
Ornithologists’ Help to Spiders: Factors Influencing Spiders Overwintering in Bird Nesting Boxes


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- (1)
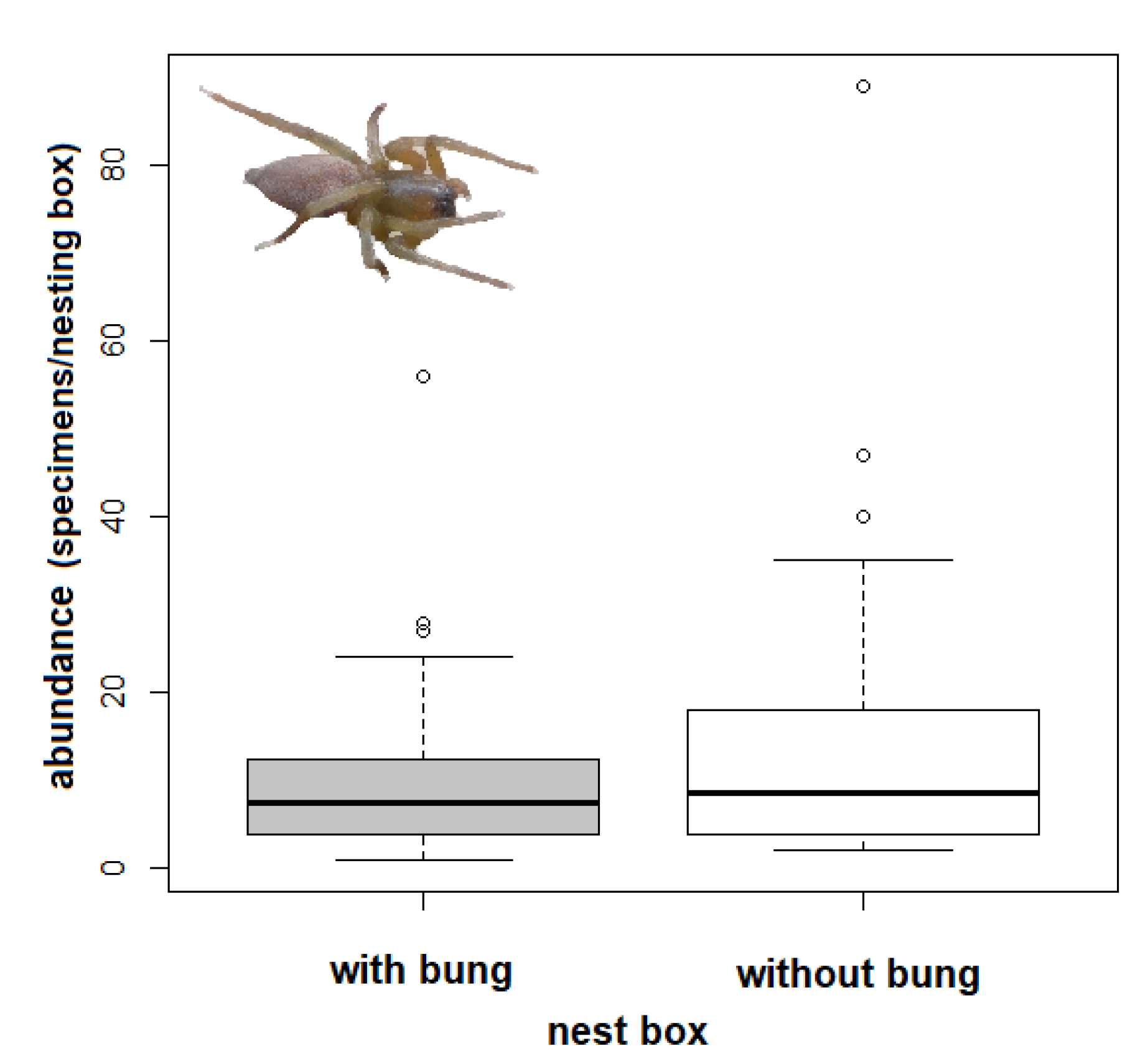
- The effect of tree species and bunging nesting boxes on spider abundance
- (2)
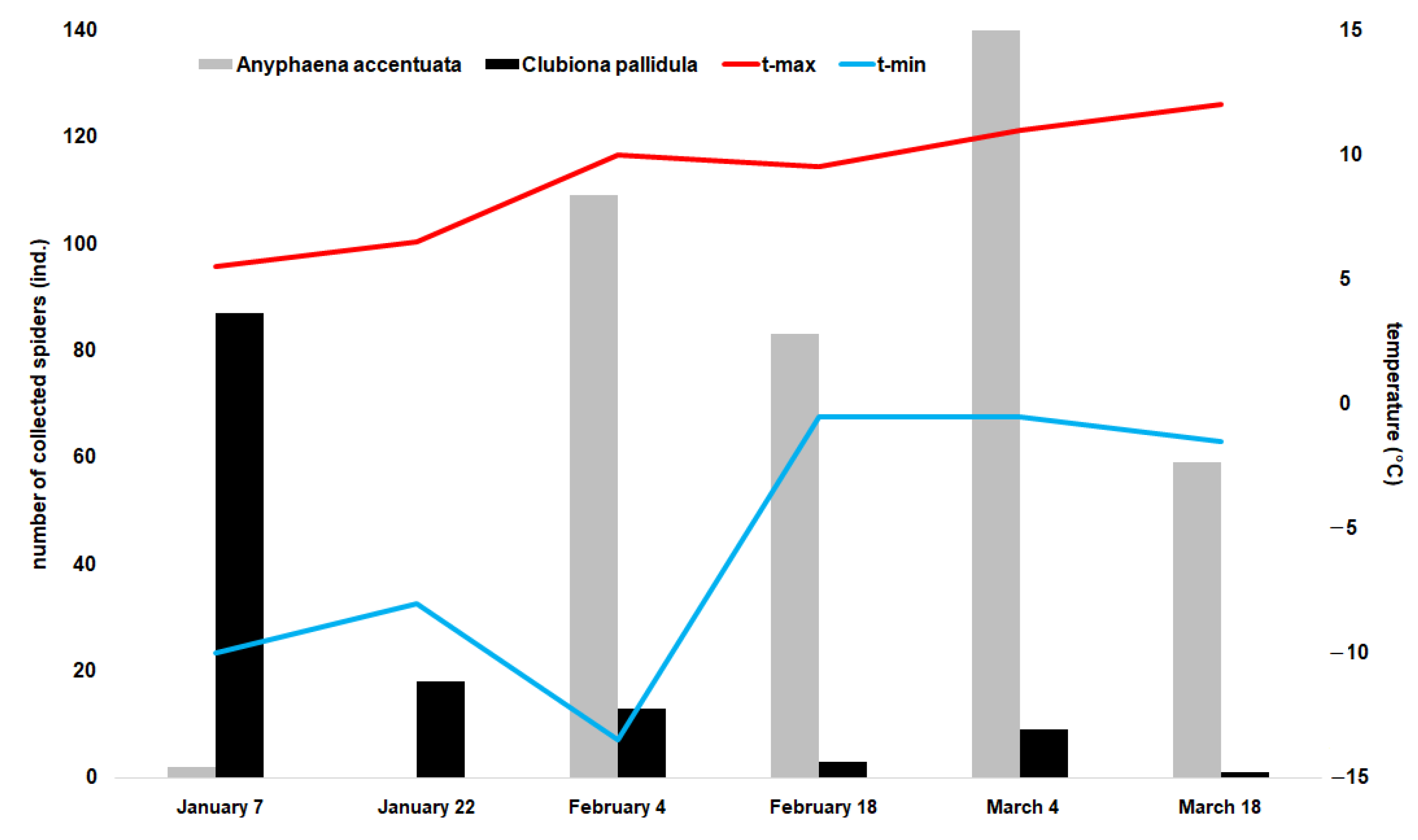
- The effect of temperature on nesting box resettlement by spiders
- (3)
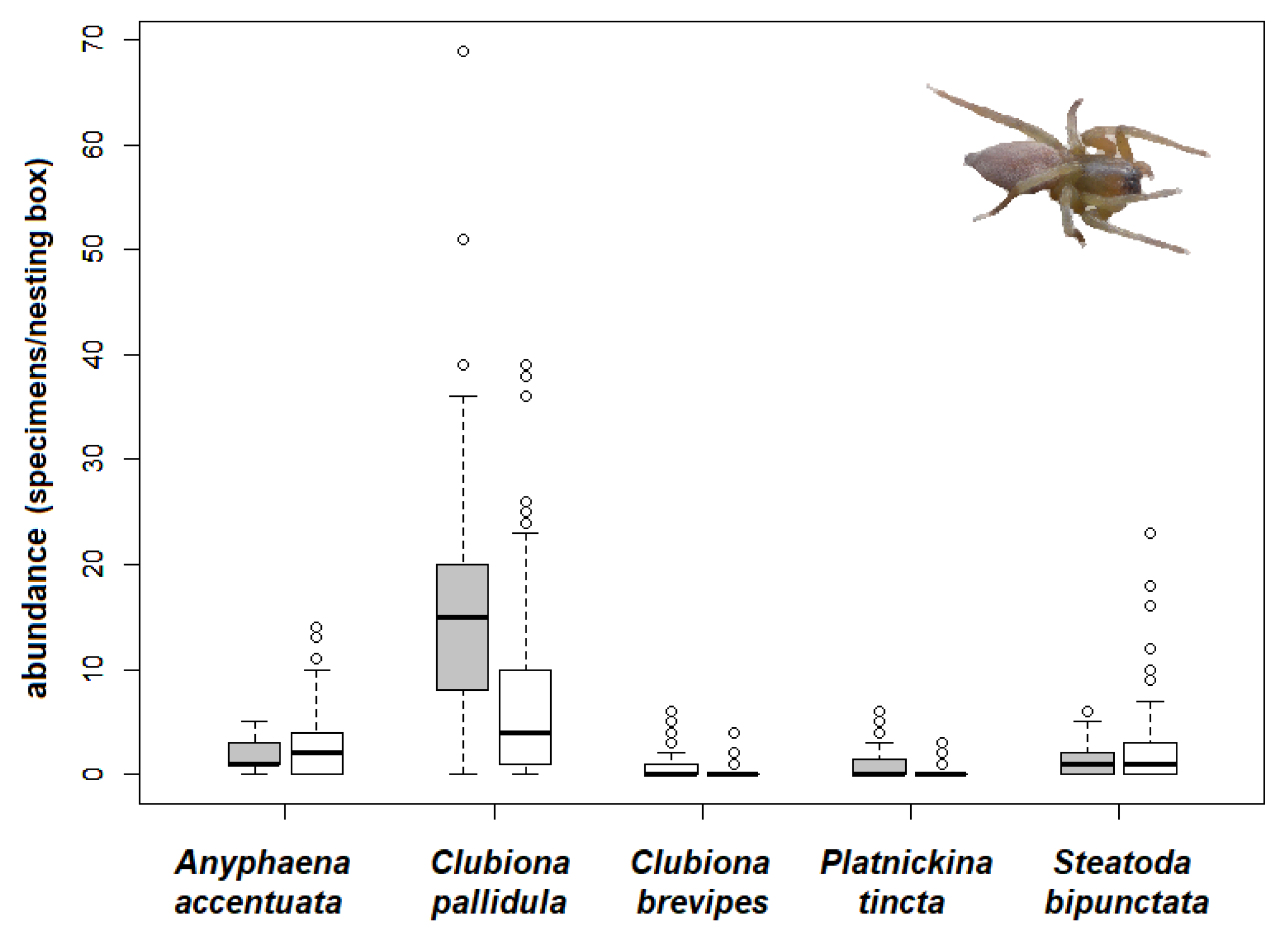
- The effect of the presence of nest material on spider abundance
3. Results
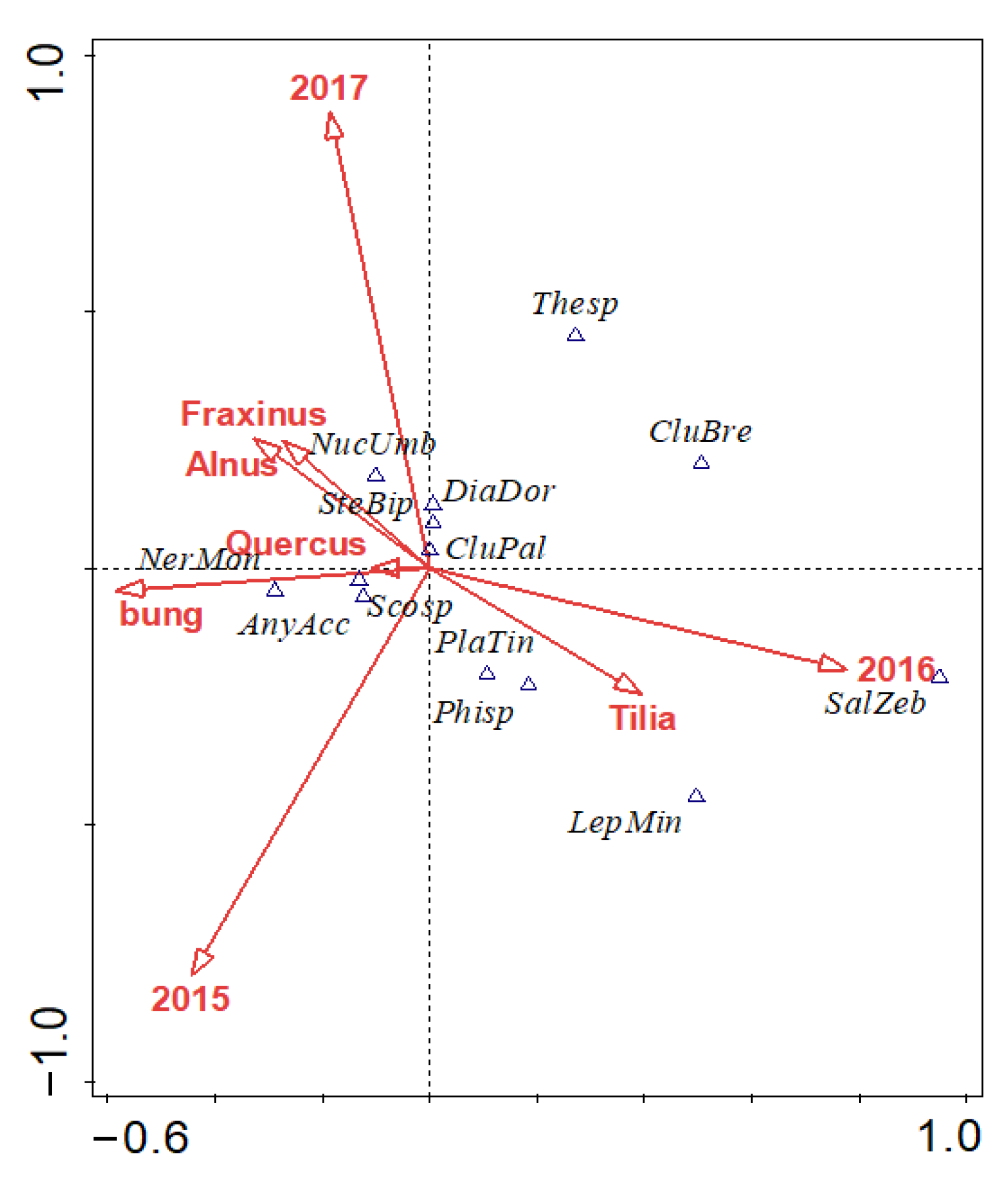
3.1. Assemblages of Spiders in Nesting Boxes
3.2. Effect of Tree Species and Bunging Nesting Boxes
3.3. Effect of Temperature on the Nesting Box Resettlement by Spiders
3.4. Effect of Presence of Nest Material on Spider Abundances
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Wunderlich, J. Mitteleuropäische Spinnen (Araneae) der Baumrinde. Z. Angew. Entomol. 1987, 94, 9–21. [Google Scholar] [CrossRef]
- Nikolai, V. The bark of trees: Thermal properties, microclimate and fauna. Oecologia 1986, 69, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Szinetár, C.; Horváth, R. A review of spiders on tree trunks in Europe (Araneae). Acta Zool. Bulg. 2005, 1, 221–257. [Google Scholar]
- Laška, V.; Kopecký, O.; Růžička, V.; Mikula, J.; Véle, A.; Šarapatka, B.; Tuf, I.H. Vertical distribution of spiders in soil. J. Arachnol. 2011, 39, 393–398. [Google Scholar] [CrossRef]
- Otto, S.; Floren, A. The spider fauna (Araneae) of tree canopies in the Bialowieza Forest. Fragm. Faunistica. 2007, 50, 57–70. [Google Scholar] [CrossRef]
- Machač, O.; Tuf, I.H. Spiders and harvestmen on tree trunks obtained by three sampling methods. Arachnol. Mitt. 2016, 51, 67–72. [Google Scholar] [CrossRef]
- Růžička, V.; Boháč, J.; Macek, J. Bezobratlí živočichové dutých stromů na Třeboňsku. Sbor. Jihočes. Muz. Čes. Budějovicích Přír. Vědy. 1991, 31, 33–46. [Google Scholar]
- Nitu, E.; Olenici, N.; Pop, I.; Nae, A.; Biris, I.A. Soil and saproxylic species (Coleoptera, Collembola, Araneae) in primeral forests from the northern part of South Eastern Carpathians. Ann. For. Res. 2009, 52, 27–54. [Google Scholar]
- Machač, O.; Christophoryová, J.; Krajčovičová, K.; Budka, J.; Schlaghamerský, J. Spiders and pseudoscorpions (Arachnida: Araneae, Pseudoscorpiones) in old oaks of Central European floodplains. Arachnol. Mitt. 2018, 56, 24–31. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, M.; Wichmann, C.; Schwager, M. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Siitonen, J.; Ranius, T. The importance of veteran trees for saproxylic insects. In Europe’s Changing Woods and Forests: From Wildwood to Cultural Landscape; Kirby, K., Watkins, C., Eds.; CAB International: Wallingford, UK, 2015; pp. 140–153. [Google Scholar]
- Mainwaring, M.C. The use of nestboxes by roosting birds during the non-breeding season: A review of the costs and benefits. Ardea 2011, 99, 167–176. [Google Scholar] [CrossRef]
- Robles, H.; Ciudad, C.; Matthysen, E. Tree-cavity occurrence, cavity occupation and reproductive performance of secondary cavity-nesting birds in oak forests: The role of traditional management practices. For. Ecol. Manag. 2011, 261, 1428–1435. [Google Scholar] [CrossRef]
- Nordberg, S. Biologisch-Ökologische Untersuchungen über die Vogelnidicolen. Acta Zool. Fenn. 1936, 21, 1–168. [Google Scholar]
- Svatoň, J. Pavúky (Araneida) v hniezdach belorítky obyčajnej (Delichon urbica) na západnom Slovensku. Zbor. Slov. Nár. Múz. Prír. Vedy. 1985, 31, 189–192. [Google Scholar]
- Miller, F.; Buchar, J. Spiders (Chelicerata: Araneae) collected in small mammal nests in the Western Carpathians. Acta Soc. Zool. Bohem. 1997, 61, 373–379. [Google Scholar]
- Krumpál, M.; Cyprich, D.; Fenďa, P.; Pinowski, J. Invertebrate fauna in nests of the house sparrow Paser domesticus and the tree sparrow Paser montanus in central Poland. Int. Stud. Sparrows 2001, 27–28, 35–58. [Google Scholar]
- Henenberg, J.; Řezáč, M.; Nováková, M. Spider assemblages in bird burrows. Biologia 2016, 73, 267–272. [Google Scholar] [CrossRef]
- Černecká, Ľ.; Michalko, R.; Krištín, A. Abiotic factors and biotic interactions jointly drive spider assemblages in nest-boxes in mixed forests. J. Arachnol. 2017, 45, 213–222. [Google Scholar] [CrossRef]
- Pekár, S. Some observations on overwintering of spiders (Araneae) in two contrasting orchards in the Czech Republic. Agric. Ecosyst. Environ. 1999, 73, 205–210. [Google Scholar] [CrossRef]
- Horváth, R.; Szinetár, C. 2002: Ecofaunistical study of bark-dwelling spiders (Araneae) on black pine (Pinus nigra) in urban and forest habitats. Acta Biol. Debrecina 2002, 24, 87–101. [Google Scholar]
- Horváth, R.; Lengyel, S.; Szinetár, C.; Jakab, S. The effects of prey availability on spider assemblages on European black pine (Pinus nigra) bark: Spatial patterns and guild structure. Can. J. Zool. 2005, 83, 324–335. [Google Scholar] [CrossRef]
- Spitzer, L.; Konvička, O.; Tropek, R.; Roháčová, M.; Tuf, I.H.; Nedvěd, O. Společenstvo členovců (Arthropoda) zimujících na jedli bělokoré na Valašsku (okr. Vsetín, Česká republika). [Assemblage of overwintering arthropods on white fir (Abies alba) in the Moravian Wallachia region (West Carpathians, Czech Republic)]. Časopis Slez. Muz. Opava 2010, 59, 217–232. [Google Scholar]
- Schaefer, M. Winter ecology of spiders (Araneida). Z. Angew. Entomol. 1977, 83, 113–134. [Google Scholar] [CrossRef]
- Aitchison, C.W. Low temperature feeding by winter-active spiders. J. Arachnol. 1984, 12, 297–305. [Google Scholar]
- Koomen, P. Winter activity of Anyphaena accentuata (Walckenaer, 1802) (Araneae: Anyphaenidae). In Proceedings of the 17th European Colloquium of Arachnology Edinburgh 1997; Selden, P., Ed.; British Arachnological Society: Edinburgh, UK, 1998; pp. 20–23. [Google Scholar]
- Krištofík, J.; Mašán, P.; Šustek, Z. Arthropods (Pseudoscorpionidea, Acari, Coleoptera, Siphonaptera) in the nests of the bearded tit (Panurus biarmicus). Biologia 2007, 62, 749–755. [Google Scholar] [CrossRef]
- Larrieu, L.; Cabanettes, A. Species, live status, and diameter are important tree features for diversity and abundance of tree microhabitats in subnatural montane beech-fir forests. Can. J. For. Res. 2012, 42, 1433–1445. [Google Scholar] [CrossRef]
- Bradshaw, R.H.W. Past anthropogenic influence on European forests and some possible genetic consequences. For. Ecol. Manag. 2004, 197, 203–212. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Twedt, D.J.; Henne-Kerr, J.L. Artificial cavities enhance breeding bird densities in managed cottonwood forests. Wildl. Soc. Bull. 2001, 29, 680–687. [Google Scholar]
- Mänd, R.; Tilgar, V.; Lõhmus, A.; Leivits, A. Providing nesting boxes for hole-nesting birds—Does habitat matter? Biodivers. Conserv. 2005, 14, 1823–1840. [Google Scholar] [CrossRef]
- Tyller, Z.; Paclík, M.; Remeš, V. Winter night inspections of nesting boxes affect their occupancy and reuse for roosting by cavity nesting birds. Acta Ornithol. 2012, 47, 79–85. [Google Scholar] [CrossRef]
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Kropf, C. Spiders of Europe. Version 01.2021. Available online: https://www.araneae.nmbe.ch (accessed on 10 January 2021).
- World Spider Catalog. World Spider Catalog. Version 21.5. Natural History Museum Bern. Available online: http://wsc.nmbe.ch (accessed on 10 January 2021).
- Cardoso, P.; Pekár, S.; Jocque, R.; Coddington, J. Global patterns of guild composition and functional diversity of spiders. PLoS ONE 2011, 6, e21710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination, Version 5.0; Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment For Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Řezáč, M.; Kůrka, A.; Růžička, V.; Heneberg, P. Red List of Czech spiders: 3rd edition, adjusted according to evidence-based national conservation priorities. Biologia 2015, 70, 645–666. [Google Scholar] [CrossRef]
- McComb, W.C.; Noble, E.R. Invertebrate Use of Natural Tree Cavities and Vertebrate Nesting boxes. Am. Midl. Nat. 1982, 107, 163–172. [Google Scholar] [CrossRef]
- Conner, R.N.; Saenz, D.; Rudolph, D.C. Fauna using nesting boxes in four timber types in eastern Texas. Bull. Tex. Ornithol. Soc. 1995, 28, 2–6. [Google Scholar]
- Gajdoš, P.; Krištofík, J.; Šustek, Z. Spiders (Araneae) in the birds’ nests in Slovakia. Biologia 1991, 48, 887–905. [Google Scholar]
- Kubcová, L.; Schlaghamerský, J. Zur Spinnenfauna der Stammregion stehenden Totholzes in südmährischen Auenwäldern. Arachnol. Mitt. 2002, 24, 35–61. [Google Scholar] [CrossRef]
- Machač, O. Pavouci a sekáči na kmenech stromů Hostýnsko-vsetínské hornatiny. (Spiders and harvestmen on tree trunks in Hostýnsko-vsetínská highlands). Acta Carp. Occ. 2014, 5, 64–67. [Google Scholar]
- Isaia, M.; Bona, F.; Badino, G. Comparison of polyethylene bubble wrap and corrugated cardboard traps for sampling tree inhabiting spiders. Environ. Entomol. 2006, 35, 1654–1660. [Google Scholar] [CrossRef]
- Pekár, S.; Michalko, R.; Loverre, P.; Líznarová, E.; Černecká, Ľ. Biological control in winter: Novel evidence for the importance of generalist predators. J. Appl. Ecol. 2015, 52, 270–279. [Google Scholar] [CrossRef]
- Krištín, A.; Mihál, I.; Urban, P. Roosting of the great tit, Parus major and the nuthatch, Sitta europaea in nesting boxes in an oak-hornbeam forest. Folia Zool. 2001, 50, 43–53. [Google Scholar]
- Korenko, S.; Kula, E.; Šimon, V.; Michalková, V.; Pekár, S. Are arboreal spiders associated to particular tree canopies? North-West J. Zool. 2011, 7, 261–269. [Google Scholar]
- Petráková, L.; Michalko, R.; Loverre, P.; Sentenská, L.; Korenko, S.; Pekár, S. Intraguild predation among spiders and their effect on the pear psylla during winter. Agric. Ecosyst. Environ. 2016, 233, 67–74. [Google Scholar] [CrossRef]
- Korenko, S.; Pekár, S. Is there intraguild predation between winter-active spiders (Araneae) on apple tree bark? Biol. Control. 2010, 54, 206–212. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Families | Ecological Niche | Hunter Guild | Hand Collected | Heat Extracted |
|---|---|---|---|---|
| Agelenidae | ||||
| Tegenaria silvestris L. Koch, 1872 | ground, trunk base | sheet web | 3 | 0 |
| Anyphaenidae | ||||
| Anyphaena accentuata (Walckenaer, 1802) | branches, bark | other hunters | 851 | 67 |
| Araneidae | ||||
| Mangora acalypha (Walckenaer, 1802) | herbs, branches | orb web | 1 | 0 |
| Nuctenea umbratica (Clerck, 1757) | bark | orb web | 14 | 0 |
| Zilla diodia (Walckenaer, 1802) | branches | orb web | 1 | 0 |
| Clubionidae | ||||
| Clubiona brevipes Blackwall, 1841 | branches | other hunters | 84 | 31 |
| Clubiona pallidula (Clerck, 1757) | bark | other hunters | 1635 | 574 |
| Gnaphosidae | ||||
| Micaria subopaca Westring, 1861 | bark | ground hunters | 4 | 2 |
| Scotophaeus sp. | hollows | ground hunters | 112 | 9 |
| Linyphiidae | ||||
| Lepthyphantes minutus (Blackwall, 1833) | bark | sheet web | 6 | 1 |
| Neriene montana (Clerck, 1757) | hollows, branches | sheet web | 28 | 2 |
| Philodromidae | ||||
| Philodromus dispar Walckenaer, 1826 | branches | other hunters | 1 | 0 |
| Philodromus sp. | branches | other hunters | 105 | 25 |
| Salticidae | ||||
| Pseudicius encarpatus (Walckenaer, 1802) | bark | other hunters | 2 | 0 |
| Salticus zebraneus (C. L. Koch, 1837) | bark | other hunters | 20 | 9 |
| Theridiidae | ||||
| Dipoena sp. | bark | space web | 2 | 0 |
| Platnickina tincta (Walckenaer, 1802) | branches | space web | 162 | 42 |
| Steatoda bipunctata (Linnaeus, 1758) | hollow | space web | 449 | 51 |
| Theridion sp. | bark, branches | space web | 14 | 3 |
| Thomisidae | ||||
| Diaea dorsata (Fabricius, 1777) | branches | ambush hunters | 12 | 3 |
| Tetragnathidae | ||||
| Tetragnatha sp. | branches | orb web | 5 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machač, O.; Tuf, I.H. Ornithologists’ Help to Spiders: Factors Influencing Spiders Overwintering in Bird Nesting Boxes. Insects 2021, 12, 465. https://doi.org/10.3390/insects12050465
Machač O, Tuf IH. Ornithologists’ Help to Spiders: Factors Influencing Spiders Overwintering in Bird Nesting Boxes. Insects. 2021; 12(5):465. https://doi.org/10.3390/insects12050465
Chicago/Turabian StyleMachač, Ondřej, and Ivan Hadrián Tuf. 2021. "Ornithologists’ Help to Spiders: Factors Influencing Spiders Overwintering in Bird Nesting Boxes" Insects 12, no. 5: 465. https://doi.org/10.3390/insects12050465
APA StyleMachač, O., & Tuf, I. H. (2021). Ornithologists’ Help to Spiders: Factors Influencing Spiders Overwintering in Bird Nesting Boxes. Insects, 12(5), 465. https://doi.org/10.3390/insects12050465