
Diel Rhythmicity of Field Responses to Synthetic Pheromone Lures in the Pine Sawyer Monochamus saltuarius


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Period
2.2. Periodicity of the Response of M. saltuarius to Pheromone
2.2.1. Pheromone Source
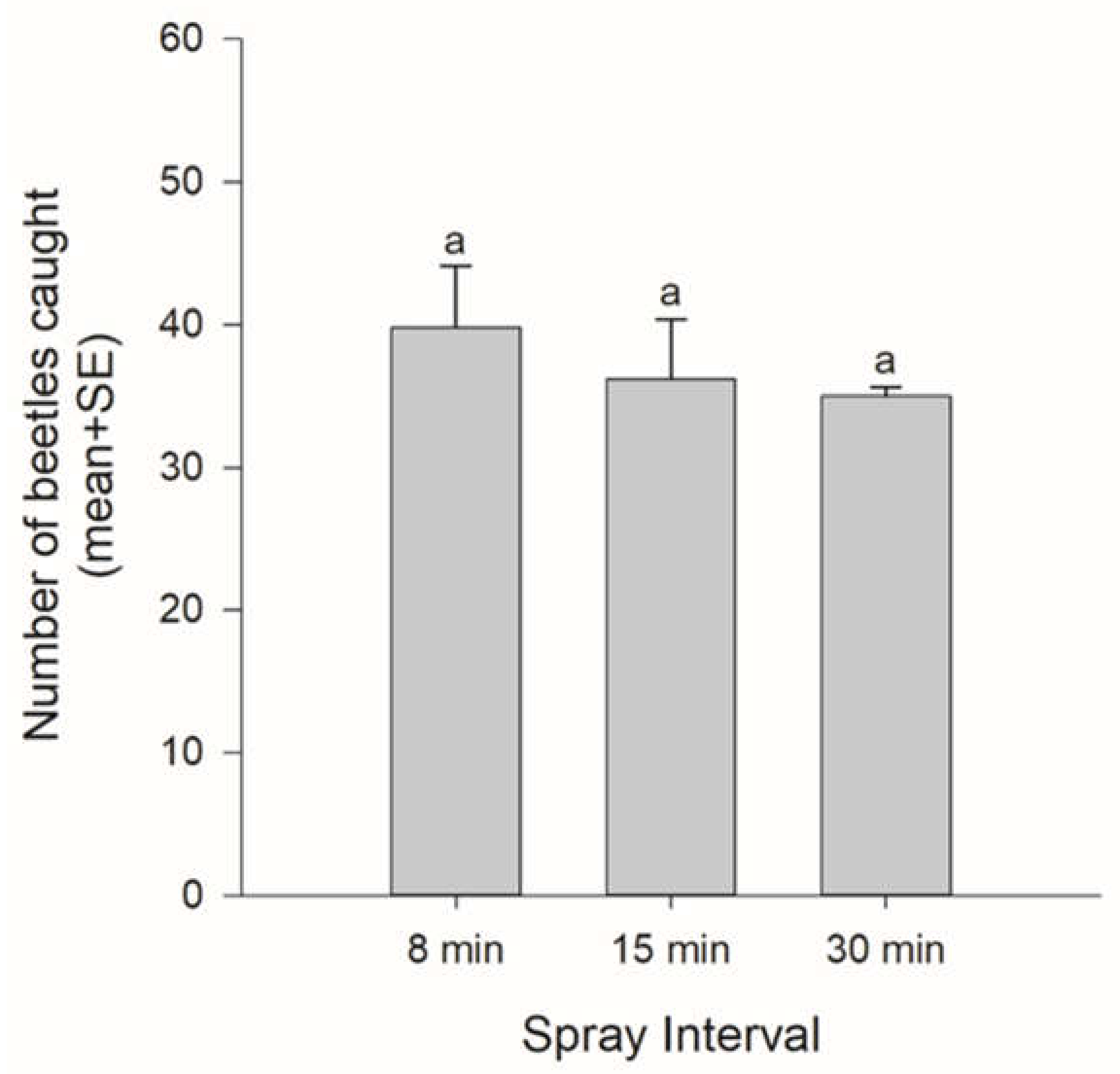
2.2.2. Effect of PA Concentration on Attraction (Emission Interval 8, 15, 30 min)
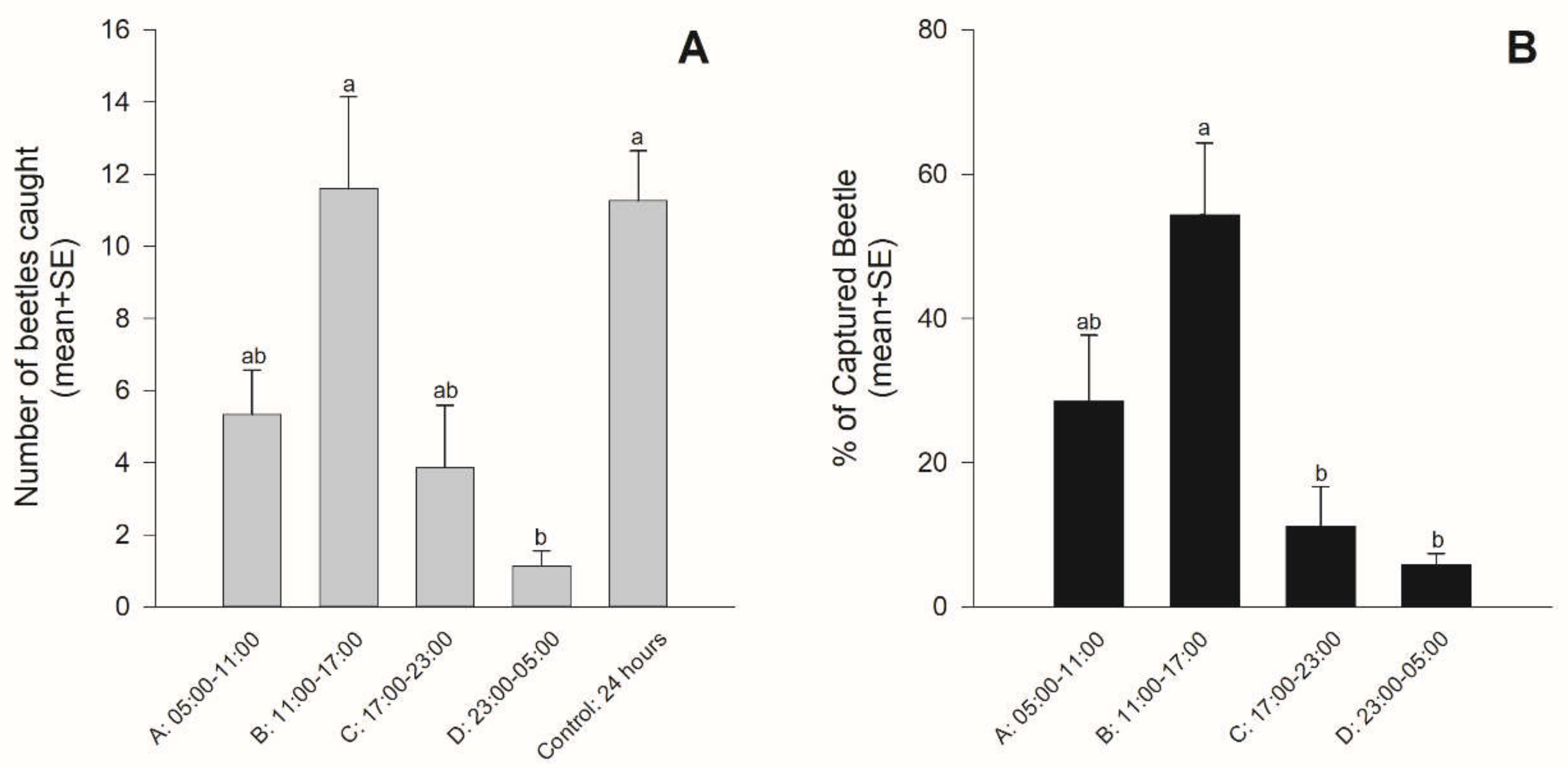
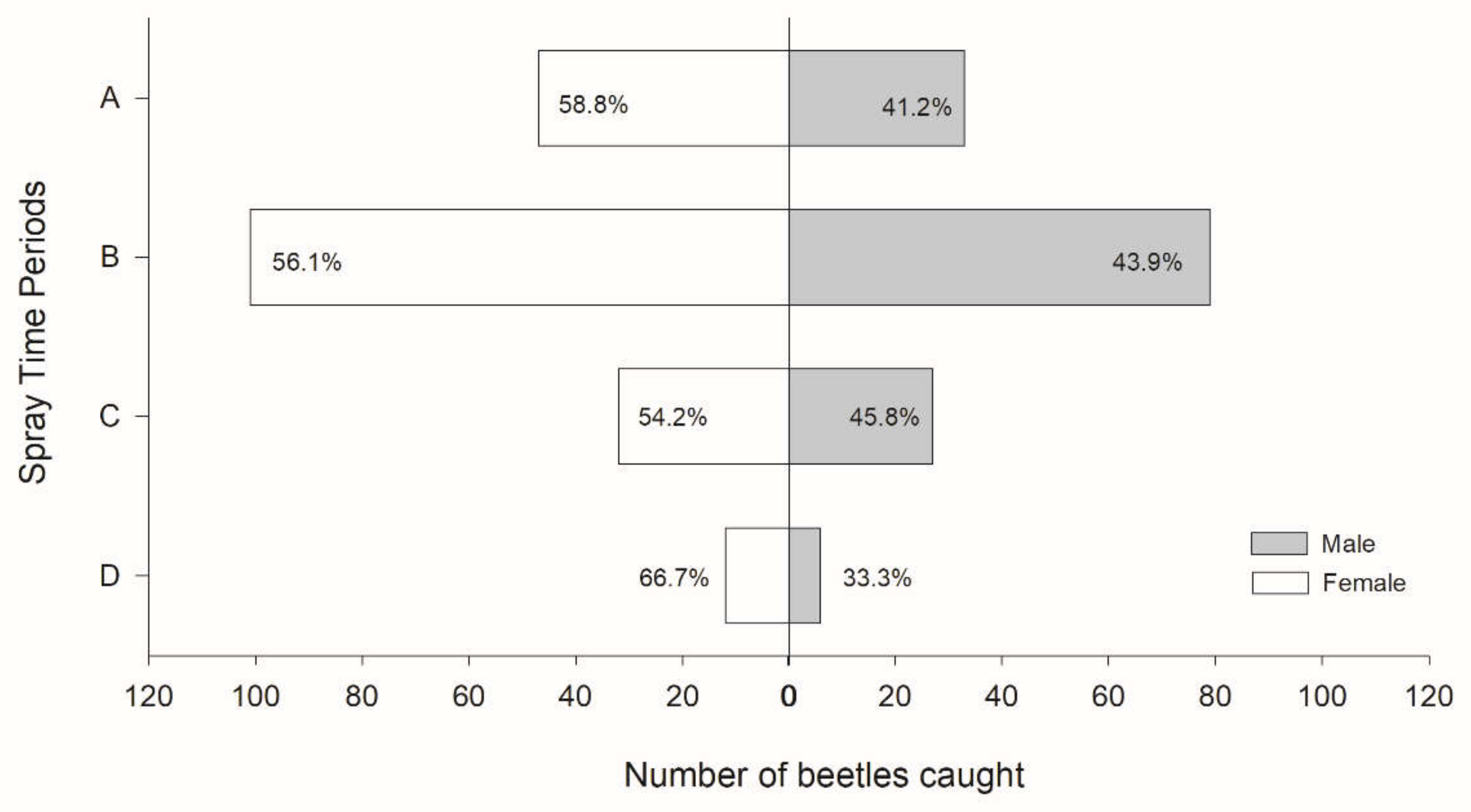
2.2.3. The Diel Rhythm of Responses to Aggregation PA
2.3. Statistical Analysis
3. Results
3.1. Effect of PA Concentration on Attraction
3.2. The Diel Rhythm of Response to Aggregation PA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, B.; Futai, K.; Sutherland, J.R.; Takeuchi, Y. Pine Wilt Disease; Springer: Tokyo, Japan, 2008. [Google Scholar]
- Sato, H.; Sakuyama, T.; Kobayashi, M. Transmission of Bursaphelenchus xylophilus (STEINER et BUHRER) NICKLE (Nematoda, Aphelenchoididae) by Monochamus saltuarius (GEBLER) (Coleoptera, Cerambycidae). J. Jpn. For. Soc. 1987, 69, 492–496. [Google Scholar]
- Zhao, B.G. Pine wilt disease in China. In Pine Wilt Disease; Zhao, B., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 18–25. [Google Scholar]
- Shin, S.-C. Pine wilt disease in Korea. In Pine Wilt Disease; Zhao, B., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 26–32. [Google Scholar]
- Pajares, J.A.; Alvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.R.; Farman, D.I. Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Korea Forest Research Institute. Annual Report of Monitoring for Forest Insect Pest and Diseases in Korea; Korea Forest Research Institute: Seoul, Korea, 2007; p. 259. [Google Scholar]
- Potenko, V.V.; Velikov, A.V. Genetic diversity and differentiation of natural populations of Pinus koraiensis (Sieb. et Zucc.) in Russia. Silvae Genet. 1998, 47, 202–208. [Google Scholar]
- Li, M.; Li, H.; Sheng, R.C.; Sun, H.; Sun, S.H.; Chen, F.M. The first record of Monochamus saltuarius (Coleoptera; Cerambycidae) as vector of Bursaphelenchus xylophilus and its new potential hosts in China. Insects 2020, 11, 636. [Google Scholar] [CrossRef]
- Pan, L.; Li, Y.; Cui, R.; Liu, Z.; Zhang, X. Monochamus saltuarius endangers Pinus tabuliformis Carr. and Carries Bursaphelenchus xylophilus (Steiner and Buhrer) in China. Forests 2020, 11, 1051. [Google Scholar] [CrossRef]
- Akbulut, S.; Stamps, W.T. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Han, J.-H.; Kim, H.K.; Kang, W.J.; Kim, G.-H. Feeding and oviposition preference of the Sakhalin pine sawyer Monochamus saltuarius (Coleoptera: Cerambycidae) for various tree species. Entomol. Res. 2016, 46, 331–336. [Google Scholar] [CrossRef]
- Han, J.-H.; Yoon, C.; Shin, S.-C.; Kim, G.-H. Seasonal occurrence and morphological measurements of pine sawyer, Monochamus saltuarius adults (Coleoptera: Cerambycidae). J. Asia Pac. Entomol. 2007, 10, 63–67. [Google Scholar] [CrossRef]
- Kim, M.-K.; Han, J.-H.; Kim, Y.-J.; Yoon, C.; Kim, G.-H. Mating behavior of pine sawyer, Monochamus saltuarius Gebler (Coleoptera: Cerambycidae). J. Asia Pac. Entomol. 2006, 9, 275–280. [Google Scholar] [CrossRef]
- Kwon, T.-S.; Lim, J.-H.; Shin, S.-J.; Kwon, Y.-D.; Son, S.-K.; Lee, K.-Y.; Kim, Y.-T.; Park, J.-W.; Shin, C.-H.; Ryu, S.-B.; et al. Distribution patterns of Monochamus alternatus and M. salturarius (Coleoptera: Cerambycidae) in Korea. J. Korean For. Soc. 2006, 95, 543–550. [Google Scholar]
- Kim, G.-H.; Takabayashi, J.; Takahashi, S.; Tabata, K. Function of pheromones in mating behavior of the Japanese pine sawyer beetle, Monochamus alternatus HOPE. Appl. Entomol. Zool. 1992, 27, 489–497. [Google Scholar] [CrossRef] [Green Version]
- Fauzia, M.K.; Hidaka, T.; Tabata, K. The reproductive behavior of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1987, 22, 272–285. [Google Scholar] [CrossRef]
- Nishimura, M. Daily observation on behaviors of Japanese pine sawyer adult, Monochamus alternatus Hope. J. Jpn. For. Soc. 1973, 55, 100–104. [Google Scholar] [CrossRef]
- Taniwaki, T.; Okitsu, M.; Kishi, Y. Diurnal emergence of Monochamus alternatus Hope (Coleoptera: Cerambycidae) from pine logs. J. Jpn. For. Soc. 2004, 86, 158–163. [Google Scholar]
- Kim, D.S.; Lee, S.M.; Kim, C.S.; Lee, D.W.; Park, C.G. Movement of Monochamus alternatus Hope (Coleoptera: Cerambycidae) adults among young black pine trees in a screen cage. Kor. J. Appl. Entomol. 2011, 50, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.J.; Jung, J.-K.; Jung, C.; Han, H.; Koh, S.-H. Dispersal capacity of Monochamus saltuarius on flight mills. Entomol. Exp. Appl. 2018, 166, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Teale, S.A.; Wickham, J.D.; Zhang, F.; Chen, Y.; Wei, X.; Hanks, L.M.; Millar, J.G. A male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. J. Econ. Entomol. 2011, 104, 1592–1598. [Google Scholar] [CrossRef] [PubMed]
- Allison, J.D.; McKenney, J.L.; Millar, J.G.; McElfresh, J.S.; Mitchell, R.F.; Hanks, L.M. Response of the woodborers Monochamus carolinensis and Monochamus titillator (Coleoptera: Cerambycidae) to known cerambycid pheromones in the presence and absence of the host plant volatile a-pinene. Environ. Entomol. 2012, 41, 1587–1596. [Google Scholar] [CrossRef] [Green Version]
- Pajares, J.A.; Álvarez, G.; Hall, D.R.H.; Douglas, P.; Centeno, F.; Ibarra, N.; Schroeder, M.; Teale, S.A.; Wang, Z.; Yan, S.; et al. 2-(Undecyloxy)-ethanol is a major component of the male-produced aggregation pheromone of Monochamus sutor. Entomol. Exp. Appl. 2013, 149, 118–127. [Google Scholar]
- Lee, H.-R.; Lee, S.-C.; Lee, D.H.; Choi, W.-S.; Jung, C.-S.; Jeon, J.-H.; Kim, J.-E.; Park, I.-K. Identification of the aggregation-sex pheromone produced by male Monochamus saltuarius, a major insect vector of the pine wood nematode. J. Chem. Ecol. 2017, 43, 67–678. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.-M.; Jung, Y.H.; Kwon, Y.-D.; Kim, D.-S.; Lee, D.W.; Park, C.G. Field evaluation on the synergistic attractiveness of 2-(1-undecyloxy)-1-ethanol and ipsenol to Monochamus saltuarius. Entomol. Res. 2016, 46, 31–35. [Google Scholar] [CrossRef]
- Ma, T.; Shi, X.; Shen, J.; Wang, C.; Zhang, S.; Lu, X.; Sun, Z.; Chen, X.; Wang, C.; Xie, W.; et al. Field evaluation of commercial attractants and trap placement for monitoring pine sawyer beetle, Monochamus alternatus (Coleoptera: Cerambycidae) in Guangdong, China. J. Econ. Entomol. 2018, 111, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.R.; Allison, J.D.; Crowe, C.M.; Dickinson, D.M.; Eglitis, A.; Hofstetter, R.W.; Munson, A.S.; Poland, T.M.; Reid, L.S.; Steed, B.E.; et al. Pine sawyers (Coleoptera: Cerambycidae) attracted to α-pinene, monochamol, and ipsenol in North America. J. Econ. Entomol. 2016, 109, 1205–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, M. Trapping strategy for Monochamus sutor and Monochamus galloprovincialis: Potential vectors of the pine wood nematode in Scandinavia. Agric. For. Entomol. 2019, 21, 372–378. [Google Scholar] [CrossRef]
- Boone, C.K.; Sweeney, J.; Silk, P.; Hughes, C.; Webster, R.P.; Stephen, F.; Maclauchlan, L.; Bentz, B.; Drumont, A.; Zhao, B.; et al. Monochamus species from different continents can be effectively detected with the same trapping protocol. J. Pest Sci. 2018, 92, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Skabeikis, D.D.; Teale, S.A.; Fierke, M.K. Diel rhythms in Monochamus (Coleoptera: Cerambycidae): Production of and response to a male-produced aggregation pheromone. Environ. Entomol. 2016, 45, 1017–1021. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Giannone, F.; Moino, S.; Rassati, D. Exploiting trap color to improve surveys of longhorn beetles. J. Pest Sci. 2020. [Google Scholar] [CrossRef]
- Lyu, F.; Hai, X.-X.; Wang, Z.-G. Green-colored paperboard enhances the Asian longhorned beetle response to host plant odor cues. J. Pest Sci. 2021. [Google Scholar] [CrossRef]
- Park, I.K.; Lee, S.C.; Hur, M.J.; Kwon, J.H. Research on Improvement the Attractivenss of Aggregation-Sex Pheromone and Analysis of Behaviour Response; National Instutute of Forest Science: Seoul, Korea, 2018; p. 35. [Google Scholar]
- Kim, J.; Lee, S.-M.; Park, C.G. Bursaphelenchus xylophilus is killed by homologues of 2-(1-undecyloxy)-1-ethanol. Sci. Rep. 2016, 6, 29300. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Enviroment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Baker, T.C.; Dittl, T.; Mafra-Neto, A. Disruption of sex pheromone communication in the blackheaded fireworm in Wisconsin cranberry marshes by using MSTRS™ devices. J. Agric. Entomol. 1997, 14, 449–457. [Google Scholar]
- Baker, T.C.; Mafra-Neto, A.; Dittl, T.; Rice, M.E. A novel controlled-release device for disrupting sex pheromone communication in moths. In Technology Transfer in Mating Disruption IOBC wprs Bull; Witzgall, P., Minks, A.K., Eds.; 1997; Volume 20, pp. 141–149. Available online: https://www.openagrar.de/servlets/MCRFileNodeServlet/openagrar_derivate_00012719/1997_0045.pdf (accessed on 11 January 2021).
- McGhee, P.S.; Miller, J.R.; Thomson, D.R.; Gut, L.J. Optimizing aerosol dispensers for mating dsruption of codling moth, Cydia pomonella L. J. Chem. Ecol. 2016, 42, 612–616. [Google Scholar] [CrossRef]
- Burks, C.S.; Thomson, D.R. Factors affecting disruption of navel orangeworm (Lepidoptera: Pyralidae) using aerosol dispensers. J. Econ. Entomol. 2020, 113, 1190–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelinski, L.; Gut, L.; Haas, M.; McGhee, P.; Epstein, D. Evaluation of aerosol devices for simultaneous disruption of sex pheromone communication in Cydia pomonella and Grapholita molesta (Lepidoptera: Tortricidae). J. Pest Sci. 2007, 80, 225–233. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Reagel, P.F.; Wong, J.C.H.; Meier, L.R.; Silva, W.D.; Mongold-Diers, J.A.; Millar, J.G.; Hanks, L.M. Cerambycid beetle species with similar pheromones are segregated by phenology and minor pheromone components. J. Chem. Ecol. 2015, 41, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Rassati, D.; Marchioro, M.; Flaherty, L.; Poloni, R.; Edwards, S.; Faccoli, M.; Sweeney, J. Response of native and exotic longhorn beetles to common pheromone components provides partial support for the pheromone-free space hypothesis. Insect Sci. 2020. [Google Scholar] [CrossRef]
- Landin, B.-O. The diel flight activity of dung-beetles (Coleoptera Scarabaeidae). A study of the influence of environmental factors, with particular reference to the light. Opusc. Entomol. Suppl. 1968, 32, 1–172. [Google Scholar]
- Feer, F.; Pincebourde, S. Diel flight activity and ecological segregation within an assemblage of tropical forest dung and carrion beetles. J. Trop. Ecol. 2005, 21, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Park, C.G.; Kim, D.S.; Lee, S.M.; Moon, Y.S.; Jin, C.Y.; Kim, D.-S. A forecasting model for the adult emergence of overwintered Monochamus alternatus (Coleoptera: Cerambycidae) larvae based on degree-days in Korea. Appl. Entomol. Zool. 2014, 49, 35–42. [Google Scholar] [CrossRef]
- Lee, H.-R.; Lee, S.-C.; Lee, D.H.; Jung, M.; Kwon, J.-H.; Huh, M.-J.; Kim, D.S.; Lee, J.-E.; Park, I.-K. Identification of aggregation-sex pheromone of the Korean Monochamus alternatus (Coleoptera: Cerambycidae) population, the main vector of pine wood nematode. J. Econ. Entomol. 2018, 111, 1768–1774. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Jung, Y.H.; Lee, S.-M. Diel Rhythmicity of Field Responses to Synthetic Pheromone Lures in the Pine Sawyer Monochamus saltuarius. Insects 2021, 12, 441. https://doi.org/10.3390/insects12050441
Kim J, Jung YH, Lee S-M. Diel Rhythmicity of Field Responses to Synthetic Pheromone Lures in the Pine Sawyer Monochamus saltuarius. Insects. 2021; 12(5):441. https://doi.org/10.3390/insects12050441
Chicago/Turabian StyleKim, Junheon, Young Hak Jung, and Sang-Myeong Lee. 2021. "Diel Rhythmicity of Field Responses to Synthetic Pheromone Lures in the Pine Sawyer Monochamus saltuarius" Insects 12, no. 5: 441. https://doi.org/10.3390/insects12050441
APA StyleKim, J., Jung, Y. H., & Lee, S. -M. (2021). Diel Rhythmicity of Field Responses to Synthetic Pheromone Lures in the Pine Sawyer Monochamus saltuarius. Insects, 12(5), 441. https://doi.org/10.3390/insects12050441