
A Review of Effects of Environment on Brain Size in Insects


Abstract
:Simple Summary
Abstract
1. Introduction
2. Brain Size
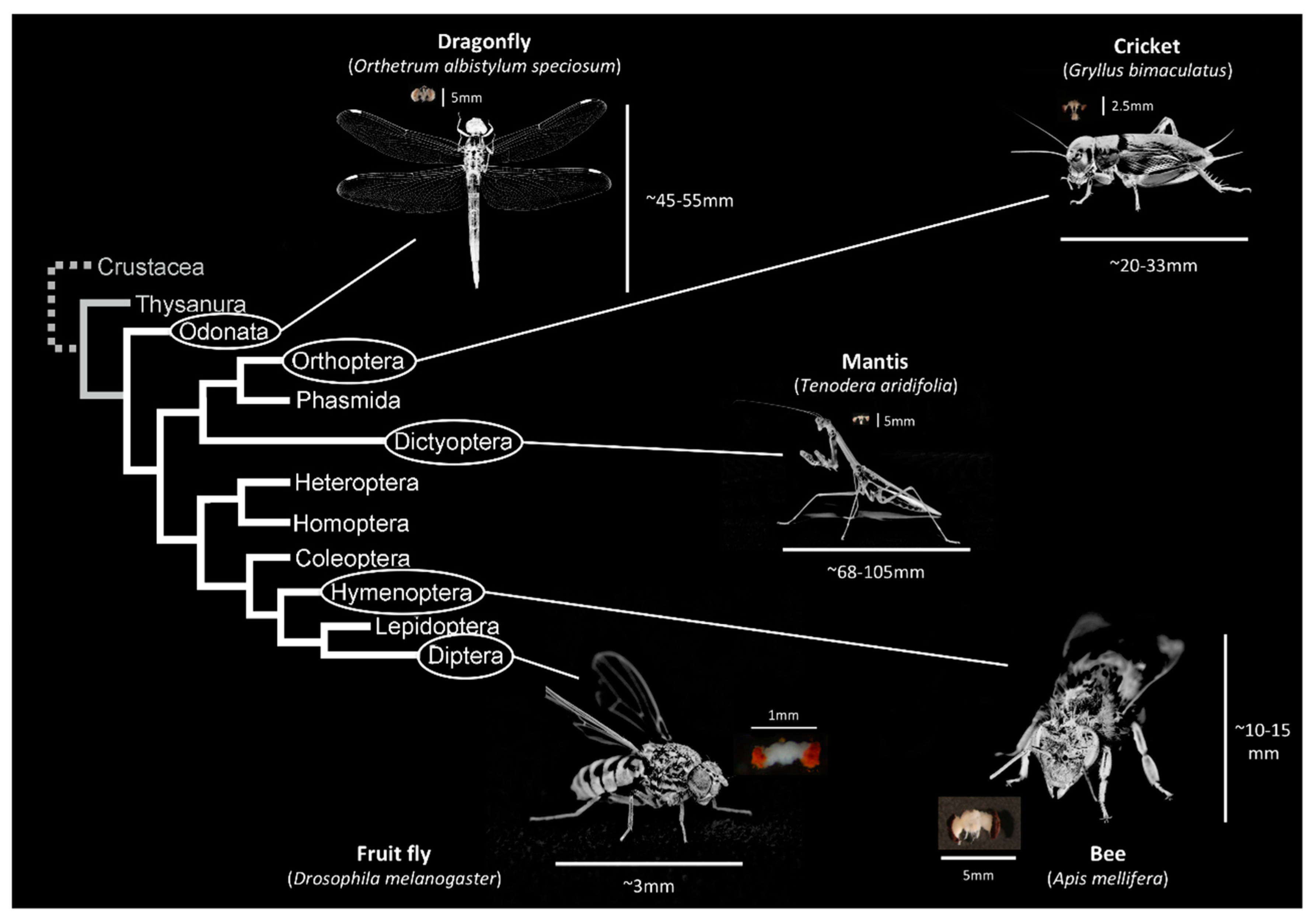
3. Haller’s Rule, Brain Size in Insects
4. Effect of Environment on Brain Size in Vertebrates
5. Effect of Environment on Brain Size in Insects
6. Balance Between Vision and Olfaction
7. Effect of Environment on Brain Size through This Vision and Olfaction Trade-Off
8. Vision and Olfaction Trade-Off A Universal Rule
9. Conclusions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Herculano-Houzel, S.; Messeder, D.J.; Fonseca-Azevedo, K.; Pantoja, N.A. When larger brains do not have more neurons: Increased numbers of cells are compensated by decreased average cell size across mouse individuals. Front. Neuroanat. 2015, 9, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerison, H.J. Brain to body ratios and the evolution of intelligence. Science 1955, 121, 447–449. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, E. Relative brain size and metabolism in mammals on JSTOR. Science 1983, 220, 1302–1304. [Google Scholar] [CrossRef]
- Armstrong, E. A look at relative brain size in mammals. Neurosci. Lett. 1982, 34, 101–104. [Google Scholar] [CrossRef]
- Burger, J.R.; George, M.A., Jr.; Leadbetter, C.; Shaikh, F. The allometry of brain size in mammals. J. Mammal. 2019, 100, 276–283. [Google Scholar] [CrossRef]
- Shultz, S.; Dunbar, R. Encephalization is not a universal macroevolutionary phenomenon in mammals but is associated with sociality. Proc. Natl. Acad. Sci. USA 2010, 107, 21582–21586. [Google Scholar] [CrossRef] [Green Version]
- Dunbar, R.I.M.; Shultz, S. Evolution in the social brain. Science 2007, 317, 1344–1347. [Google Scholar] [CrossRef] [Green Version]
- Dunbar, R.I.M. Neocortex size as a constraint on group-size in primates. J. Hum. Evol. 1992, 22, 469–493. [Google Scholar] [CrossRef]
- Barton, R.A. Neocortex and behavioural ecology in primates. Proc. R. Soc. B Biol. Sci. 1996, 263, 173–177. [Google Scholar]
- Dunbar, R.I.M. The social brain hypothesis. Evol. Anthropol. Issues News Rev. 1998, 6, 178–190. [Google Scholar] [CrossRef]
- Lihoreau, M.; Latty, T.; Chittka, L. An exploration of the social brain hypothesis in insects. Front. Physiol. 2012, 3, 442. [Google Scholar] [CrossRef] [Green Version]
- DeCasien, A.R.; Williams, S.A.; Higham, J.P. Primate brain size is predicted by diet but not sociality. Nat. Ecol. Evol. 2017, 1, 1–7. [Google Scholar] [CrossRef]
- González-Forero, M.; Gardner, A. Inference of ecological and social drivers of human brain-size evolution. Nature 2018, 557, 554–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farris, S.M.; Roberts, N.S. Coevolution of generalist feeding ecologies and gyrencephalic mushroom bodies in insects. Proc. Natl. Acad. Sci. USA 2005, 102, 17394–17399. [Google Scholar] [CrossRef] [Green Version]
- Sol, D.; Bacher, S.; Reader, S.M.; Lefebvre, L. Brain size predicts the success of mammal species introduced into novel environments. Am. Nat. 2008, 172, S63–S71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isler, K.; van Schaik, C. Costs of encephalization: The energy trade-off hypothesis tested on birds. J. Hum. Evol. 2006, 51, 228–243. [Google Scholar] [CrossRef]
- Gonzalez-Lagos, C.; Sol, D.; Reader, S.M. Large-brained mammals live longer. J. Evol. Biol. 2010, 23, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Benson-Amram, S.; Dantzer, B.; Stricker, G.; Swanson, E.M.; Holekamp, K.E. Brain size predicts problem-solving ability in mammalian carnivores. Proc. Natl. Acad. Sci. USA 2016, 113, 2532–2537. [Google Scholar] [CrossRef] [Green Version]
- Healy, S.D.; Rowe, C. A critique of comparative studies of brain size. Proc. R. Soc. B Biol. Sci. 2007, 274, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Chittka, L.; Niven, J. Are bigger brains better? Curr. Biol. 2009, 19, R995–R1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healy, S.D.; Rowe, C. Costs and benefits of evolving a larger brain: Doubts over the evidence that large brains lead to better cognition. Anim. Behav. 2013, 86, e1–e3. [Google Scholar] [CrossRef]
- Font, E.; García-Roa, R.; Pincheira-Donoso, D.; Carazo, P. Rethinking the effects of body size on the study of brain size evolution. Brain. Behav. Evol. 2019, 93, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Rensch, B. Increase of learning capability with increase of brain-size. Am. Nat. 1956, 90, 81–95. [Google Scholar] [CrossRef]
- Eberhard, W.G.; Wcislo, W.T. Grade changes in brain-body allometry. morphological and behavioural correlates of brain size in miniature spiders, insects and other invertebrates. In Advances in Insect Physiology; Academic Press Inc.: Cambridge, MA, USA, 2011; Volume 40, pp. 155–214. [Google Scholar]
- Seid, M.A.; Castillo, A.; Wcislo, W.T. The allometry of brain miniaturization in ants. Brain. Behav. Evol. 2011, 77, 5–13. [Google Scholar] [CrossRef]
- Polilov, A.A.; Makarova, A.A. The scaling and allometry of organ size associated with miniaturization in insects: A case study for Coleoptera and Hymenoptera. Sci. Rep. 2017, 7, 43095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, S.; Bulova, S. Development and evolution of brain allometry in wasps (Vespidae): Size, ecology and sociality. Curr. Opin. Insect Sci. 2017, 22, 54–61. [Google Scholar] [CrossRef]
- Roth, G.; Dicke, U. Evolution of the brain and intelligence. Trends Cogn. Sci. 2005, 9, 250–257. [Google Scholar] [CrossRef]
- Hart, B.L.; Hart, L.A.; McCoy, M.; Sarath, C.R. Congnitive behavior in Asia elephant: Use and modification of branches for fly switching. Anim. Behav. 2001, 62, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T. Brain-to-body ratios and time of maturation of the mouse brain. Am. J. Physiol. 1963, 204, 343–346. [Google Scholar] [CrossRef]
- Renzi-Hammond, L.M.; Sallet, J.; Cozzi, B.; Buffenstein, R.; Orr, M.E.; Garbarino, V.R.; Salinas, A. Extended postnatal brain development in the longest-lived rodent: Prolonged maintenance of neotenous traits in the naked mole-rat Brain. Front. Neurosci. 2016, 10, 504. [Google Scholar]
- Jerison, H.J.; Barlow, H.B. Animal intelligence as encephalization. Philos. Trans. R. Soc. B. Biol. Sci. 1985, 308, 21–35. [Google Scholar]
- Dicke, U.; Roth, G. Neuronal factors determining high intelligence. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehner, R.; Fukushi, T.; Isler, K. On being small: Brain allometry in ants. Brain. Behav. Evol. 2007, 69, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riveros, A.J.; Gronenberg, W. Brain allometry and neural plasticity in the bumblebee bombus occidentalis. Brain. Behav. Evol. 2010, 75, 138–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groothuis, J.; Smid, H.M. Nasonia parasitic wasps escape from Haller’s rule by diphasic, partially isometric brain-body size scaling and selective neuropil adaptations. Brain. Behav. Evol. 2017, 90, 243–254. [Google Scholar] [CrossRef]
- Van Der Woude, E.; Smid, H.M.; Chittka, L.; Huigens, M.E. Breaking Haller’s rule: Brain-body size isometry in a minute parasitic wasp. Brain. Behav. Evol. 2013, 81, 86–92. [Google Scholar] [CrossRef]
- Dunbar, R.I.M.; Shultz, S.; Dunbar, R.I.M.; Shultz, S. Understanding primate brain evolution. Philos. Trans. R. Soc. B Biol. Sci. 2009, 362, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Gowlett, J.; Gamble, C.; Dunbar, R. Human evolution and the archaeology of the social brain. Curr. Anthropol. 2012, 53, 693–722. [Google Scholar] [CrossRef]
- Niven, J.E.; Laughlin, S.B. Energy limitation as a selective pressure on the evolution of sensory systems. J. Exp. Biol. 2008, 211, 1792–1804. [Google Scholar] [CrossRef] [Green Version]
- Weisbecker, V.; Blomberg, S.; Goldizen, A.W.; Brown, M.; Fisher, D. The evolution of relative brain size in marsupials is energetically constrained but not driven by behavioral complexity. Brain. Behav. Evol. 2015, 85, 125–135. [Google Scholar] [CrossRef]
- Moran, D.; Softley, R.; Warrant, E.J. The energetic cost of vision and the evolution of eyeless Mexican cavefish. Sci. Adv. 2015, 1, e1500363. [Google Scholar] [CrossRef] [Green Version]
- Aiello, L.C.; Wheeler, P. The expensive-tissue hypothesis: The brain and the digestive system in human and primate evolution. Curr. Anthropol. 1995, 36, 199–221. [Google Scholar] [CrossRef]
- Bulova, S.; Purce, K.; Khodak, P.; Sulger, E.; O’Donnell, S. Into the black and back: The ecology of brain investment in Neotropical army ants (formicidae: Dorylinae). Sci. Nat. 2016, 103, 31. [Google Scholar] [CrossRef]
- Barton, R.A.; Purvis, A.; Harvey, P.H. Evolutionary radiation of visual and olfactory systems in primates, bats and insectivores. Philos. Trans. R. Soc. B Biol. Sci. 1995, 348, 381–392. [Google Scholar]
- Garamszegi, L.Z.; Møller, A.P.; Erritzøe, J. Coevolving avian eye size and brain size in relation to prey capture and nocturnality. Proc. R. Soc. B Biol. Sci. 2002, 269, 961–967. [Google Scholar] [CrossRef]
- O’Carroll, D.C.; Warrant, E.J. Vision in dim light: Highlights and challenges. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 1–8. [Google Scholar] [CrossRef]
- Tierney, S.M.; Friedrich, M.; Humphreys, W.F.; Jones, T.M.; Warrant, E.J.; Wcislo, W.T. Consequences of evolutionary transitions in changing photic environments. Austral Entomol. 2017, 56, 23–46. [Google Scholar] [CrossRef]
- Catania, K.C.; Kaas, J.H. The unusual nose and brain of the star-nosed mole, a star in the brain. Bioscience 1996, 46, 578–586. [Google Scholar] [CrossRef] [Green Version]
- Catania, K.C.; Remple, M.S. Somatosensory cortex dominated by the representation of teeth in the naked mole-rat brain. Proc. Natl. Acad. Sci. USA 2002, 99, 5692–5697. [Google Scholar] [CrossRef] [Green Version]
- Niven, J.E. Evolution: Convergent eye losses in fishy circumstances. Curr. Biol. 2008, 18, R27–R29. [Google Scholar] [CrossRef] [Green Version]
- Soares, D.; Niemiller, M.L. Sensory adaptations of fishes to subterranean environments. Bioscience 2013, 63, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Soares, D.; Yamamoto, Y.; Strickler, A.G.; Jeffery, W.R. The lens has a specific influence on optic nerve and tectum development in the blind cavefish Astyanax. Dev. Neurosci. 2004, 26, 308–317. [Google Scholar] [CrossRef]
- Meng, F.; Zhao, Y.; Titus, T.; Zhang, C.; Postlethwait, J.H. Brain of the blind: Transcriptomics of the golden-line cavefish brain. Curr. Zool. 2018, 64, 765–773. [Google Scholar] [CrossRef] [Green Version]
- Chow, K.L.; Riesen, A.H.; Newell, F.W. Degeneration of retinal ganglion cells in infant chimpanzees reared in darkness. J. Comp. Neurol. 1957, 107, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Giffin, F.; Mitchell, D.E. The rate of recovery of vision after early monocular deprivation in kittens. J. Physiol. 1978, 274, 511–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, S.; Berry, M. The effects of dark rearing on the development of the visual cortex of the rat. J. Comp. Neurol. 1978, 180, 277–300. [Google Scholar] [CrossRef]
- Borges, S.; Berry, M. Preferential orientation of stelliate cell dendrites in the visual cortex of the dark-reared rat. Brain Res. 1976, 112, 141–147. [Google Scholar] [CrossRef]
- Gabbott, P.L.A.; Stewart, M.G. Experimental brain research quantitative morphological effects of dark-rearing and light exposure on the synaptic connectivity of layer 4 in the rat visual cortex (area 17). Exp. Brain Res. 1987, 68, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Wiesel, T.N.; Hubel, D.H. Single-cell responses in striate cortex of kittens deprived of vision in one eye. J. Neurophysiol. 1963, 26, 1003–1017. [Google Scholar] [CrossRef]
- Wiesel, T.N.; Hubel, D.H. Effects of visual deprivation on morphology and physiology of cells in the cat’s lateral geniculate body. J. Neurophysiol. 1963, 26, 978–993. [Google Scholar] [CrossRef] [PubMed]
- Cragg, B.G. The effects of vision and dark-rearing on the size and density of synapses in the lateral geniculate nucleus measured by electron microscopy. Brain Res. 1969, 13, 53–67. [Google Scholar] [CrossRef]
- Jameie, S.B.E.D.; Abdolrahmani, M.; Nobakht, M. Effects of total light deprivation on dorsal lateral geniculate nucleus of male neonate rats. Oman Med. J. 2010, 25, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Henderson, N.D. Brain weight increases resulting from environmental enrichment: A directional dominance in mice. Science 1970, 169, 776–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, J.G.; Saravanan, A.; Helen Rodd, F. Rearing environment affects the brain size of guppies: Lab-reared guppies have smaller brains than wild-caught guppies. Ethology 2008, 115, 122–133. [Google Scholar] [CrossRef]
- Diamond, M.C.; Law, F.; Rhodes, H.; Lindner, B.; Rosenzweig, M.R.; Krech, D.; Bennett, E.L. Increases in cortical depth and glia numbers in rats subjected to enriched environment. J. Comp. Neurol. 1966, 128, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.R.; Bennett, E.L. Effects of differential environments on brain weights and enzyme activities in gerbils, rats, and mice. Dev. Psychobiol. 1969, 2, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Sol, D.; Duncan, R.P.; Blackburn, T.M.; Cassey, P.; Lefebvre, L. Big brains, enhanced cognition, and response of birds to novel environments. Proc. Natl. Acad. Sci. USA 2005, 102, 5460–5465. [Google Scholar] [CrossRef] [Green Version]
- Kotrschal, A.; Deacon, A.E.; Magurran, A.E.; Kolm, N. Predation pressure shapes brain anatomy in the wild. Evol. Ecol. 2017, 31, 619–633. [Google Scholar] [CrossRef] [Green Version]
- Gonda, A.; Trokovic, N.; Herczeg, G.; Laurila, A.; Merilä, J. Predation- and competition-mediated brain plasticity in Rana temporaria tadpoles. J. Evol. Biol. 2010, 23, 2300–2308. [Google Scholar] [CrossRef]
- Beston, S.M.; Broyles, W.; Walsh, M.R. Increased juvenile predation is not associated with evolved differences in adult brain size in Trinidadian killifish (Rivulus hartii). Ecol. Evol. 2017, 7, 884–894. [Google Scholar] [CrossRef]
- Samuk, K.; Xue, J.; Rennision, D.J. Exposure to predators does not lead to the evolution of larger brains in experimental populations of threespine stickleback. Evolution 2018, 72, 916–929. [Google Scholar] [CrossRef]
- Kotrschal, A.; Sundström, L.F.; Brelin, D.; Devlin, R.H.; Kolm, N. Inside the heads of David and Goliath: Environmental effects on brain morphology among wild and growth-enhanced coho salmon Oncorhynchus kisutch. J. Fish Biol. 2012, 81, 987–1002. [Google Scholar] [CrossRef] [PubMed]
- Kotrschal, A.; Rogell, B.; Maklakov, A.A.; Kolm, N. Sex-specific plasticity in brain morphology depends on social environment of the guppy, Poecilia reticulata. Behav. Ecol. Sociobiol. 2012, 66, 1485–1492. [Google Scholar] [CrossRef]
- Gonda, A.; Herczeg, G.; Merilä, J. Evolutionary ecology of intraspecific brain size variation: A review. Ecol. Evol. 2013, 3, 2751–2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, S.; Buechel, S.D.; Boussard, A.; Kotrschal, A.; Kolm, N. Plastic changes in brain morphology in relation to learning and environmental enrichment in the guppy (Poecilia reticulata). J. Exp. Biol. 2019, 222, jeb200402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebb, D.O. The Organization of Behavior: A Neuropsychological Theory; Wiley: New York, NY, USA, 1949. [Google Scholar]
- Rosenzweig, M.R.; Bennett, E.L. Psychobiology of Plasticity: Effects of Training and Experience on Brain and Behavior. Behav. Brain Res. 1996, 78, 57–65. [Google Scholar] [CrossRef]
- Technau, G.M. Fiber number in the mushroom bodies of adult Drosophila melanogaster depends on age, sex and experience. J. Neurogenet. 1984, 21, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Heisenberg, M.; Heusipp, M.; Wanke, C. Structural plasticity in the Drosophila brain. J. Neurosci. 1995, 15, 1951–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Amei, A.; de Belle, J.S.; Roberts, S.P. Environmental effects on Drosophila brain development and learning. J. Exp. Biol. 2018, 221, jeb.169375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ott, S.R.; Rogers, S.M. Gregarious desert locusts have substantially larger brains with altered proportions compared with the solitarious phase. Proc. R. Soc. B Biol. Sci. 2010, 277, 3087–3096. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Green, D.S.; Roberts, S.P.; de Belle, J.S. Thermal disruption of mushroom body development and odor learning in Drosophila. PLoS ONE 2007, 2, e1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Withers, G.S.; Fahrbach, S.E.; Robinson, G.E. Selective neuroanatomical plasticity and division of labour in the honeybee. Nature 1993, 364, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Durst, C.; Eichmüller, S.; Menzel, R. Development and experience lead to increased volume of subcompartments of the honey bee mushroom body. Behav. Neural Biol. 1994, 62, 259–263. [Google Scholar] [CrossRef]
- Julian, G.E.; Gronenberg, W. Reduction of brain volume correlates with behavioral changes in queen ants. Brain. Behav. Evol. 2002, 60, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Barth, M.; Hirsch, H.V.B.; Meinertzhagen, I.A.; Heisenberg, M. Experience-dependent developmental plasticity in the optic lobe of Drosophila melanogaster. J. Neurosci. 1997, 17, 1493–1504. [Google Scholar] [CrossRef] [Green Version]
- Barth, M.; Heisenberg, M. Vision affects mushroom bodies and central complex in Drosophila melanogaster. Learn. Mem. 1997, 4, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Stieb, S.M.; Muenz, T.S.; Wehner, R.; Rössler, W. Visual experience and age affect synaptic organization in the mushroom bodies of the desert ant Cataglyphis fortis. Dev. Neurobiol. 2010, 70, 408–423. [Google Scholar] [CrossRef]
- Jones, B.M.; Leonard, A.S.; Papaj, D.R.; Gronenberg, W. Plasticity of the worker bumble bee brain in relation to age and rearing environment. Brain. Behav. Evol. 2013, 82, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Özer, I.; Carle, T. Back to the light, coevolution between vision and olfaction in the “Dark-flies” (Drosophila melanogaster). PLoS ONE 2020, 15, e0228939. [Google Scholar] [CrossRef]
- Scotto-Lomassese, S.; Strambi, C.; Strambi, A.; Charpin, P.; Augier, R.; Aouane, A.; Cayre, M. Influence of environmental stimulation on neurogenesis in the adult insect brain. J. Neurobiol. 2000, 45, 162–171. [Google Scholar] [CrossRef]
- Scotto-Lomassese, S.; Strambi, C.; Aouane, A.; Strambi, A.; Cayre, M. Sensory inputs stimulate progenitor cell proliferation in an adult insect brain. Curr. Biol. 2002, 12, 1001–1005. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, S.H.; Ott, S.R. Brain composition in Godyris zavaleta, a diurnal butterfly, reflects an increased reliance on olfactory information. J. Comp. Neurol. 2015, 523, 869–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheehan, Z.B.V.; Kamhi, J.F.; Seid, M.A.; Narendra, A. Differential investment in brain regions for a diurnal and nocturnal lifestyle in Australian Myrmecia ants. J. Comp. Neurol. 2019, 527, 1261–1277. [Google Scholar] [CrossRef] [PubMed]
- Stöckl, A.L.; Heinze, S.; Charalabidis, A.; Jundi, B.; Warrant, E.J.; Kelber, A. Differential investment in visual and olfactory brain areas reflects behavioural choices in hawk moths. Sci. Rep. 2016, 6, 26041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stieb, S.M.; Kelber, C.; Wehner, R.; Rossler, W. Antennal-lobe organization in desert ants of the genus Cataglyphis. Brain Behav. Evol. 2011, 77, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Arganda, S.; Hoadley, A.P.; Razdan, E.S.; Muratore, I.B.; Traniello, J.F.A. The neuroplasticity of division of labor: Worker polymorphism, compound eye structure and brain organization in the leafcutter ant Atta cephalotes. J. Comp. Physiol. A 2020, 206, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Penick, C.A.; Ghaninia, M.; Haight, K.L.; Opachaloemphan, C.; Yan, H.; Reinberg, D.; Liebig, J. Reversible plasticity in brain size, behaviour and physiology characterizes caste transitions in a socially flexible ant (Harpegnathos saltator). Proc. R. Soc. B 2021, 288, 20210141. [Google Scholar] [CrossRef]
- Fahrbach, S.E.; Moore, D.; Capaldi, E.A.; Farris, S.M.; Robinson, G.E. Experience-expectant plasticity in the mushroom bodies of the honeybee. Learn. Mem. 1998, 5, 115–123. [Google Scholar]
- Hirsch, H.V.B.; Potter, D.; Zawierucha, D.; Choudhri, T.; Glasser, A.; Murphey, R.K.; Byers, D. Rearing in darkness changes visually guided choice behavior in Drosophila. Vis. Neurosci. 1990, 5, 281–289. [Google Scholar] [CrossRef]
- Fuse, N.; Kitamura, T.; Haramura, T.; Arikawa, K.; Imafuku, M. Evolution in the Dark-Adaptation of Drosophila in the Laboratory; Springer: New York, NY, USA, 2014; ISBN 978-4-431-54146-2. [Google Scholar]
- Guillet, A.; Stergiou, A.; Carle, T. Effect of light exposure upon food consumption and brain size in Dark-Flies (Drosophila melanogaster). Brain Behav. Evol. 2019, 94, 18–26. [Google Scholar] [CrossRef]
- Barton, R.A. Visual specialization and brain evolution in primates. Proc. R. Soc. B Biol. Sci. 1998, 265, 1933–1937. [Google Scholar] [CrossRef]
- Keesey, I.W.; Grabe, V.; Gruber, L.; Koerte, S.; Obiero, G.F.; Bolton, G.; Khallaf, M.A.; Kunert, G.; Lavista-Llanos, S.; Valenzano, D.R.; et al. Inverse resource allocation between vision and olfaction across the genus Drosophila. Nat. Commun. 2019, 10, 1162. [Google Scholar] [CrossRef] [Green Version]
- Gronenberg, W.; Liebig, J. Smaller brains and optic lobes in reproductive workers of the ant Harpegnathos. Naturwissenschaften 1999, 86, 343–345. [Google Scholar] [CrossRef]
- Corral-López, A.; Garate-Olaizola, M.; Buechel, S.D.; Kolm, N.; Kotrschal, A. On the role of body size, brain size, and eye size in visual acuity. Behav. Ecol. Sociobiol. 2017, 71, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehner, R. Desert ant navigation: How miniature brains solve complex tasks. J. Comp. Physiol. A 2003, 189, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Muscedere, M.L.; Gronenberg, W.; Moreau, C.S.; Traniello, J.F.A. Investment in higher order central processing regions is not constrained by brain size in social insects. Proc. R. Soc. B Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [Green Version]
- Strausfeld, N.J.; Hansen, L.; Li, Y.; Gomez, R.S.; Ito, K. Evolution, discovery, and interpretations of arthropod mushroom bodies. Learn. Mem. 1998, 5, 11–37. [Google Scholar] [PubMed]
- Farris, S.M. Evolution of insect mushroom bodies: Old clues, new insights. Arthropod Struct. Dev. 2005, 34, 211–234. [Google Scholar] [CrossRef]
- Kunz, T.H.; Fenton, M.B. Bat Ecology; University of Chicago Press: Chicago, IL, USA, 2003; ISBN 9780226462073. [Google Scholar]
- Crish, S.D.; Rice, F.L.; Park, T.J.; Comer, C.M. Somatosensory organization and behavior in naked mole-rats I: Vibrissa-like body hairs comprise a sensory array that mediates orientation to tactile stimuli. Brain Behav. Evol. 2003, 62, 141–151. [Google Scholar] [CrossRef]
- Sanes, J.R.; Zipursky, S.L. Design principles of insect and vertebrate visual systems. Neuron 2010, 66, 15–36. [Google Scholar] [CrossRef] [Green Version]
- Hansson, B.S.; Stensmyr, M.C. Evolution of insect olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristan, W.B. Early evolution of neurons. Curr. Biol. 2016, 26, R949–R954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giurfa, M.; Menzel, R. Cognitive Architecture of a Mini-Brain. In Adaptivity and Learning; Springer: Berlin/Heidelberg, Germany, 2001; Volume 5, pp. 22–48. [Google Scholar]
- Menzel, R.; Giurfa, M. Cognition by a mini brain. Nature 1999, 400, 718–719. [Google Scholar] [CrossRef] [PubMed]
- Mizunami, M.; Yokohari, F.; Takahata, M. Further exploration into the adaptive design of the arthropod “microbrain”: I. Sensory and memory-processing systems. Zool. Sci. 2004, 21, 1141–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Type | Species | Environmental Changes | Effects | Refs |
|---|---|---|---|---|
| Developmental modifications | Fruit flies | Social isolation, deprivation antennal input | Reduction in the number of fibres at the mushroom bodies | [79] |
| Extreme larval crowding | Absolute volume of calyx, optic lobes, central brain and central complex increased | [80] | ||
| Heat stress | Absolute volume of mushroom bodies reduced | [81] | ||
| Extreme larval crowding | Absolute volumes of antennal lobes, optic lobes and central complex reduced | |||
| Desert locusts | Aggregation | Gregarious locusts have larger brains (larger midbrain) | [82] | |
| Fruit flies | Heat stress | Absolute volume of antennal lobes, calyx and pedunculus reduced | [83] | |
| Honeybees | Foraging experience | Bigger olfactory glomeruli in nurses than foragers | [84] | |
| Experience (foragers vs. nurses) | Absolute volume of mushroom bodies increased | [85] | ||
| Ants | Dark and excavation (reduction of behaviours) | Reduction of medulla and total brain | [86] | |
| Fruit flies | Rearing in darkness | Absolute volume of optic lobes reduced | [87] | |
| Rearing in darkness | Absolute volume of mushroom bodies and central complex reduced | [88] | ||
| Ants | Light exposure | Mushroom body calycal growth and reduction in microglumeruli numbers in the visual and olfactory input regions of the calyx | [89] | |
| Bumble bees | Presence of visual stimuli | Relative volume of antennal lobes and mushroom bodies reduced | [90] | |
| Fruit flies | Light enrichment | Whole brain volume increased. Absolute and relative volume of optic lobes increased. Absolute and relative volume of antennal lobes decreased | [91] | |
| Crickets | Complex environmental and congeneric stimulations | Increased number of newborn cells in their mushroom bodies | [92] | |
| Enriched sensory and social conditions | Enhanced neuroblast proliferation in the mushroom bodies | [93] | ||
| Evolutive adaptations | Butterflies, moth | Diurnal vs. nocturnal | Inversed investment in visual and olfactory systems: diurnal species invest more in vision whereas nocturnal species invest more in olfaction | [94] |
| Ants | [95] | |||
| Hawk moths | [96] | |||
| Ants | Desert vs. forest | Number of glomeruli in the antennal lobes reduced | [97] | |
| Light vs. dark | Larger eye structure and visual neuropils in workers performing tasks in light | [98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carle, T. A Review of Effects of Environment on Brain Size in Insects. Insects 2021, 12, 461. https://doi.org/10.3390/insects12050461
Carle T. A Review of Effects of Environment on Brain Size in Insects. Insects. 2021; 12(5):461. https://doi.org/10.3390/insects12050461
Chicago/Turabian StyleCarle, Thomas. 2021. "A Review of Effects of Environment on Brain Size in Insects" Insects 12, no. 5: 461. https://doi.org/10.3390/insects12050461
APA StyleCarle, T. (2021). A Review of Effects of Environment on Brain Size in Insects. Insects, 12(5), 461. https://doi.org/10.3390/insects12050461