
Host Range and Impact of Dichrorampha aeratana, the First Potential Biological Control Agent for Leucanthemum vulgare in North America and Australia

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
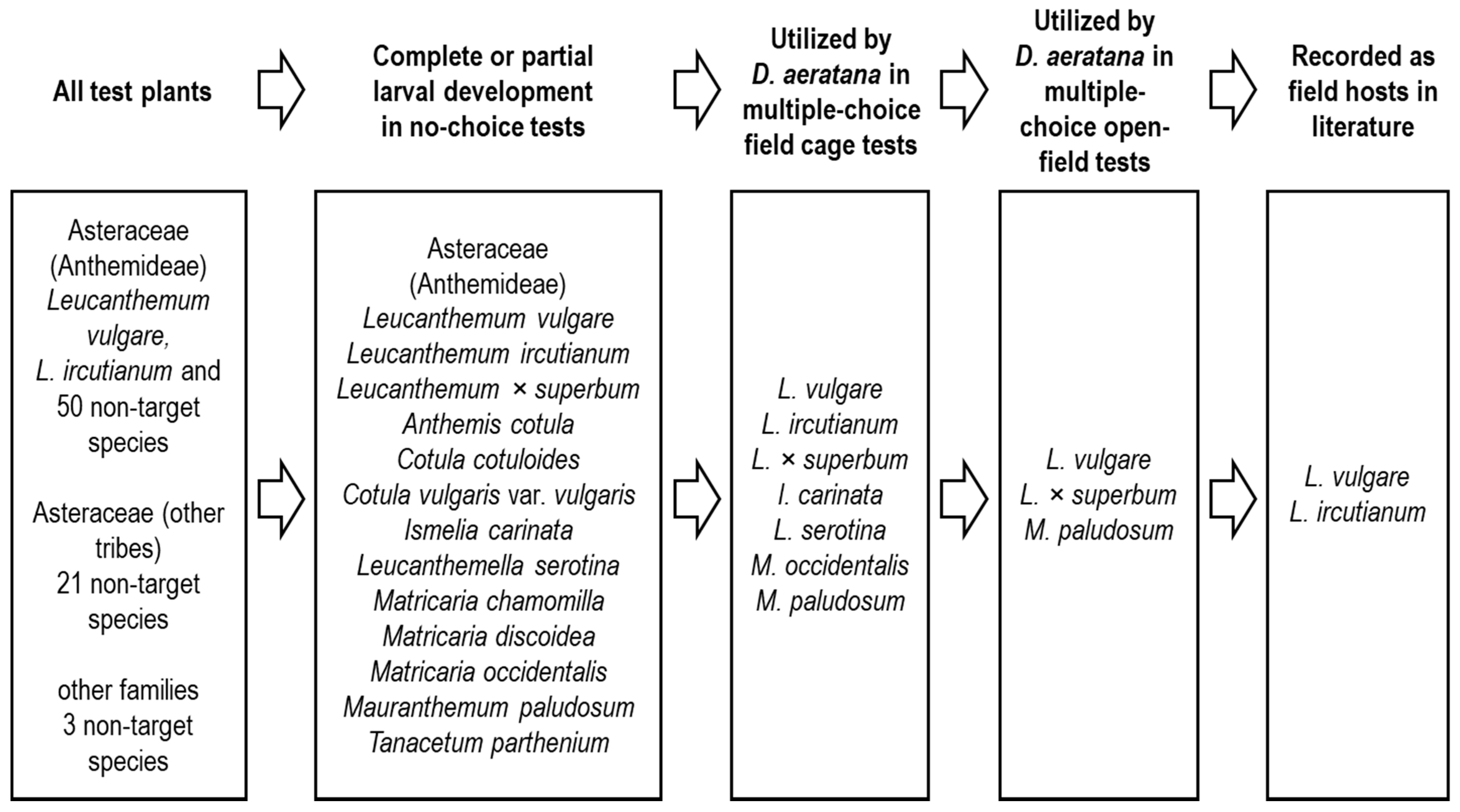
2.2. Test Plant Species
2.3. Host-Range Testing
2.3.1. No-Choice Larval Development Tests
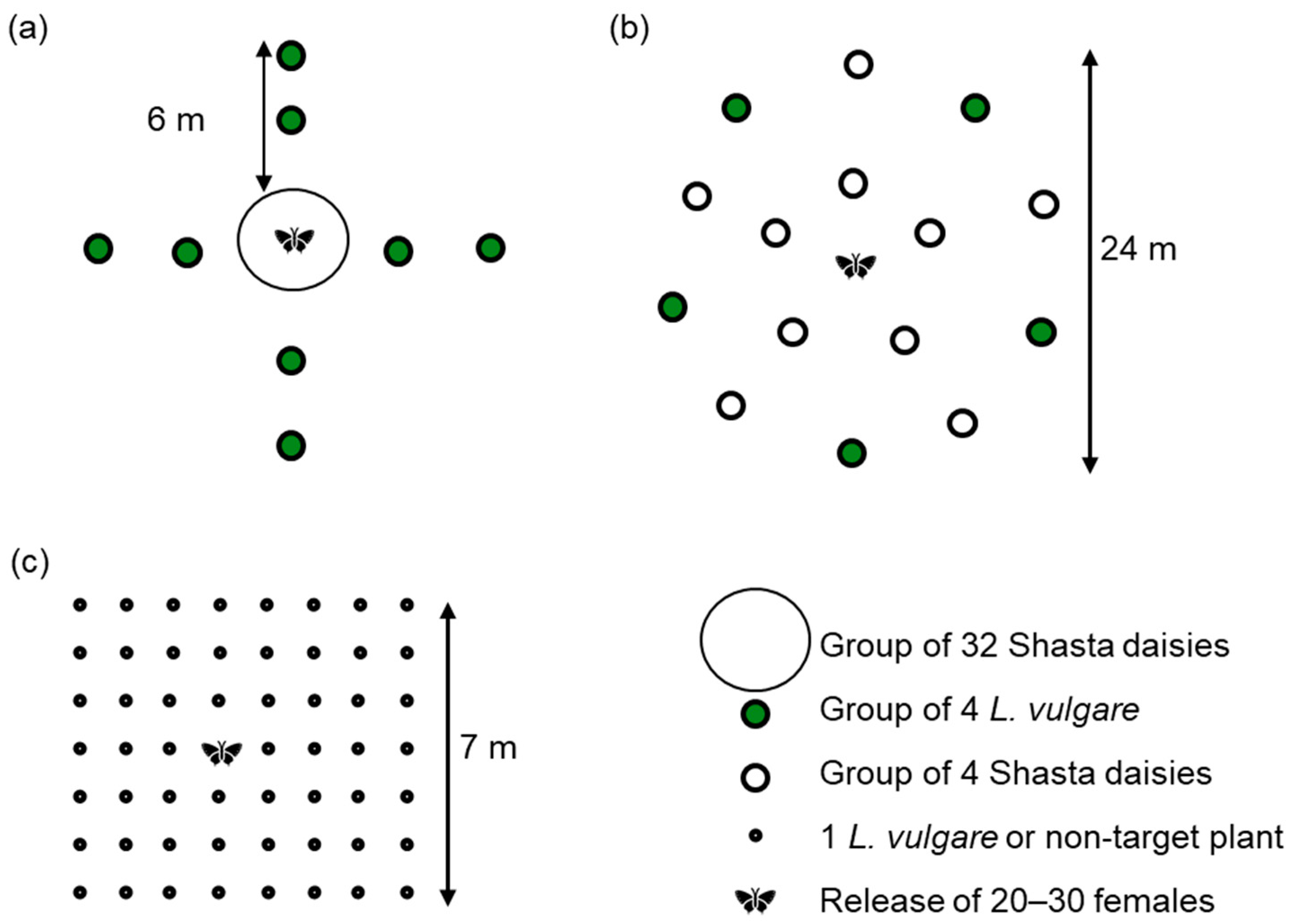
2.3.2. Oviposition and Development Test with Shasta Daisy in Field Cages
2.3.3. Multiple-Choice Oviposition and Larval Development Tests in Field Cages
2.3.4. Multiple-Choice Oviposition and Larval Development Tests under Open-Field Conditions
2.4. Impact of Dichrorampha aeratana on Shasta Daisy and Impact of D. aeratana and Plant Competition on Leucanthemum vulgare
2.5. Statistical Analyses
3. Results
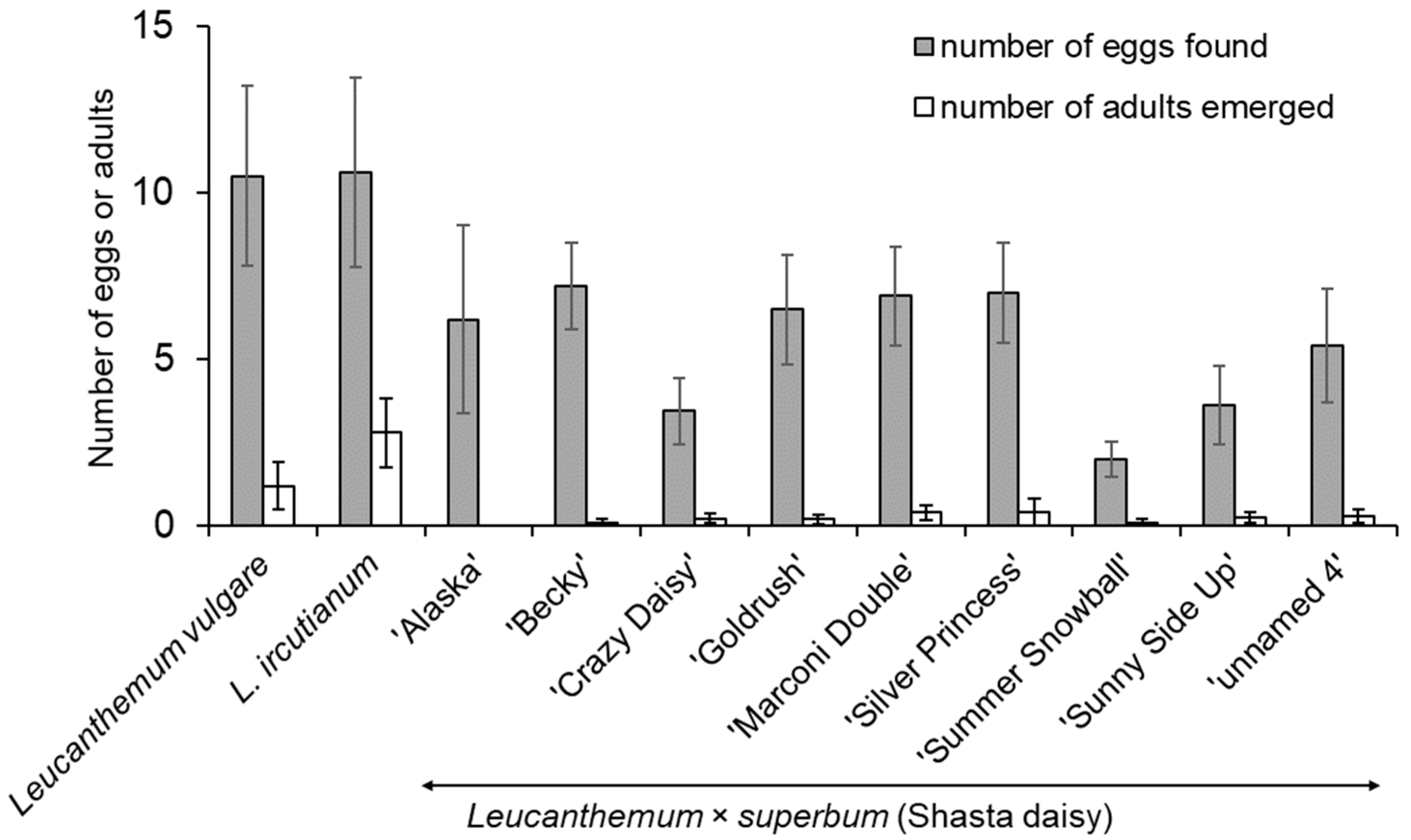
3.1. No-Choice Larval Development Tests
3.2. Oviposition and Development Test with Shasta Daisy in Field Cages
3.3. Multiple-Choice Oviposition and Larval Development Tests in Field Cages
3.4. Multiple-Choice Oviposition and Larval Development Tests under Open-Field Conditions
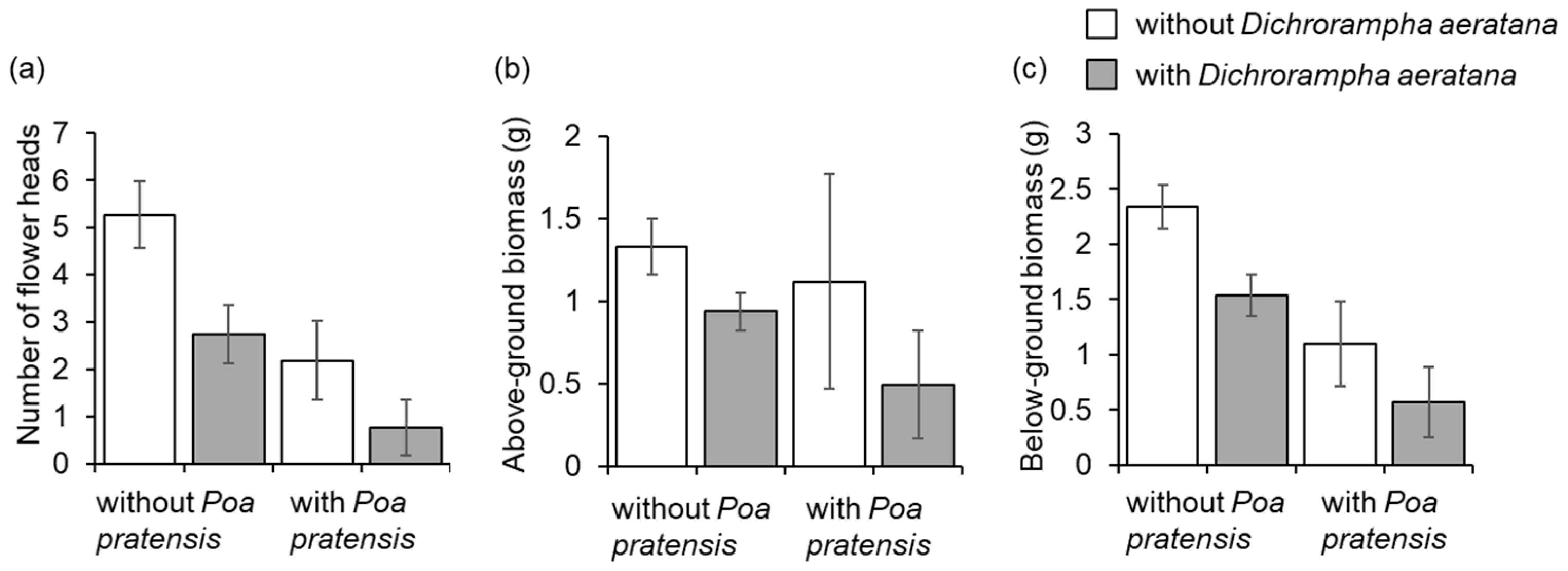
3.5. Impact of Dichrorampha aeratana on Shasta Daisy and Impact of D. aeratana and Plant Competition on Leucanthemum vulgare
4. Discussion
4.1. Host-Range of Dichrorampha aeratana
4.2. Impact of Dichrorampha aeratana on the Non-Target Species Shasta Daisy
4.3. Impact of Dichrorampha aeratana on the Target Species Leucanthemum vulgare
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holm, L.; Pancho, J.V.; Herberger, J.P.; Plucknett, D.L. A geographical Atlas of World Weeds; John Wiley and Sons: New York, NY, USA, 1979. [Google Scholar]
- Clements, D.R.; Cole, D.E.; Darbyshire, S.; King, J.; McClay, A. The biology of Canadian weeds. 128. Leucanthemum vulgare Lam. Can. J. Plant Sci. 2004, 84, 343–363. [Google Scholar] [CrossRef]
- McDougall, K.; Wright, G.; Peach, E. Coming to terms with Ox-eye Daisy (Leucanthemum vulgare) in Kosciuszko National Park, New South Wales. Ecol. Manag. Restor. 2018, 19, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Khuroo, A.A.; Malik, A.H.; Reshi, Z.A.; Dar, G.H. From ornamental to detrimental: Plant invasion of Leucanthemum vulgare Lam. (Ox-eye Daisy) in Kashmir valley, India. Curr. Sci. 2010, 98, 600–602. [Google Scholar]
- Ahmad, R.; Khuroo, A.A.; Charles, B.; Hamid, M.; Rashid, I.; Aravind, N. Global distribution modelling, invasion risk assessment and niche dynamics of Leucanthemum vulgare (Ox-eye Daisy) under climate change. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, B.E.; Wallander, R.T. Oxeye daisy (Chrysanthemum leucanthemum L.). In Biology and Management of Noxious Rangeland Weeds; Sheley, R.L., Petroff, J.K., Eds.; Oregon State University Press: Corvallis, OR, USA, 1999; pp. 282–289. [Google Scholar]
- McConnachie, A.J.; Peach, E.; Turner, P.J.; Stutz, S.; Schaffner, U.; Simmons, A. The invasive weed ox-eye daisy, Leucanthemum vulgare Lam. (Asteraceae): Prospects for its management in New South Wales. Plant Prot. Q. 2015, 30, 103. [Google Scholar]
- McClay, A.S.; Stutz, S.; Schaffner, U. Leucanthemum vulgare Lam., Oxeye Daisy (Asteraceae). In Biological Control Programmes in Canada 2001–2012; Mason, P., Gillespie, D., Eds.; CABI: Wallingford, UK, 2013; pp. 337–342. [Google Scholar]
- Greuter, W. Compositae (Pro Parte Majore). Euro+Med Plantbase: The Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 1 March 2021).
- Fernald, M.L. Chrysanthemum leucanthemum and the American white weed. Rhodora 1903, 5, 177–181. [Google Scholar]
- Stutz, S.; Štajerová, K.; Hinz, H.L.; Müller-Schärer, H.; Schaffner, U. Can enemy release explain the invasion success of the diploid Leucanthemum vulgare in North America? Biol. Invasions 2016, 18, 2077–2091. [Google Scholar] [CrossRef] [Green Version]
- Mulligan, G.A. Chromosome races in the Chrysanthemum leucanthemum complex. Rhodora 1958, 60, 122–125. [Google Scholar]
- Thompson, I.R. A taxonomic treatment of tribe Anthemideae (Asteraceae) in Australia. Muelleria 2007, 25, 21–58. [Google Scholar]
- Oberprieler, C.; Himmelreich, S.; Källersjö, M.; Vallès, J.; Watson, L.E.; Vogt, R. Anthemideae. In Systematics, Evolution, and Biogeography of the Compositae; Funk, V., Susanna, A., Stuessy, T., Bayer, R., Eds.; IAPT: Vienna, Austria, 2009; pp. 631–666. [Google Scholar]
- Stutz, S.; Hinz, H.L.; Schaffner, U. Evaluation of Cyphocleonus trisulcatus (Coleoptera: Curculionidae) as a potential biological control agent for Leucanthemum vulgare in North America. J. Appl. Entomol. 2020, 144, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Tahara, M. Cytologische Studien an einigen Kompositen. J. Coll. Sci. Imp. Univ. Tokyo Jpn. 1921, 43, 1–53. [Google Scholar]
- Hawke, R.G. A Report on Leucanthemum × superbum and Related Daisies; Chicago Botanic Garden: Chicago, IL, USA, 2007; Volume 30, pp. 1–4. [Google Scholar]
- Anderson, N.O.; Olsen, R.T. A vast array of beauty: The accomplishments of the father of American ornamental breeding, Luther Burbank. HortScience 2015, 50, 161–188. [Google Scholar] [CrossRef] [Green Version]
- Bland, K.P.; Razowski, J.; Hancock, E.F. The Moths and Butterflies of Great Britain and Ireland; Tortricidae, Olethreutinae Brill: Leiden, The Netherlands, 2015; Volume 5, Part 2. [Google Scholar]
- Razowski, J. Tortricidae (Lepidoptera) of Europe: Olethreutinae; František Slamka: Bratislava, Slovakia, 2003; Volume 2. [Google Scholar]
- Stutz, S.; Hinz, H.L.; Konowalik, K.; Müller-Schärer, H.; Oberprieler, C.; Schaffner, U. Ploidy level in the genus Leucanthemum correlates with resistance to a specialist herbivore. Ecosphere 2016, 7, e01460. [Google Scholar] [CrossRef]
- Sabourin, M. First report of two palearctic Dichrorampha (Lepidoptera: Tortricidae) species for Vermont. VES News Newsl. Vt. Entomol. Soc. 2009, 64, 9–10. [Google Scholar]
- Hebert, P.D.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; deWaard, J.R. Counting animal species with DNA barcodes: Canadian insects. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaffner, U. Host range testing of insects for biological weed control: How can it be better interpreted? Bioscience 2001, 51, 951–959. [Google Scholar] [CrossRef] [Green Version]
- Briese, D.T. Open field host-specificity tests: Is “natural” good enough for risk assessment? In Proceedings of the Host specificity testing in Australasia: Towards Improved Assays for Biological Control, Paper from the Introduction of Exotic Biocontrol Agents-Recommendations on Host Specificity Testing Procedures in Australasia Workshop, Brisbane, Australia, 29 September–2 October 1998. [Google Scholar]
- Clement, S.L.; Cristofaro, M. Open-field tests in host-specificity determination of insects for biological control of weeds. Biocontrol Sci. Technol. 1995, 5, 395–406. [Google Scholar] [CrossRef]
- Schaffner, U.; Smith, L.; Cristofaro, M. A review of open-field host range testing to evaluate non-target use by herbivorous biological control candidates. BioControl 2018, 63, 405–416. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Buckley, Y.M.; Ventim, R.; Fowler, S.V.; Memmott, J. Apparent competition can compromise the safety of highly specific biocontrol agents. Ecol. Lett. 2008, 11, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Pearson, D.E.; Callaway, R.M. Indirect nontarget effects of host-specific biological control agents: Implications for biological control. Biol. Control 2005, 35, 288–298. [Google Scholar] [CrossRef]
- Sheppard, A.W. The interaction between natural enemies and interspecific plant competition in the control of invasive pasture weeds. In Proceedings of the IX International Symposium on Biological Control of Weeds, Stellenbosch, South Africa, 19–26 January 1996; pp. 47–53. [Google Scholar]
- Cole, D.E. Effect of Competition on Growth of ox-Eye Daisy (Chrysanthemum leucanthemum L.) in Pastures and Hay Land. Master’s. Thesis, University of Alberta, Edmonton, Alberta, 1998. [Google Scholar]
- Wapshere, A.J. A strategy for evaluating the safety of organisms for biological weed control. Ann. Appl. Biol. 1974, 77, 201–211. [Google Scholar] [CrossRef]
- Briese, D.T. The centrifugal phylogenetic method used to select plants for host-specificity testing of weed biological control agents: Can and should it be modernised? In Improving the Selection, Testing, and Evaluation of Weed Biological Control Agents; Spafford Jacob, H., Briese, D., Eds.; CRC for Australian Weed Management: Glen Osmond, Australia, 2003; pp. 23–33. [Google Scholar]
- Kelch, D.G.; McClay, A. Putting the phylogeny into the centrifugal phylogenetic method. In Proceedings of the XI International Symposium on Biological Control of Weeds, Canberra, Australia, 27 April–2 May 2003; pp. 287–291. [Google Scholar]
- Wapshere, A.J. A testing sequence for reducing rejection of potential biological control agents for weeds. Ann. Appl. Biol. 1989, 114, 515–526. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org (accessed on 20 January 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Pemberton, R.W. Predictable risk to native plants in weed biological control. Oecologia 2000, 125, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Marohasy, J. The design and interpretation of host-specificity tests for weed biological control with particular reference to insect behaviour. Biocontrol News Inf. 1998, 19, 13N–20N. [Google Scholar]
- Turanli, F.; Schaffner, U. Oviposition specificity of the specialist Tinthia myrmosaeformis under different degrees of behavioral restrictions. Biol. Control 2004, 30, 274–280. [Google Scholar] [CrossRef]
- Catton, H.A.; Lalonde, R.G.; De Clerck-Floate, R.A. Differential host-finding abilities by a weed biocontrol insect create within-patch spatial refuges for nontarget plants. Environ. Entomol. 2014, 43, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Lefoe, G.; Haegi, L.; Rumpff, L.; Gopurenko, D.; Slater, A.T.; Butler, K.; Hauser, C.E. Assessing the fundamental host-range of Leptinotarsa texana Schaeffer as an essential precursor to biological control risk analysis. Biol. Control 2020, 143, 104165. [Google Scholar] [CrossRef]
- McConnachie, A.J. Host range and risk assessment of Zygogramma bicolorata, a defoliating agent released in South Africa for the biological control of Parthenium hysterophorus. Biocontrol Sci. Technol. 2015, 25, 975–991. [Google Scholar] [CrossRef]
- Wild, C.; McFadyen, R.; Tomley, A.; Willson, B. The biology and host specificity of the stem-boring weevil Listronotus setosipennis [Col.: Curculionidae] A potential biocontrol agent for Parthenium hysterophorus [Asteraceae]. Entomophaga 1992, 37, 591–598. [Google Scholar] [CrossRef]
- Hogg, B.N.; Moran, P.J.; Smith, L. Relative performance and impacts of the psyllid Arytinnis hakani (Hemiptera: Psyllidae) on nontarget plants and the target weed Genista monspessulana (Fabales: Fabaceae). Environ. Entomol. 2019, 48, 524–532. [Google Scholar] [CrossRef]
- Willis, A.J.; Ash, J.E.; Groves, R.H. Combined effects of two arthropod herbivores and water stress on growth of Hypericum species. Oecologia 1993, 96, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Willis, A.; Ash, J.; Groves, R. The effects of herbivory by a mite, Aculus hyperici, and nutrient deficiency on growth in Hypericum species. Aust. J. Bot. 1995, 43, 305–316. [Google Scholar] [CrossRef]
- Willis, A.J.; Berentson, P.R.; Ash, J.E. Impacts of a weed biocontrol agent on recovery from water stress in a target and a non-target Hypericum species. J. Appl. Ecol. 2003, 40, 320–333. [Google Scholar] [CrossRef]
- Shabbir, A.; Dhileepan, K.; Zalucki, M.P.; O’Donnell, C.; Khan, N.; Hanif, Z.; Adkins, S.W. The combined effect of biological control with plant competition on the management of parthenium weed (Parthenium hysterophorus L.). Pak. J. Bot. 2015, 47, 157–159. [Google Scholar]
- Shabbir, A.; Dhileepan, K.; O’Donnell, C.; Adkins, S.W. Complementing biological control with plant suppression: Implications for improved management of parthenium weed (Parthenium hysterophorus L.). Biol. Control 2013, 64, 270–275. [Google Scholar] [CrossRef]
- Dube, N.; Uyi, O.; Zachariades, C.; Munyai, T.C.; Whitwell, M. Impact of the shoot-boring moth Dichrorampha odorata (Lepidoptera: Tortricidae) on growth and reproductive potential of Chromolaena odorata (Asteraceae) in the laboratory. Biocontrol Sci. Technol. 2019, 29, 350–364. [Google Scholar] [CrossRef]
- Zachariades, C.; Dube, N.; Nqayi, S.; Dlomo, S.; Uyi, O. Attempts to establish Dichrorampha odorata on Chromolaena odorata in South Africa. In Proceedings of the XV International Symposium on Biological Control of Weeds, Engelberg, Switzerland, 26–31 August 2018; pp. 272–274. [Google Scholar]
- Fogliani, R.; Strickland, G. Biological Control of Dock: Enhanced Distribution of the Dock Moth; Final report to Meat and Livestock Australia; Agriculture Western Australia: North Sydney, Australia, 2000. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Plants Dissected for Larvae | Plants Kept for Adult Emergence or with Larvae Transferred Back on Same Plant Species | |||||
|---|---|---|---|---|---|---|---|
| No. Plants | % Plants with Larvae | % Larvae Recovered/Plant (Mean ± SE) | No. Plants | % Plants with Adult Emergence | % Adults Emerged/Plant (Mean ± SE) | ||
| Family Asteraceae | |||||||
| Tribe Anthemideae | |||||||
| Subtribe Leucantheminae | |||||||
| Leucanthemum vulgare (Vaill.) Lam. (tests at CABI) | 242 | 90.9 | 51.0 ± 1.9 | 59 | 81.4 | 36.4 ± 3.3 | |
| Leucanthemum vulgare (tests at AAFC) | 12 | 83.3 | 30.3 ± 0.6 | ||||
| Leucanthemum vulgare (tests at NSW DPI) | 15 | 100.0 | 73.3 ± 4.7 | ||||
| Leucanthemum ircutianum (Turcz.) Turcz. ex DC. | 141 | 85.8 | 44.5 ± 2.4 | 35 | 91.4 | 37.5 ± 4.2 | |
| Leucanthemum × superbum (Bergmans ex J. Ingram) Kent ‘Alaska’ | 14 | 35.7 | 7.1 ± 2.7 | 10 | 10.0 | 2.0 ± 2.0 | |
| Leucanthemum × superbum ‘Amelia’ | 15 | 73.3 | 30.7 ± 7.0 | 6 | 33.3 | 6.7 ± 4.2 | |
| Leucanthemum × superbum ‘Becky’ | 8 | 12.5 | 2.5 ± 1.3 | ||||
| Leucanthemum × superbum ‘Crazy Daisy’ | 11 | 63.6 | 20.0 ± 7.6 | ||||
| Leucanthemum × superbum ‘Goldrush’ | 7 | 71.4 | 22.9 ± 8.1 | ||||
| Leucanthemum × superbum ‘Marconi Double’ | 5 | 20.0 | 4.0 ± 4.0 | 2 | 50.0 | 20.0 ± 20.0 | |
| Leucanthemum × superbum ‘Silver Princess’ | 10 | 80.0 | 20.0 ± 5.2 | 3 | 0.0 | ||
| Leucanthemum × superbum ‘Snow Lady’ | 4 | 25.5 | 5.0 ± 5.0 | 2 | 0.0 | ||
| Leucanthemum× superbum ‘unnamed 1’ (from B & T Seeds) | 10 | 50.0 | 14.4 ± 5.2 | 8 | 37.5 | 7.5 ± 3.7 | |
| Leucanthemum× superbum ‘unnamed 2’ (from Eden Seeds) | 10 | 70.0 | 17.5 ± 5.5 | ||||
| Leucanthemum× superbum ‘unnamed 3’ (from Australian Seed co.) | 10 | 80.0 | 18.0 ± 3.6 | ||||
| Mauranthemum paludosum (Poir.) Vogt & Oberpr. | 7 | 28.6 | 5.7 ± 3.7 | ||||
| Subtribe Anthemidinae | |||||||
| Anthemis arvensis L. | 11 | 0.0 | |||||
| Anthemis cotula L. | 13 | 30.8 | 7.7 ± 5.1 | 10 | 10.0 d | 10.0 ± 10.0 d | |
| Anthemis tinctoria L. | 7 | 0.0 | |||||
| Tanacetum camphoratum Less. a | 7 | 0.0 | |||||
| Tanacetum cinerariifolium (Trevir.) Sch.Bip. | 7 | 0.0 | |||||
| Tanacetum huronense Nutt. a | 7 | 0.0 | |||||
| Tanacetum parthenium (L.) Sch.Bip. | 4 | 50.0 | 15.0 ± 9.6 | 5 | 20.0 | 4.0 ± 4.0 | |
| Tanacetum vulgare L. | 7 | 0.0 | |||||
| Tripleurospermum inodorum (L.) Sch.Bip. | 7 | 0.0 | |||||
| Subtribe Artemisiinae | |||||||
| Arctanthemum arcticum (L.) Tzvelev (ornamental) | 7 | 0.0 | |||||
| Arctanthemum arcticum a | 9 | 0.0 | |||||
| Artemisia absinthium L. | 9 | 0.0 | |||||
| Artemisia arctica Less. a | 2 | 0.0 | |||||
| Artemisia biennis Willd a | 7 | 0.0 | |||||
| Artemisia californica Less. a | 7 | 0.0 | |||||
| Artemisia campestris L. a | 7 | 0.0 | |||||
| Artemisia cana Pursh a | 7 | 0.0 | |||||
| Artemisia dracunculus L. a | 7 | 0.0 | |||||
| Artemisia filifolia Torr. a | 10 | 0.0 | |||||
| Artemisia frigida Willd. a | 7 | 0.0 | |||||
| Artemisia ludoviciana Nutt. a | 7 | 0.0 | |||||
| Artemisia scopulorum A. Gray a | 7 | 0.0 | |||||
| Artemisia spinescens D.C.Eaton a | 8 | 0.0 | |||||
| Artemisia tridentata Nutt. a | 7 | 0.0 | |||||
| Artemisia vulgaris L. a | 7 | 0.0 | |||||
| Chrysanthemum × grandiflorum (Ramat.) Kitam. ‘Garden Mums’ | 9 | 0.0 | |||||
| Chrysanthemum × grandiflorum ‘Morden Canary’ | 7 | 0.0 | |||||
| Chrysanthemum × grandiflorum ‘Morden Delight’ | 7 | 0.0 | |||||
| Chrysanthemum × indicum L. | 5 | 0.0 | |||||
| Hulteniella integrifolia (Richardson) a | 12 | 0.0 | |||||
| Leucanthemella serotina (L.) Tzvelev | 12 | 25.0 | 5.0 ± 2.6 | 4 | 25.0 e | 25.0 ± 25.0 e | |
| Subtribe Cotulinae | |||||||
| Cotula alpina (Hook.f.) Hook.f. b | 8 | 0.0 | |||||
| Cotula australis (Sieber ex Spreng.) Hook.f b | 17 | 0.0 | |||||
| Cotula coronopifolia L. | 7 | 0.0 | |||||
| Cotula cotuloides (Steetz) Druce b | 8 | 16.7 | 3.3 ± 3.3 | ||||
| Cotula vulgaris Levyns var. vulgaris c | 11 | 9.1 | 1.8 ± 1.8 | ||||
| Leptinella filicula (Hook.f.) Hook.f b | 14 | 0.0 | |||||
| Leptinella longipes Hook.f. b | 12 | 0.0 | |||||
| Leptinella reptans D.G.Lloyd & C.J.Webb b | 7 | 0.0 | |||||
| Subtribe Glebionidinae | |||||||
| Argyranthemum frutescens (L.) Sch.Bip. | 13 | 0.0 | |||||
| Glebionis coronaria (L.) Cass. ex Spach | 6 | 0.0 | |||||
| Glebionis segetum (L.) Fourr. | 7 | 0.0 | |||||
| Ismelia carinata (Schousb.) Sch.Bip. | 15 | 6.7 | 1.3 ± 1.3 | ||||
| Subtribe Matricariinae | |||||||
| Achillea alpina L. a | 14 | 0.0 | |||||
| Achillea borealis Bong. a | 10 | 0.0 | |||||
| Achillea ptarmica L. | 23 | 4.4 f | 0.9 ± 0.9 f | ||||
| Matricaria chamomilla L. | 24 | 8.3 | 1.7 ± 1.2 | ||||
| Matricaria discoidea DC. a | 18 | 16.7 | 3.3 ± 1.8 | ||||
| Matricaria occidentalis Greene a | 15 | 13.3 | 2.6 ± 1.8 | 13 | 0.0 | ||
| Subtribe Santolininae | |||||||
| Chamaemelum nobile (L.) All. | 7 | 0.0 | |||||
| Santolina chamaecyparissus L. | 7 | 0.0 | |||||
| Tribe Astereae | |||||||
| Brachyscome aculeata (Labill.) Less. b | 16 | 0.0 | |||||
| Brachyscome multifida DC. b | 5 | 0.0 | |||||
| Calotis pubescens (F.Muell. ex Benth.) N.G.Walsh & K.L. McDougall b | 12 | 0.0 | |||||
| Erigeron compositus Pursh a | 8 | 0.0 | |||||
| Solidago nemoralis Aiton a | 9 | 0.0 | |||||
| Symphyotrichum laeve (L.) Á.Löve & D.Lövea | 7 | 0.0 | |||||
| Tribe Calenduleae | |||||||
| Osteospermum ecklonis (DC.) Norl. | 5 | 0.0 | |||||
| Tribe Cardueae | |||||||
| Carthamus tinctorius L. | 7 | 0.0 | |||||
| Cirsium flodmanii (Rydb.) Arthur a | 6 | 0.0 | |||||
| Cynara scolymus L. | 7 | 0.0 | |||||
| Tribe Coreopsideae | |||||||
| Coreopsis tinctoria Nutt. a | 7 | 0.0 | |||||
| Tribe Eupatorieae | |||||||
| Eutrochium maculatum (L.) E.E.Lamont a | 7 | 0.0 | |||||
| Tribe Gnaphalieae | |||||||
| Anaphalis margaritacea (L.) Benth. a | 7 | 0.0 | |||||
| Tribe Helenieae | |||||||
| Helenium autumnale L. a | 7 | 0.0 | |||||
| Tribe Heliantheae | |||||||
| Helianthus annuus L. a | 14 | 0.0 | |||||
| Echinacea purpurea (L.) Moench | 8 | 0.0 | |||||
| Tribe Lactuceae | |||||||
| Cichorium intybus L. | 7 | 0.0 | |||||
| Lactuca sativa L. | 15 | 0.0 | |||||
| Tribe Madieae | |||||||
| Arnica chamissonis Less. a | 9 | 0.0 | |||||
| Tribe Senecioneae | |||||||
| Senecio eremophilus Richardson a | 7 | 0.0 | |||||
| Tribe Tageteae | |||||||
| Tagetes lucida Cav. | 7 | 0.0 | |||||
| Family Apiaceae | |||||||
| Daucus carota L. | 10 | 0.0 | |||||
| Petroselinum crispum (Mill.) Fuss | 7 | 0.0 | |||||
| Family Campanulaceae | |||||||
| Lobelia cardinalis L. a | 7 | 0.0 | |||||
| Plant Species/Cultivar | No. Plants | % Plants with Larvae | % Plants with Eggs or Larvae | No. Eggs/Plant (Mean ± SE) | No. Larvae/Plant (Mean ± SE) | No. Larvae with Weight and/or Instar Measured | Larval Weight (mg) (Mean ± SE) | % 3rd Instar | % 4th Instar | |
|---|---|---|---|---|---|---|---|---|---|---|
| 2012 (6 cages) | ||||||||||
| Leucanthemum vulgare (rosettes) | 18 | 77.8 | 94.4 | 8.2 ± 2.4 | 5.8 ± 1.3 | 69 | 2.1 ± 0.1 | 43.5 | 56.5 | |
| L. vulgare (with stems) | 9 | 100.0 | 100.0 | 16.9 ± 3.1 | 85 | 2.2 ± 0.1 | 43.5 | 56.5 | ||
| L. ircutianum | 18 | 88.9 | 94.4 | 6.2 ± 1.3 | 6.9 ± 1.2 | 73 | 2.3 ± 0.1 | 43.8 | 56.2 | |
| L. × superbum ‘Alaska’ | 18 | 33.3 | 50.0 | 5.8 ± 2.7 | 0.6 ± 0.2 | 8 | 1.3 ± 0.1 | 87.5 | 12.5 | |
| L. × superbum ‘Amelia’ | 18 | 38.9 | 61.1 | 2.7 ± 0.7 | 0.9 ± 0.3 | 11 | 1.4 ± 0.1 | 90.9 | 9.1 | |
| L. × superbum ‘unnamed 1’ | 18 | 20.0 | 33.3 | 1.3 ± 0.7 | 0.3 ± 0.1 | 2 | 1.0 ± 0.1 | 100.0 | 0.0 | |
| 2013 (5 cages) | ||||||||||
| L. vulgare | 15 | 100.0 | 10.3 ± 1.1 | 26 | 1.9 ± 0.2 | 73.1 | 26.9 | |||
| L. ircutianum | 14 | 92.9 | 6.4 ± 1.5 | 18 | 2.2 ± 0.2 | 66.7 | 33.3 | |||
| Anthemis cotula | 15 | 0.0 | 0.0 | |||||||
| Matricaria occidentalis a | 15 | 6.7 | 0.07 ± 0.07 | 1 | 3.2 | 0.0 | 100.0 | |||
| Leucanthemella serotina | 15 | 6.7 | 0.07 ± 0.07 | |||||||
| 2016 (4 cages) b | ||||||||||
| L. vulgare | 6 | 33.3 | 2.7 ± 1.7 | 4 | 1.9 ± 0.1 | 25.0 | 75.0 | |||
| Achillea ptarmica c | 3 | 0.0 | 0.0 | |||||||
| Ismelia carinata | 6 | 16.7 | 0.2 ± 0.2 | 1 | 1.6 | 0.0 | 100.0 | |||
| Matricaria chamomilla | 6 | 0.0 | 0.0 | |||||||
| Matricaria discoidea a | 6 | 0.0 | 0.0 | |||||||
| 2017 (4 cages) | ||||||||||
| L. vulgare | 12 | 66.7 | 3.4 ± 1.1 | 9 | 2.2 ± 0.2 | 33.3 | 66.7 | |||
| Ismelia carinata | 12 | 0.0 | 0.0 | |||||||
| Matricaria chamomilla | 12 | 0.0 | 0.0 | |||||||
| Matricaria discoidea a | 12 | 0.0 | 0.0 | |||||||
| 2020 (6 cages) | ||||||||||
| L. vulgare | 18 | 94.4 | 5.8 ± 1.4 | 65 | 7.7 e | 78.5 e | ||||
| Cotula cotuloides d | 14 | 0.0 | 0.0 | |||||||
| Mauranthemum paludosum | 16 | 25.0 | 0.4 ± 0.2 | 5 | 0.0 | 100 | ||||
| Tanacetum parthenium | 18 | 0.0 | 0.0 | |||||||
| Year | Plant Species/Cultivar | No. Plants | % Plants with Larvae | No. Larvae/Plant (Mean ± SE) | No. Larvae with Weight and/or Instar Measured | Larval Weight (mg) (Mean ± SE) | % 3rd Instar | % 4th Instar | % 5th Instar |
|---|---|---|---|---|---|---|---|---|---|
| 2013 (design as in Figure 1a) | |||||||||
| Leucanthemum vulgare (rosettes) | 16 | 100.0 | 5.8 ± 0.7 | 26 | 5.4 ± 0.4 | 0.0 | 19.2 | 80.8 | |
| L. vulgare (with stems) | 16 | 93.8 | 3.8 ± 0.6 | 12 | 4.3 ± 0.4 | 8.3 | 58.3 | 33.3 | |
| L. × superbum ‘Alaska’ | 12 | 8.3 | 0.2 ± 0.2 | 1 | 0.6 | 100.0 | 0.0 | 0.0 | |
| L. × superbum ‘Amelia’ | 12 | 83.3 | 1.8 ± 0.4 | 11 | 4.8 ± 0.4 | 0.0 | 36.4 | 63.6 | |
| L. × superbum ‘unnamed 1’ | 12 | 25.0 | 0.3 ± 0.1 | 1 | 3.5 | 0.0 | 0.0 | 100.0 | |
| 2014 (design as in Figure 1b) | |||||||||
| L. vulgare (rosettes) | 15 | 100.0 | 9.1 ± 1.5 | 10 | 5.0 ± 0.6 | 0.0 | 40.0 | 60.0 | |
| L. vulgare (with stems) | 2 | 50.0 | 0.5 ± 0.5 | ||||||
| L. × superbum ‘Alaska’ | 10 | 40.0 | 0.8 ± 0.4 | 6 | 4.0 ± 0.8 | 0.0 | 50.0 | 50.0 | |
| L. × superbum ‘Becky’ | 10 | 20.0 | 0.2 ± 0.1 | 1 | 2.6 | 0.0 | 100.0 | 0.0 | |
| L. × superbum ‘Crazy Daisy’ | 8 | 12.5 | 0.1 ± 0.1 | ||||||
| L. × superbum ‘unnamed 4’ | 10 | 40.0 | 0.6 ± 0.3 | 1 | 4.2 | 0.0 | 0.0 | 100.0 | |
| 2014c (design as in Figure 1c) | |||||||||
| L. vulgare (rosettes) | 9 | 100.0 | 14.4 ± 2.4 | 12 | 3.1 ± 0.3 | 0.0 | 100.0 | 0.0 | |
| L. vulgare (with stems) | 7 | 71.4 | 2.3 ± 2.0 | ||||||
| L. × superbum ‘Silver Princess’ | 8 | 50.0 | 1.6 ± 0.9 | 4 | 2.0 ± 0.2 | 75.0 | 25.0 | 0.0 | |
| Achillea ptarmica a | 9 | 0.0 | 0.0 | ||||||
| Anthemis cotula | 7 | 0.0 | 0.0 | ||||||
| Matricaria occidentalis b | 8 | 0.0 | 0.0 | ||||||
| Leucanthemella serotina | 9 | 0.0 | 0.0 | ||||||
| 2017 (design as in Figure 1c) | |||||||||
| L. vulgare (rosettes) | 16 | 93.8 | 4.6 ± 0.8 | 12 | 2.5 ± 0.2 | 41.7 | 58.3 | 0.0 | |
| Ismelia carinata | 13 | 0 | 0.0 | ||||||
| Matricaria chamomilla | 16 | 0 | 0.0 | ||||||
| Matricaria discoidea b | 16 | 0 | 0.0 | ||||||
| Matricaria occidentalis b | 15 | 0 | 0.0 | ||||||
| 2019 (design as in Figure 1c) | |||||||||
| L. vulgare (rosettes) | 15 | 80.0 | 2.9 ± 0.7 | 4 | 50.0 | 50.0 | 0.0 | ||
| Cotula cotuloides c | 8 | 0.0 | 0.0 | ||||||
| Mauranthemum paludosum | 16 | 6.3 | 0.1 ± 0.1 | 2 | 50.0 | 50.0 | 0.0 | ||
| Tanacetum parthenium | 16 | 0.0 | 0.0 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stutz, S.; De Clerck-Floate, R.; Hinz, H.L.; McClay, A.; McConnachie, A.J.; Schaffner, U. Host Range and Impact of Dichrorampha aeratana, the First Potential Biological Control Agent for Leucanthemum vulgare in North America and Australia. Insects 2021, 12, 438. https://doi.org/10.3390/insects12050438
Stutz S, De Clerck-Floate R, Hinz HL, McClay A, McConnachie AJ, Schaffner U. Host Range and Impact of Dichrorampha aeratana, the First Potential Biological Control Agent for Leucanthemum vulgare in North America and Australia. Insects. 2021; 12(5):438. https://doi.org/10.3390/insects12050438
Chicago/Turabian StyleStutz, Sonja, Rosemarie De Clerck-Floate, Hariet L. Hinz, Alec McClay, Andrew J. McConnachie, and Urs Schaffner. 2021. "Host Range and Impact of Dichrorampha aeratana, the First Potential Biological Control Agent for Leucanthemum vulgare in North America and Australia" Insects 12, no. 5: 438. https://doi.org/10.3390/insects12050438
APA StyleStutz, S., De Clerck-Floate, R., Hinz, H. L., McClay, A., McConnachie, A. J., & Schaffner, U. (2021). Host Range and Impact of Dichrorampha aeratana, the First Potential Biological Control Agent for Leucanthemum vulgare in North America and Australia. Insects, 12(5), 438. https://doi.org/10.3390/insects12050438