He, Y. Characterisation of Isaria fumosorosea isolates and their virulence toward the Diamondback Moth, Plutella xylostella. Biocontrol Sci. Technol. 2014, 24, 1167–1182. [Google Scholar] [CrossRef]
- Hussain, A.; Tian, M.Y.; He, Y.R.; Bland, J.M.; Gu, W.X. Behavioral and electrophysiological responses of Coptotermes formosanus Shiraki towards entomopathogenic fungal volatiles. Biol. Control 2010, 55, 166–173. [Google Scholar] [CrossRef]
- Tian, J.; Diao, H.; Liang, L.; Hao, C.; Arthurs, S.; Ma, R. Pathogenicity of Isaria fumosorosea to Bemisia tabaci, with some observations on the fungal infection process and host immune response. J. Invertebr. Pathol. 2015, 130, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Wang, Z.; Huang, Y.; Keyhani, N.O.; Huang, Z. Lack of resistance development in Bemisia tabaci to Isaria fumosorosea after multiple generations of selection. Sci. Rep. 2017, 7, 42727. [Google Scholar] [CrossRef] [Green Version]
- Avery, P.B.; Wekesa, V.W.; Hunter, W.B.; Hall, D.G.; McKenzie, C.L.; Osborne, L.S.; Powell, C.A.; Rogers, M.E. Effects of the fungus Isaria fumosorosea (Hypocreales: Cordycipitaceae) on reduced feeding and mortality of the Asian citrus psyllid, Diaphorina citri (Hemiptera: Psyllidae). Biocontrol Sci. Technol. 2011, 21, 1065–1078. [Google Scholar] [CrossRef]
- Keppanan, R.; Krutmuang, P.; Sivaperumal, S.; Hussain, M.; Bamisile, B.S.; Aguila, L.C.R.; Dash, C.K.; Wang, L. Synthesis of mycotoxin protein IF8 by the entomopathogenic fungus Isaria fumosorosea and its toxic effect against adult Diaphorina citri. Int. J. Biol. Macromol. 2019, 125, 1203–1211. [Google Scholar] [CrossRef]
- Huang, W.; Huang, Y.; Hao, Y.; Huang, S.; Gao, T.; Keyhani, N.O.; Huang, Z. Host-dependent contributions of the Cfcdp1 protease gene to virulence in the entomopathogenic fungus Cordyceps fumosorosea. Pest Manag. Sci. 2020, 76, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; AlJabr, A. Susceptibility and immune defence mechanisms of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) against entomopathogenic fungal infections. Int. J. Mol. Sci. 2016, 17, 1518. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Rizwan-ul-Haq, M.; AlJabr, A.M.; Al-Ayedh, H. Evaluation of host–pathogen interactions for selection of entomopathogenic fungal isolates against Oligonychus afrasiaticus (McGregor). BioControl 2020, 65, 185–195. [Google Scholar] [CrossRef]
- Hussain, A.; Tian, M.Y.; He, Y.R.; Ahmed, S. Entomopathogenic fungi disturbed the larval growth and feeding performance of Ocinara varians (Lepidoptera: Bombycidae) larvae. Insect Sci. 2009, 16, 511–517. [Google Scholar] [CrossRef]
- Lei, Y.Y.; He, Y.R.; Lü, L.H. Physiological defense responses of Plutella xylostella (Lepidoptera: Plutellidae) larvae infected by entomopathogenic fungus Isaria fumosorosea. Acta Entomol. Sin. 2011, 54, 887–893. [Google Scholar]
- Feldhaar, H.; Gross, R. Immune reactions of insects on bacterial pathogens and mutualists. Microbes Infect. 2008, 10, 1082–1088. [Google Scholar] [CrossRef]
- Kwon, H.; Bang, K.; Cho, S. Characterization of the hemocytes in larvae of Protaetia brevitarsis seulensis: Involvement of Granulocyte-Mediated Phagocytosis. PLoS ONE 2014, 9, e103620. [Google Scholar] [CrossRef] [Green Version]
- Valanne, S.; Wang, J.-H.; Rämet, M. The Drosophila Toll Signaling Pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokes, B.A.; Yadav, S.; Shokal, U.; Smith, L.C.; Eleftherianos, I. Bacterial and fungal pattern recognition receptors in homologous innate signaling pathways of insects and mammals. Front. Microbiol. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Ali, M.W.; AlJabr, A.M.; Al-Kahtani, S.N. Insights into the Gryllus bimaculatus immune-related transcriptomic profiling to combat naturally invading pathogens. J. Fungi 2020, 6, 232. [Google Scholar] [CrossRef]
- Hussain, A.; Li, Y.F.; Cheng, Y.; Liu, Y.; Chen, C.C.; Wen, S.Y. Immune-related transcriptome of Coptotermes formosanus Shiraki workers: The defense mechanism. PLoS ONE 2013, 8, e69543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaka, M.; Kittakoop, P.; Kirtikara, K.; Hywel-Jones, N.L.; Thebtaranonth, Y. Bioactive Substances from Insect Pathogenic Fungi. Acc. Chem. Res. 2005, 38, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Wang, S. Interaction of entomopathogenic fungi with the host immune system. Dev. Comp. Immunol. 2018, 83, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A. Reprogramming the virulence: Insect defense molecules navigating the epigenetic landscape of Metarhizium robertsii. Virulence 2018, 9, 447–449. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Xu, L. Beauvericin, a Bioactive Compound Produced by Fungi: A Short Review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hu, W.; Xiao, M.; Ou, S.; Hu, Q. Destruxin A Induces and Binds HSPs in Bombyx mori Bm12 Cells. J. Agric. Food Chem. 2017, 65, 9849–9853. [Google Scholar] [CrossRef]
- Mallebrera, B.; Prosperini, A.; Font, G.; Ruiz, M.J. In vitro mechanisms of Beauvericin toxicity: A review. Food Chem. Toxicol. 2018, 111, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Weng, Q.; Zhang, X.; Chen, W.; Hu, Q. Secondary Metabolites and the Risks of Isaria fumosorosea and Isaria farinosa. Molecules 2019, 24, 664. [Google Scholar] [CrossRef] [Green Version]
- Moino, A., Jr.; Alves, S.B.; Lopes, R.B.; Neves, P.M.O.J.; Pereira, R.M.; Vieira, S.A. External development of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in the subterranean termite Heterotermes tenuis. Sci. Agric. 2002, 59, 267–273. [Google Scholar] [CrossRef]
- Wang, Y.; Lei, Z.R.; Zhang, Q.W.; Wen, J. Microscopic observations of infection process of Metarhizium anisopliae on the cuticle of the diamondback moth, Plutella xylostella. Acta Entomol. Sin. 2005, 48, 188–193. [Google Scholar]
- Lei, Y.Y.; Lü, L.H.; He, Y.R.; Wei, B. The symptoms and histopathological changes of Plutella xylostella larvae infected with Isaria fumosorose. Acta Phytopathol. Sin. 2011, 38, 147–152. [Google Scholar]
- Nian, X.; He, Y.; Lu, L.; Zhao, R. Evaluation of alternative Plutella xylostella control by two Isaria fumosorosea conidial formulations—Oil-based formulation and wettable powder—Combined with Bacillus thuringiensis. Pest Manag. Sci. 2015, 71, 1675–1684. [Google Scholar] [CrossRef]
- Nian, X.; He, Y.; Lu, L.; Zhao, R. Evaluation of the time-concentration-mortality responses of Plutella xylostella larvae to the interaction of Isaria fumosorosea with the insecticides beta-cypermethrin and Bacillus thuringiensis. Pest Manag. Sci. 2015, 71, 216–224. [Google Scholar] [CrossRef]
- Safavi, S.A.; Shah, F.A.; Pakdel, A.K.; Reza Rasoulian, G.; Bandani, A.R.; Butt, T.M. Effect of nutrition on growth and virulence of the entomopathogenic fungus Beauveria bassiana. FEMS Microbiol. Lett. 2007, 270, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Urquiza, A.; Keyhani, N. Action on the Surface: Entomopathogenic Fungi versus the Insect Cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Xiong, Q.; Xie, Y.; Zhu, Y.; Xue, J.; Li, J.; Fan, R. Morphological and ultrastructural characterization of Carposina sasakii larvae (Lepidoptera: Carposinidae) infected by Beauveria bassiana (Ascomycota: Hypocreales: Clavicipitaceae). Micron 2013, 44, 303–311. [Google Scholar] [CrossRef]
- St Leger, R.J.; Butt, T.M.; Goettel, M.S.; Staples, R.C.; Roberts, D.W. Production in vitro of appressoria by the entomopathogenic fungus Metarhizium anisopliae. Exp. Mycol. 1989, 13, 274–288. [Google Scholar] [CrossRef]
- Wraight, S.P.; Butt, T.M.; Galaini-Wraight, S.; Allee, L.L.; Soper, R.S.; Roberts, D.W. Germination and infection processes of the entomophthoralean fungus Erynia radicans on the potato leafhopper, Empoasca fabae. J. Invertebr. Pathol. 1990, 56, 157–174. [Google Scholar] [CrossRef]
- Pekrul, S.; Grula, E.A. Mode of infection of the corn earworm (Heliothis zea) by Beauveria bassiana as revealed by scanning electron microscopy. J. Invertebr. Pathol. 1979, 34, 238–247. [Google Scholar] [CrossRef]
- Wang, X.H.; Huang, D.Z.; Yang, Z.Q.; Li, H.P.; Zheng, J.W. Microscopic observations of infection process of Beauveria bassiana on the cuticle of Apriona germari larvae. Sci. Seric. 2009, 35, 374–378. [Google Scholar]
- Askary, H.; Benhamou, N.; Brodeur, J. Ultrastructural and Cytochemical Characterization of Aphid Invasion by the Hyphomycete Verticillium lecanii. J. Invertebr. Pathol. 1999, 74, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Safavi, S. Isolation, Identification and Pathogenicity Assessment of a new Isolate of Entomopathogenic Fungus, Beauveria bassiana in Iran. J. Plant Prot. Res. 2010, 50, 158–163. [Google Scholar] [CrossRef]
- Khan, S.; Nadir, S.; Lihua, G.; Xu, J.; Holmes, K.A.; Dewen, Q. Identification and characterization of an insect toxin protein, Bb70p, from the entomopathogenic fungus, Beauveria bassiana, using Galleria mellonella as a model system. J. Invertebr. Pathol. 2016, 133, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Asaff, A.; Cerda-García-Rojas, C.; de la Torre, M. Isolation of dipicolinic acid as an insecticidal toxin from Paecilomyces fumosoroseus. Appl. Microbiol. Biotechnol. 2005, 68, 542–547. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, J.; Chen, C.; Teng, J.; Wang, C.; Luo, D. Structure and biosynthesis of fumosorinone, a new protein tyrosine phosphatase 1B inhibitor firstly isolated from the entomogenous fungus Isaria fumosorosea. Fungal Genet. Biol. 2015, 81, 191–200. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, Y.; Hussain, A.; Guan, Z.; Wang, D.; Jaleel, W.; Lyu, L.; He, Y. Unraveling the Mode of Action of Cordyceps fumosorosea: Potential Biocontrol Agent against Plutella xylostella (Lepidoptera: Plutellidae). Insects 2021, 12, 179. https://doi.org/10.3390/insects12020179
Lei Y, Hussain A, Guan Z, Wang D, Jaleel W, Lyu L, He Y. Unraveling the Mode of Action of Cordyceps fumosorosea: Potential Biocontrol Agent against Plutella xylostella (Lepidoptera: Plutellidae). Insects. 2021; 12(2):179. https://doi.org/10.3390/insects12020179
Chicago/Turabian StyleLei, Yanyuan, Abid Hussain, Zhaoying Guan, Desen Wang, Waqar Jaleel, Lihua Lyu, and Yurong He. 2021. "Unraveling the Mode of Action of Cordyceps fumosorosea: Potential Biocontrol Agent against Plutella xylostella (Lepidoptera: Plutellidae)" Insects 12, no. 2: 179. https://doi.org/10.3390/insects12020179
APA StyleLei, Y., Hussain, A., Guan, Z., Wang, D., Jaleel, W., Lyu, L., & He, Y. (2021). Unraveling the Mode of Action of Cordyceps fumosorosea: Potential Biocontrol Agent against Plutella xylostella (Lepidoptera: Plutellidae). Insects, 12(2), 179. https://doi.org/10.3390/insects12020179