
Seasonal Abundance and Diversity of Egg Parasitoids of Halyomorpha halys in Kiwifruit Orchards in China

 , , , and
, , , and 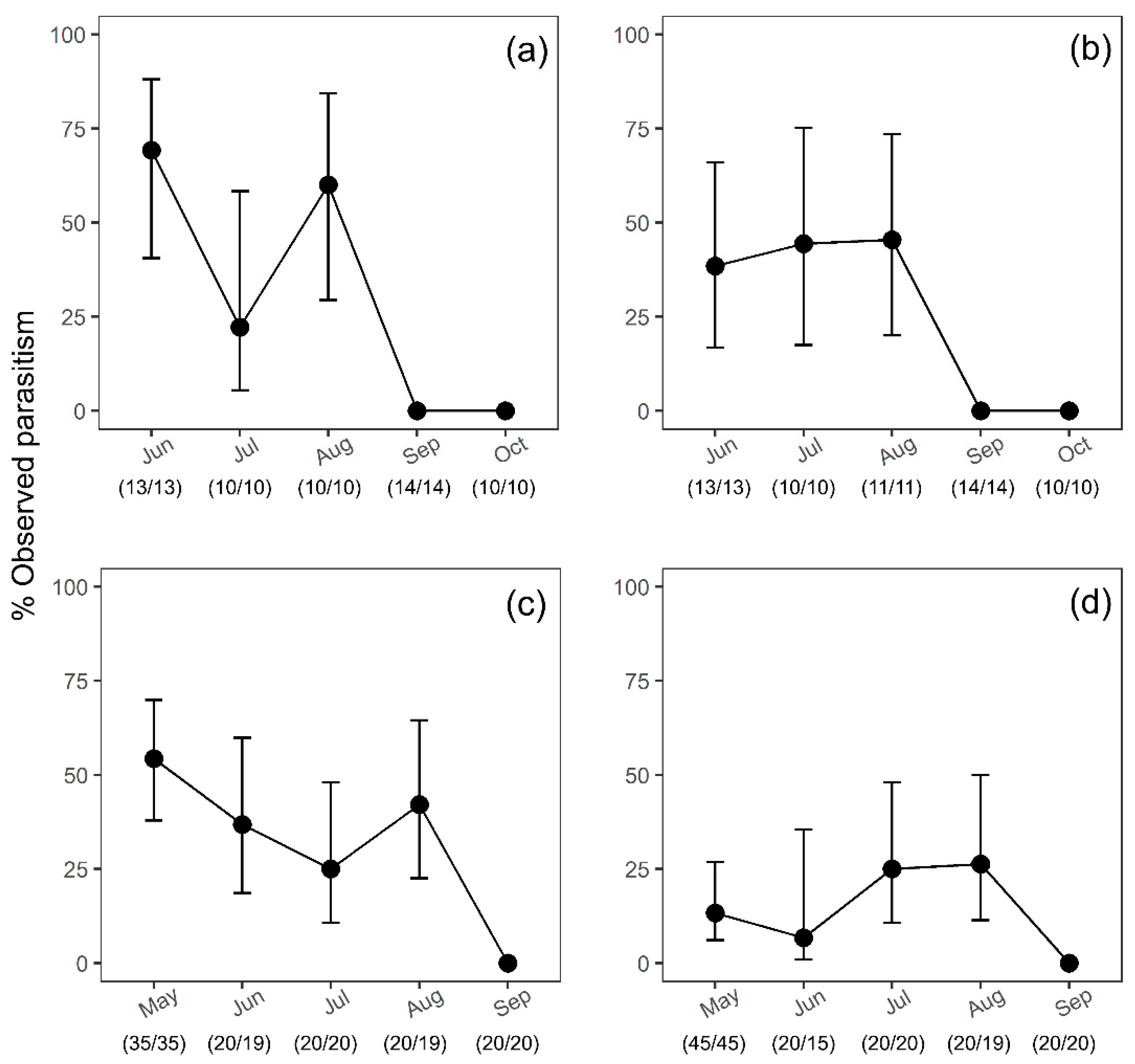{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Egg Parasitoid Surveys in Kiwifruit Orchards
2.3. Parasitoid Identification
2.4. Data Analysis
3. Results
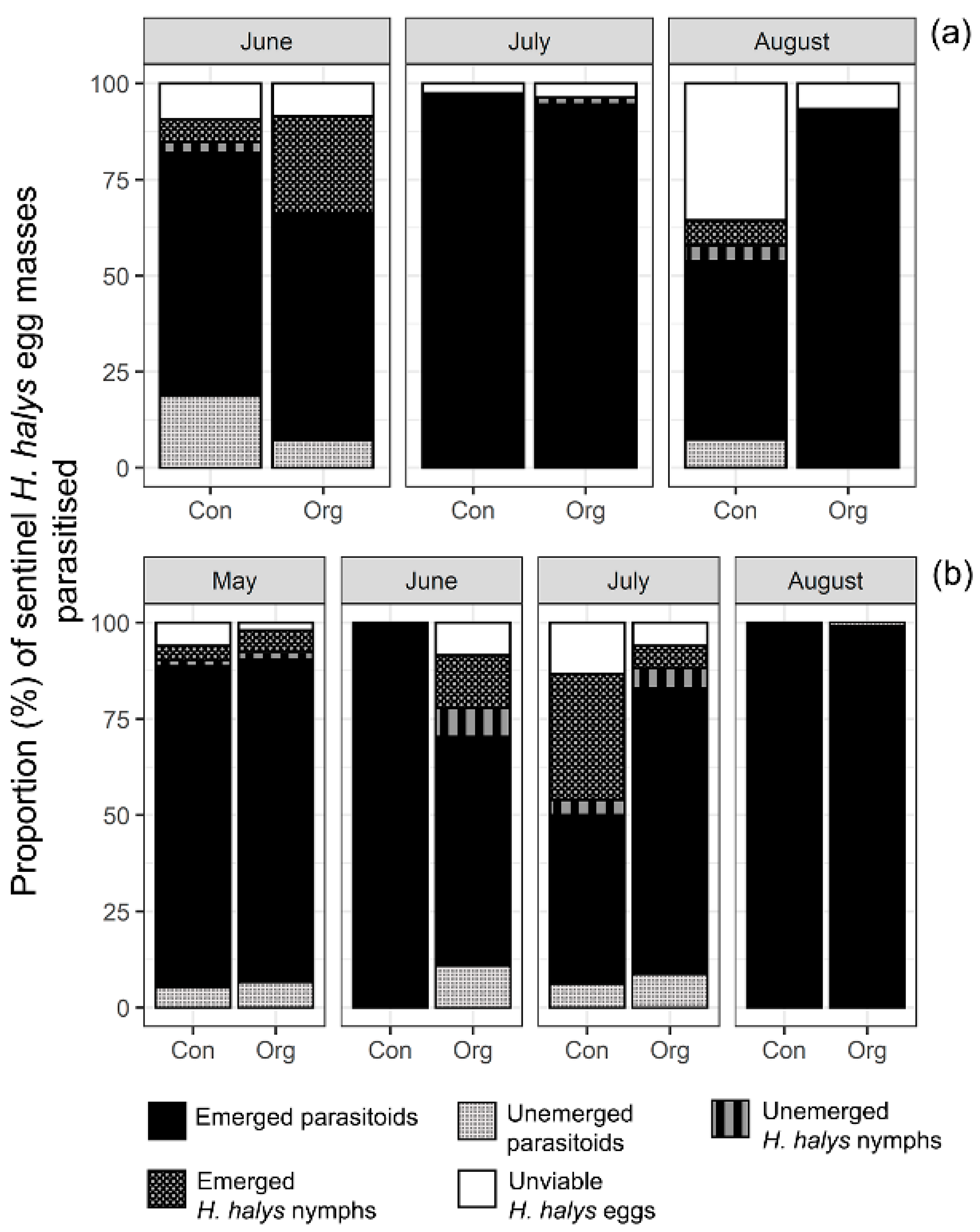
3.1. Seasonal Parasitism of H. halys Sentinel Egg Masses
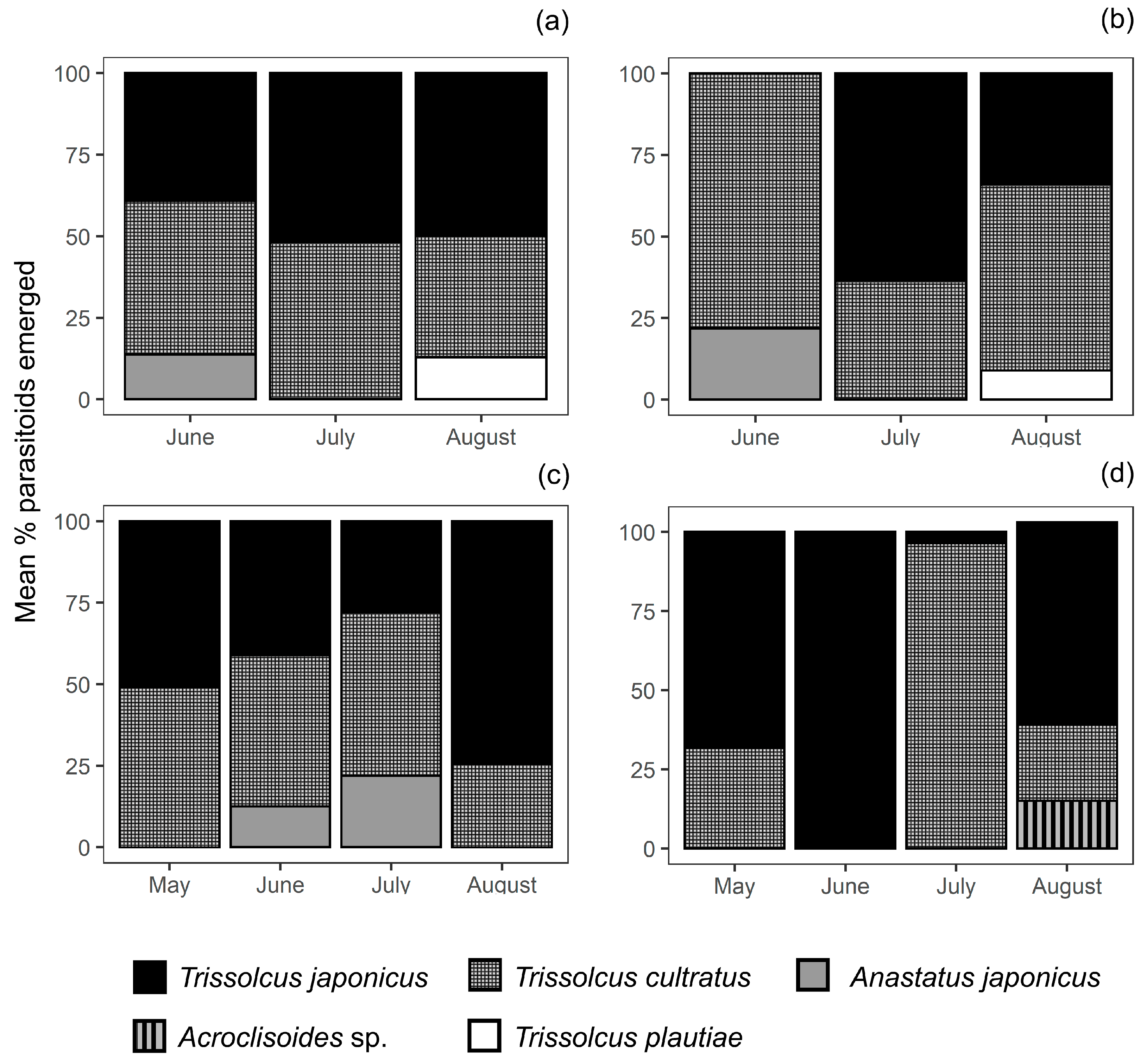
3.2. Parasitoid Species Recovered and Abundance
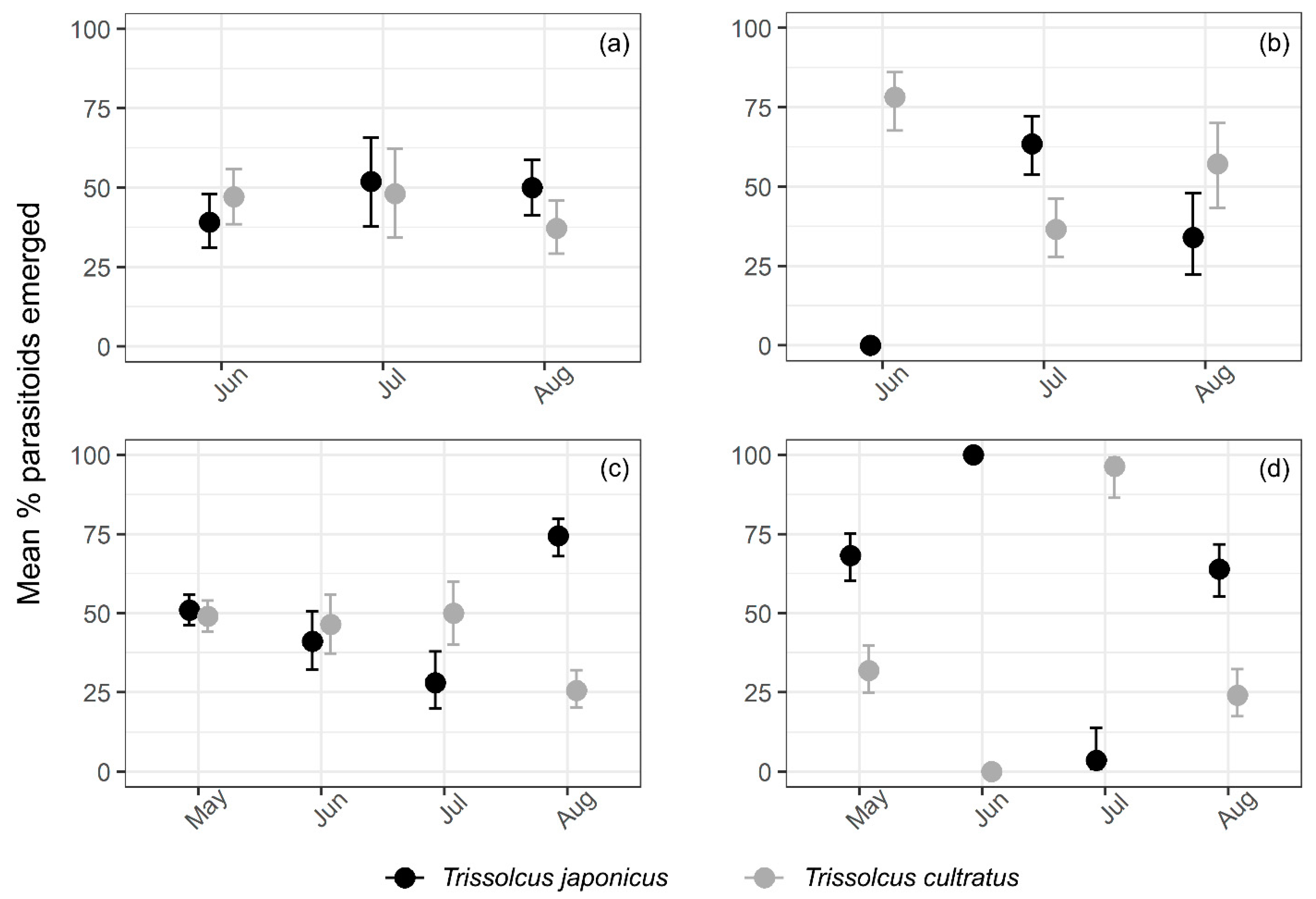
3.3. Relative Abundance of T. japonicus and T. cultratus in Kiwifruit Orchards
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Èntomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Haye, T.; Gariepy, T.; Hoelmer, K.A.; Rossi, J.-P.; Streito, J.-C.; Tassus, X.; Desneux, N. Range expansion of the invasive brown marmorated stink bug, Halyomorpha halys: An increasing threat to field, fruit and vegetable crops worldwide. J. Pest Sci. 2015, 88, 665–673. [Google Scholar] [CrossRef]
- Faúndez, E.; Rider, D. The brown marmorated stink bug Halyomorpha halys (Stål 1885) (Heteroptera: Pentatomidae) in Chile. Arq. Entomol. 2017, 17, 305–330. [Google Scholar]
- Gapon, D.A. First records of the brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera, Pentatomidae) in Russia, Abkhazia, and Georgia. Èntomol. Rev. 2016, 96, 1086–1088. [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, Biology, Ecology, and Management. Annu. Rev. Èntomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, E.J.; Venugopal, P.D.; Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Host plant use by the invasive Halyomorpha halys (Stål) on woody ornamental trees and shrubs. PLoS ONE 2016, 11, e0149975. [Google Scholar] [CrossRef]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Mi, Q.; Zhang, J.; Han, X.; Yan, Y.; Zhang, B.; Li, D.; Zhang, F. Releases of Trissolcus japonicus and Anastatus sp. for suppression of Halyomorpha halys in kiwifruit orchards. In Proceedings of the 5th International Symposium on Biological Control of Arthropods, Langkawi, Malaysia, 11–15 September 2017; p. 332. [Google Scholar]
- Chen, J.H.; Avila, G.A.; Zhang, F.; Guo, L.F.; Sandanayaka, M.; Mi, Q.-Q.; Shi, S.-S.; Zhang, J.-P. Field cage assessment of feeding damage by Halyomorpha halys on kiwifruit orchards in China. J. Pest Sci. 2020, 93, 953–963. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, Z.; Zhang, S. Occurrence and control of Halyomorpha halys in kiwifruit orchard. Northwest Hortic. 2000, 2, 38. [Google Scholar]
- Bariselli, M.; Bugiani, R.; Maistrello, L. Distribution and damage caused by Halyomorpha halys in Italy. EPPO Bull. 2016, 46, 332–334. [Google Scholar] [CrossRef]
- Bernardinelli, I.; Malossini, G.; Benvenuto, L. Halyomorpha halys: Risultati preliminari di alcune attività sperimentali condotte in Friuli Venezia Giulia nel 2016. Not. ERSA 2017, 1, 24–26. [Google Scholar]
- Andreadis, S.S.; Navrozidis, E.I.; Farmakis, A.; Pisalidis, A. First Evidence of Halyomorpha halys (Hemiptera: Pentatomidae) infesting kiwifruit (Actinidia chinensis) in Greece. J. Èntomol. Sci. 2018, 53, 402–405. [Google Scholar] [CrossRef]
- Burne, A. Pest Risk Assessment: Halyomorpha halys (Brown Marmorated Stink Bug); Ministry for Primary Industries: Wellington, New Zealand, 2019. Available online: https://www.mpi.govt.nz/dmsdocument/38075/direct (accessed on 22 February 2021).
- Haye, T.; Fischer, S.; Zhang, J.; Gariepy, T. Can native egg parasitoids adopt the invasive brown marmorated stink bug, Halyomorpha halys (Heteroptera: Pentatomidae), in Europe? J. Pest Sci. 2015, 88, 693–705. [Google Scholar] [CrossRef]
- Herlihy, M.V.; Talamas, E.J.; Weber, D.C. Attack and success of native and exotic parasitoids on eggs of Halyomorpha halys in three maryland habitats. PLoS ONE 2016, 11, e0150275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzini, D.T.; Santacruz, E.C.N.; Koch, R.L. Predation and parasitism of Halyomorpha halys (Hemiptera: Pentatomidae) Eggs in Minnesota. Environ. Èntomol. 2018, 47, 812–821. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal parasitism and host specificity of Trissolcus japonicus in northern China. J. Pest Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Li, W.; Mi, Q.; Zhang, F.; Shi, S.; Zhang, J. A newly reported parasitoid, Pentatomophaga latifascia (Diptera: Tachinidae), of adult Halyomorpha halys in Beijing, China. Insects 2020, 11, 666. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-Q.; Yao, Y.-X.; Qiu, L.-F.; Li, Z.-X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Èntomol. Soc. Am. 2009, 102, 39–47. [Google Scholar] [CrossRef]
- Haye, T.; Moraglio, S.T.; Stahl, J.; Visentin, S.; Gregorio, T.; Tavella, L. Fundamental host range of Trissolcus japonicus in Europe. J. Pest Sci. 2020, 93, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Hedstrom, C.; Lowenstein, D.; Andrews, H.; Bai, B.; Wiman, N. Pentatomid host suitability and the discovery of introduced populations of Trissolcus japonicus in Oregon. J. Pest Sci. 2017, 90, 1169–1179. [Google Scholar] [CrossRef]
- Lara, J.R.; Pickett, C.H.; Kamiyama, M.T.; Figueroa, S.; Romo, M.; Cabanas, C.; Bazurto, V.; Strode, V.; Briseno, K.; Lewis, M.; et al. Physiological host range of Trissolcus japonicus in relation to Halyomorpha halys and other pentatomids from California. BioControl 2019, 64, 513–528. [Google Scholar] [CrossRef]
- Milnes, J.M.; Beers, E.H. Trissolcus japonicus (Hymenoptera: Scelionidae) causes low levels of parasitism in three north american pentatomids under field conditions. J. Insect Sci. 2019, 19, 15. [Google Scholar] [CrossRef] [Green Version]
- Abram, P.K.; Talamas, E.J.; Acheampong, S.; Mason, P.G.; Gariepy, T.D. First detection of the samurai wasp, Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae), in Canada. J. Hymenopt. Res. 2019, 68, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Milnes, J.M.; Wiman, N.G.; Talamas, E.J.; Brunner, J.F.; Hoelmer, K.A.; Buffington, M.L.; Beers, E.H. Discovery of an exotic egg parasitoid of the brown marmorated stink bug, Halyomorpha halys (Stål) in the Pacific Northwest. Proc. Èntomol. Soc. Wash. 2016, 118, 466–470. [Google Scholar] [CrossRef]
- Peverieri, G.S.; Talamas, E.; Bon, M.C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.F.; Hoelmer, K. Two asian egg parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.-C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First discovery of adventive populations of Trissolcus japonicus in Europe. J. Pest Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Talamas, E.J.; Herlihy, M.V.; Dieckhoff, C.; Hoelmer, K.A.; Buffington, M.; Bon, M.-C.; Weber, D.C. Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) emerges in North America. J. Hymenopt. Res. 2015, 43, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini-Peverieri, G.; Dieckhoff, C.; Giovannini, L.; Marianelli, L.; Roversi, P.F.; Hoelmer, K. Rearing Trissolcus japonicus and Trissolcus mitsukurii for biological control of Halyomorpha halys. Insects 2020, 11, 787. [Google Scholar] [CrossRef]
- Charles, J.G.; Avila, G.A.; Hoelmer, K.A.; Hunt, S.; Gardner-Gee, R.; MacDonald, F.; Davis, V. Experimental assessment of the biosafety of Trissolcus japonicus in New Zealand, prior to the anticipated arrival of the invasive pest Halyomorpha halys. BioControl 2019, 64, 367–379. [Google Scholar] [CrossRef]
- EPA (Environmental Protection Authority). Application to Seek Pre-Approval to Release Trissolcus japonicus (the Samurai Wasp) as a Biological Control Agent for Brown Marmorated Stink Bug (Halyomorpha halys) Should It Arrive in New Zealand. Decision. 2018. Available online: https://www.epa.govt.nz/assets/FileAPI/hsno-ar/APP203336/0ed5350647/APP203336-Decision.pdf (accessed on 20 January 2021).
- Bergman, J.M.; Tingey, W.M. Aspects of interaction between plant genotypes and biological control. Bull. Èntomol. Soc. Am. 1979, 25, 275–279. [Google Scholar] [CrossRef]
- Tillman, G.; Toews, M.; Blaauw, B.; Sial, A.; Cottrell, T.; Talamas, E.; Buntin, D.; Joseph, S.; Balusu, R.; Fadamiro, H.; et al. Parasitism and predation of sentinel eggs of the invasive brown marmorated stink bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae), in the southeastern US. Biol. Control 2020, 145, 104247. [Google Scholar] [CrossRef]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.R.; Shrewsbury, P.M. Field surveys of egg mortality and indigenous egg parasitoids of the brown marmorated stink bug, Halyomorpha halys, in ornamental nurseries in the mid-Atlantic region of the USA. J. Pest Sci. 2017, 90, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Dieckhoff, C.; Tatman, K.M.; Hoelmer, K.A. Natural biological control of Halyomorpha halys by native egg parasitoids: A multi-year survey in northern Delaware. J. Pest Sci. 2017, 90, 1143–1158. [Google Scholar] [CrossRef]
- Holthouse, M.C.; Schumm, Z.R.; Talamas, E.J.; Spears, L.R.; Alston, D.G. Surveys in northern Utah for egg parasitoids of Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) detect Trissolcus japonicus (Ashmead) (Hymenoptera: Scelionidae). Biodivers. Data J. 2020, 8, e53363. [Google Scholar] [CrossRef] [PubMed]
- Moraglio, S.T.; Tortorici, F.; Pansa, M.G.; Castelli, G.; Pontini, M.; Scovero, S.; Visentin, S.; Tavella, L. A 3-year survey on parasitism of Halyomorpha halys by egg parasitoids in northern Italy. J. Pest Sci. 2019, 93, 183–194. [Google Scholar] [CrossRef]
- Zapponi, L.; Bon, M.; Fouani, J.; Anfora, G.; Schmidt, S.; Falagiarda, M. Assemblage of the egg parasitoids of the invasive stink bug Halyomorpha halys: Insights on plant host associations. Insects 2020, 11, 588. [Google Scholar] [CrossRef] [PubMed]
- Quinn, N.F.; Talamas, E.J.; Leskey, T.C.; Bergh, J.C. Sampling methods for adventive Trissolcus japonicus (Hymenoptera: Scelionidae) in a wild tree host of Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Èntomol. 2019, 112, 1997–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, N.F.; Talamas, E.J.; Acebes-Doria, A.L.; Leskey, T.C.; Bergh, J.C. Vertical sampling in tree canopies for Halyomorpha halys (Hemiptera: Pentatomidae) life stages and its egg parasitoid, Trissolcus japonicus (Hymenoptera: Scelionidae). Environ. Èntomol. 2019, 48, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Kaser, J.M.; Akotsen-Mensah, C.; Talamas, E.J.; Nielsen, A.L. First report of Trissolcus japonicus parasitizing Halyomorpha halys in North American agriculture. Fla. Èntomol. 2018, 101, 680–684. [Google Scholar] [CrossRef]
- Talamas, E.J.; Johnson, N.F.; Buffington, M. Key to Nearctic species of Trissolcus Ashmead (Hymenoptera, Scelionidae), natural enemies of native and invasive stink bugs (Hemiptera, Pentatomidae). J. Hymenopt. Res. 2015, 43, 45–110. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Gibson, G.A.; Tang, L.; Xiang, J. Review of the species of Anastatus (Hymenoptera: Eupelmidae) known from China, with description of two new species with brachypterous females. Zootaxa 2020, 4767, 351–401. [Google Scholar] [CrossRef] [PubMed]
- Greathead, D. Benefits and risks of classical biological control. In Biological Control: Benefits and Risks; Hokkanen, H., Lynch, J., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 53–63. [Google Scholar]
- Bale, J.S.; Van Lenteren, J.C.; Bigler, F. Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci. 2007, 363, 761–776. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, P.; Mason, P.G.; Babendreier, D. Benefits and risks of exotic biological control agents. BioControl 2011, 56, 681–698. [Google Scholar] [CrossRef]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.; Shrewsbury, P.M. Sentinel eggs underestimate rates of parasitism of the exotic brown marmorated stink bug, Halyomorpha halys. Biol. Control 2014, 78, 61–66. [Google Scholar] [CrossRef]
- Ogburn, E.C.; Bessin, R.; Dieckhoff, C.; Dobson, R.; Grieshop, M.; Hoelmer, K.A.; Mathews, C.; Moore, J.; Nielsen, A.L.; Poley, K.; et al. Natural enemy impact on eggs of the invasive brown marmorated stink bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae), in organic agroecosystems: A regional assessment. Biol. Control 2016, 101, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Roversi, P.; Binazzi, F.; Marianelli, L.; Costi, E.; Maistrello, L.; Sabbatini-Peverieri, G. Searching for native egg-parasitoids of the invasive alien species Halyomorpha halys Stål (Heteroptera, Pentatomidae) in Southern Europe. Redia 2016, 99, 63–70. [Google Scholar] [CrossRef]
- Benvenuto, L.; Bernardinelli, I.; Malossini, G.; Sabbatini Peverieri, G.; Marianelli, L.; Roversi, P.F. Monitoraggio di ovature par-assitizzate di Halyomorpha halys in un actinidieto biologico in Friuli Venezia Giulia nel biennio 2018–2019. Atti Giornate Fitopatol. 2020, 1, 161–166. [Google Scholar]
- Qiu, L.; Yang, Z.; Tao, W. Biology and population dynamics of Trissolcus halyomorphae. Sci. Silvae Sin. 2007, 43, 62–65. [Google Scholar]
- Acebes-Doria, A.L.; Leskey, T.C.; Bergh, J.C. Host plant effects on Halyomorpha halys (Hemiptera: Pentatomidae) nymphal development and survivorship. Environ. Èntomol. 2016, 45, 663–670. [Google Scholar] [CrossRef]
- Morrison, W.R., III; Blaauw, B.R.; Nielsen, A.L.; Talamas, E.; Leskey, T.C. Predation and parasitism by native and exotic natural enemies of Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) eggs augmented with semiochemicals and differing host stimuli. Biol. Control 2018, 121, 140–150. [Google Scholar] [CrossRef]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef]
- Orr, D.B.; Boethel, D.J.; Layton, M.B. Effect of insecticide applications in soybeans on Trissolcus basalis (Hymenoptera: Scelionidae). J. Econ. Èntomol. 1989, 82, 1078–1084. [Google Scholar] [CrossRef]
- Hewa-Kapuge, S.; McDougall, S.; Hoffmann, A.A. Effects of methoxyfenozide, indoxacarb, and other insecticides on the beneficial egg parasitoid Trichogramma nr. brassicae (Hymenoptera: Trichogrammatidae) under laboratory and field conditions. J. Econ. Èntomol. 2003, 96, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Turchen, L.M.; Golin, V.; Butnariu, A.R.; Guedes, R.N.C.; Pereira, M.J.B. Lethal and sublethal effects of insecticides on the egg parasitoid Telenomus podisi (Hymenoptera: Platygastridae). J. Econ. Èntomol. 2015, 109, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Ludwick, D.C.; Patterson, J.; Leake, L.B.; Carper, L.; Leskey, T.C. Integrating Trissolcus japonicus (Ashmead, 1904) (Hymenoptera: Scelionidae) into management programs for Halyomorpha halys (Stål, 1855) (Hemiptera: Pentatomidae) in apple orchards: Impact of insecticide applications and spray patterns. Insects 2020, 11, 833. [Google Scholar] [CrossRef] [PubMed]
- Lowenstein, D.M.; Andrews, H.; Mugica, A.; Wiman, N.G. Sensitivity of the egg parasitoid Trissolcus japonicus (Hymenoptera: Scelionidae) to field and laboratory-applied insecticide residue. J. Econ. Èntomol. 2019, 112, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Lim, U.T.; Mahmoud, A.M. Ecotoxicological effect of fenitrothion on Trissolcus nigripedius (Hymenoptera: Scelionidae) an egg parasitoid of Dolycoris baccarum (Hemiptera: Pentatomidae). J. Asia Pac. Èntomol. 2008, 11, 207–210. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avila, G.A.; Chen, J.; Li, W.; Alavi, M.; Mi, Q.; Sandanayaka, M.; Zhang, F.; Zhang, J. Seasonal Abundance and Diversity of Egg Parasitoids of Halyomorpha halys in Kiwifruit Orchards in China. Insects 2021, 12, 428. https://doi.org/10.3390/insects12050428
Avila GA, Chen J, Li W, Alavi M, Mi Q, Sandanayaka M, Zhang F, Zhang J. Seasonal Abundance and Diversity of Egg Parasitoids of Halyomorpha halys in Kiwifruit Orchards in China. Insects. 2021; 12(5):428. https://doi.org/10.3390/insects12050428
Chicago/Turabian StyleAvila, Gonzalo A., Juhong Chen, Wenjing Li, Maryam Alavi, Qianqian Mi, Manoharie Sandanayaka, Feng Zhang, and Jinping Zhang. 2021. "Seasonal Abundance and Diversity of Egg Parasitoids of Halyomorpha halys in Kiwifruit Orchards in China" Insects 12, no. 5: 428. https://doi.org/10.3390/insects12050428
APA StyleAvila, G. A., Chen, J., Li, W., Alavi, M., Mi, Q., Sandanayaka, M., Zhang, F., & Zhang, J. (2021). Seasonal Abundance and Diversity of Egg Parasitoids of Halyomorpha halys in Kiwifruit Orchards in China. Insects, 12(5), 428. https://doi.org/10.3390/insects12050428