
Biological and Host Range Characteristics of Lysathia flavipes (Coleoptera: Chrysomelidae), a Candidate Biological Control Agent of Invasive Ludwigia spp. (Onagraceae) in the USA

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Origin and Rearing of Lysathia flavipes
2.2. Egg Development of Lysathia flavipes
2.3. Preimaginal Development of Lysathia flavipes
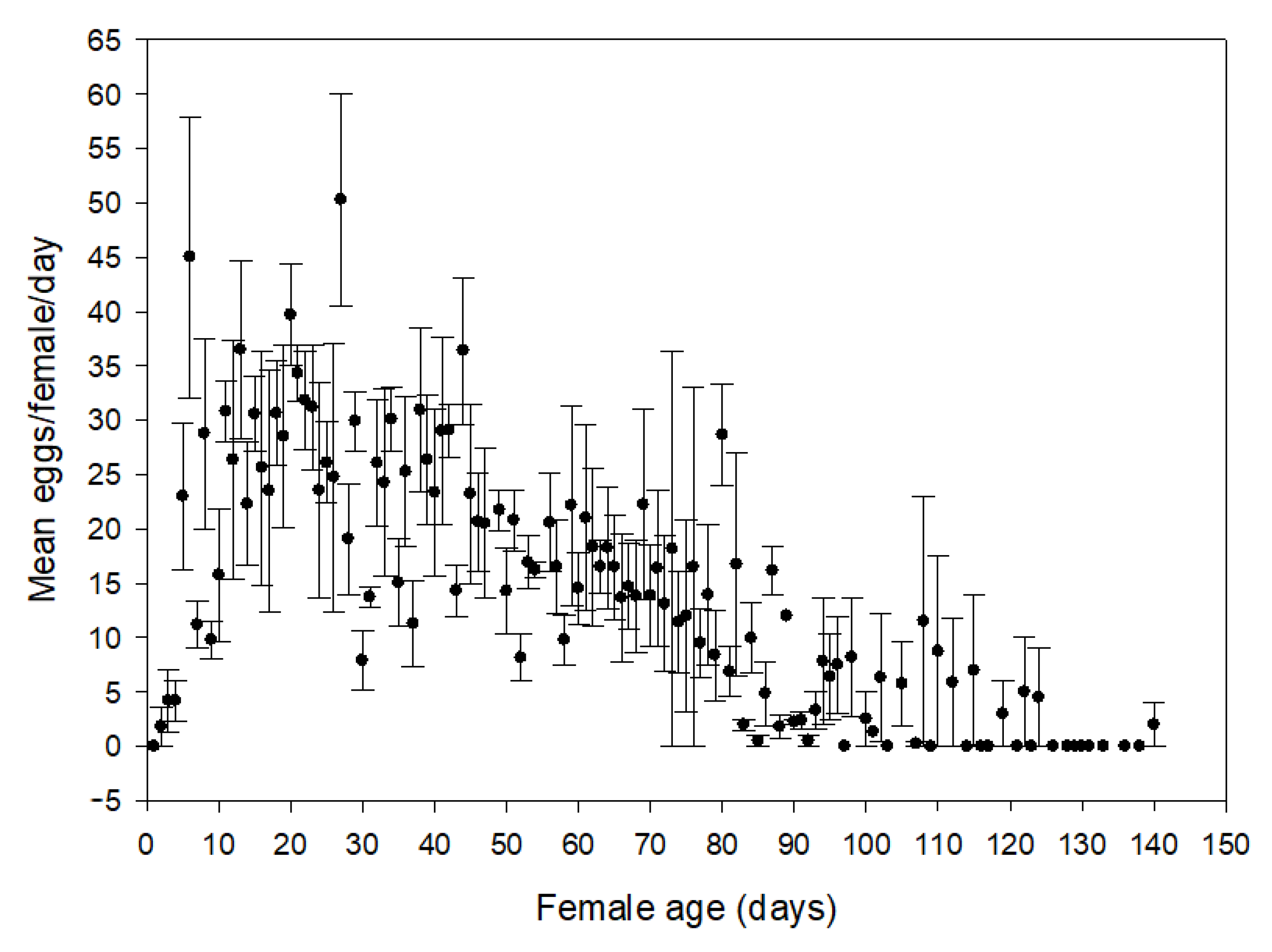
2.4. Fecundity of Lysathia flavipes
2.5. Host Range Experiments: Test Plants
2.6. No-Choice Development and Oviposition Tests
2.7. Multiple-Choice Oviposition Tests
2.8. Data Analyses
3. Results
3.1. Life History of Lysathia flavipes
3.2. No-Choice Host Range Tests
3.3. Multiple-Choice Host Range Tests
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wagner, W.L.; Hoch, P.C.; Raven, P.H. Revised classification of the Onagraceae. Syst. Bot. Monogr. 2007, 83, 1–240. [Google Scholar]
- Pesamosca, S.C.; Boldrini, I.I. Ludwigia litoranea (Onagraceae), a new species from coastal southern Brazil. Phytotaxa 2015, 230, 183–188. [Google Scholar] [CrossRef]
- Liu, S.; Hoch, P.; Diazgranados, M.; Raven, P.H.; Barber, J.C. Multi-locus phylogeny of Ludwigia (Onagraceae): Insights on infra-generic relationships and the current classification of the genus. TAXON 2017, 66, 1112–1127. [Google Scholar] [CrossRef]
- Hoch, P.C.; Wagner, W.L.; Raven, P.H. The correct name for a section of Ludwigia L. (Onagraceae). PhytoKeys 2015, 50, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Thouvenot, L.; Haury, J.; Thiébaut, G. A success story: Water primroses, aquatic plant pests. Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 790–803. [Google Scholar] [CrossRef]
- Kim, H.W.; Son, D.C.; Park, S.H.; Jang, C.; Sun, E.; Jo, H.; Yun, S.M.; Chang, K.S. Unrecorded alien plant in south Korea: Ludwigia peploides subsp. montevidensis (Spreng.) P.H. Raven. Korean J. Plant. Res. 2019, 32, 201–206. [Google Scholar] [CrossRef]
- Cronk, Q.C.B.; Fuller, J.L. Plant Invaders: The Threat to Natural Ecosystems; Earthscan Publications Ltd.: London, UK, 2001. [Google Scholar]
- Grewell, B.J.; Netherland, M.D.; Skaer Thomason, M.J.S. Establishing Research and Management Priorities for Invasive Water Primroses (Ludwigia spp.); ERDC/EL TR-162 Technical Report; U.S. Army Engineer Research and Development Center: Washington, DC, USA, 2016; p. 44. [Google Scholar]
- Grewell, B.J.; Skaer Thomason, M.J.; Futrell, C.J.; Iannucci, M.; Drenovsky, R.E. Trait responses of invasive aquatic macrophyte congeners: Colonizing diploid outperforms polyploid. AoB Plants 2016, 8, plw014. [Google Scholar] [CrossRef] [Green Version]
- Grewell, B.J.; Gillard, M.B.; Futrell, C.J.; Castillo, J.M. Seedling emergence from seed banks in Ludwigia hexapetala-invaded wetlands: Implications for restoration. Plants 2019, 8, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiers, I.; Crohain, N.; Josens, G.; Triest, L. Impact of three aquatic invasive species on native plants and macroinvertebrates in temperate ponds. Biol. Invasions 2011, 13, 2715–2726. [Google Scholar] [CrossRef]
- Khanna, S.; Santos, M.J.; Boyer, J.D.; Shapiro, K.D.; Bellvert, J.; Ustin, S.L. Water primrose invasion changes successional pathways in an estuarine ecosystem. Ecosphere 2018, 9, e02418. [Google Scholar] [CrossRef]
- Sarat, E.; Mazaubert, E.; Dutartre, A.; Poulet, N.; Soubeyran, Y. Water primrose (Ludwigia spp.). In Invasive Alien Species in Aquatic Environments. Practical Information and Management Insights; Sarat, E., Mazaubert, E., Dutartre, A., Poulet, N., Soubeyran, Y., Eds.; Knowledge for Action Series; ONEMA (French National Agency for Water and Aquatic Environments): Paris, France, 2015; Volume 2, pp. 54–73. [Google Scholar]
- Sarat, E.; Blottière, D.; Dutartre, A.; Poulet, N.; Soubeyran, Y. Water primrose (Ludwigia spp.). In Invasive Alien Species in Aquatic Environments. Practical Information and Management Insights (Cont.); Sarat, E., Blottière, D., Dutartre, A., Poulet, N., Soubeyran, Y., Eds.; Knowledge for Action Series; French Biodiversity Agency (AFB): Paris, France, 2018; Volume 3, pp. 65–88. [Google Scholar]
- Gillard, M.; Thiébaut, G.; Deleu, C.; Leroy, D. Present and future distribution of three aquatic plants taxa across the world: Decrease in native and increases in invasive ranges. Biol. Invasions 2017, 19, 2159–2170. [Google Scholar] [CrossRef]
- Gillard, M.; Grewell, B.J.; Futrell, C.J.; Deleu, C.; Thiébaut, G. Germination and seedling growth of water primroses: A cross experiment between two invaded ranges with contrasting climates. Front. Plant. Sci. 2017, 8, 1677. [Google Scholar] [CrossRef] [PubMed]
- van Driesche, R.G.; Carruthers, R.I.; Center, T.; Hoddle, M.S.; Hough-Goldstein, J.; Morin, L.; Smith, L.; Wagner, D.L.; Blossey, B.; Brancatini, V.; et al. Classical biological control for the protection of natural ecosystems. Biol. Control. 2010, 54, S2–S33. [Google Scholar] [CrossRef] [Green Version]
- Reddy, A.M.; Pratt, P.D.; Grewell, B.J.; Harms, N.E.; Cabrera Walsh, G.; Hérnandez, M.C.; Faltlhauser, A.; Cibils-Stewart, X. Biological control of invasive water primroses, Ludwigia spp., in the United States: A feasibility assessment. J. Aquat. Plant. Manag. 2021, in press. [Google Scholar]
- Cordo, H.A.; DeLoach, C.J. Notes on the weevils Tyloderma, Auletes, and other aquatic plants in the southern South America. Coleopt. Bull. 1982, 36, 291–297. [Google Scholar]
- Cordo, H.A.; DeLoach, C.J. The flea beetle, Lysathia flavipes, that attacks Ludwigia (water primrose) and Myriophyllum (Parrotfeather) in Argentina. Coleopt. Bull. 1982, 36, 299–302. [Google Scholar]
- Hernández, M.C.; Cabrera Walsh, G. Insect herbivores associated with Ludwigia species, Oligospermum section, in their Argentine distribution. J. Insect Sci. 2014, 14, ieu063. [Google Scholar] [CrossRef]
- Kelch, D. California Pest Rating for Ludwigia decurrens (Winged Water-Primrose) Myrtales; Onagraceae; Plant Health and Pest Prevention Services Division, California Department of Food and Agriculture: Sacramento, CA, USA, 2015. Available online: https://blogs.cdfa.ca.gov/Section3162/?p=1377 (accessed on 15 March 2021).
- McEvoy, P.B.; Cox, C.; Coombs, E. Successful biological control of ragwort, Senecio jacobaea, by introduced insects in Oregon. Ecol. Appl. 1991, 1, 430–442. [Google Scholar] [CrossRef]
- Buckingham, G.R. Biological Control of Alligatorweed, Alternanthera philoxeroides, the World’s First Aquatic Weed Success Story. Castanea 1996, 61, 232–243. [Google Scholar]
- Kirby, D.R.; Carlson, R.B.; Krabbenhoft, K.D.; Mundal, D.; Kirby, M.M. Biological control of leafy spurge with introduced flea beetles (Aphthona spp.). J. Range Manag. 2000, 53, 305–308. [Google Scholar] [CrossRef]
- Zimmermann, H.G.; Moran, V.C.; Hoffmann, J.H. Biological control in the management of invasive alien plants in South Africa, and the role of the Working for Water programme. S. Afr. J. Sci. 2004, 100, 34–40. [Google Scholar]
- Habeck, D.H.; Wilkerson, R. The life cycle of Lysathia ludoviciana (Fall)(Coleoptera: Chrysomelidae) on parrotfeather, Myriophyllum aquaticum (Velloso) Verde. Coleopt. Bull. 1980, 34, 167–170. [Google Scholar]
- Vogt, G.B.; Cordo, H.A. Recent South American field studies of prospective biocontrol agents of weeds. In Proceedings of the Research Planning Conference on the Aquatic Plant Control Program, Charleston, SC, USA, 22–24 October 1975; Final Report. U.S. Army Engineer Waterways Experiment Station: Vicksburg, MS, USA, 1976. Paper A-76-1. pp. 36–55. [Google Scholar]
- Montero Bulacio, N.; Oviedo Bustos, R.; Montero, G. Insectos herbívoros asociados a Oenothera indecora Cambess (Onagraceae) en la region Pampeana Norte. Agromensajes 2013, 35, 1–5. [Google Scholar]
- Cilliers, C.J. Biological control of parrot’s feather, Myriophyllum aquaticum (Vell.) Verdc. (Haloragaceae), in South Africa. In Biological Control of Weeds in South Africa (1990–1998); African Entomology Memoir; Olckers, T., Hill, M.P., Eds.; Entomological Society of Southern Africa: Hatfield, South Africa, 1999; Volume 1, pp. 113–118. [Google Scholar]


{kind=link}
| Test-Plant | Larval Survivorship (Prop.) 4 | Larval Development (Days) | Number of Eggs Per Female | Egg Hatching Rate Per Female |
|---|---|---|---|---|
| Ludwigiahexapetala1 | 0.63 ± 0.07 (15) abc | 20.8 ± 0.9 (14) a | 154.1 ± 22.2 (12) b | 0.62 ± 0.08 (9) a |
| L. peploides subsp. peploides 1 | 0.85 ± 0.06 (5) ab | 18.7 ± 1.2 (5) a | 178.8 ± 5.8 (4) ab | 0.65 ± 0.17 (4) a |
| L. peploides subsp. montevidensis 1 | 0.75 ± 0.08 (5) abc | 17.5 ± 0.6 (5) a | 109.8 ± 37.2 (5) bc | 0.57 ± 0.16 (5) a |
| L. decurrens2 | 0 (5) | |||
| L. polycarpa3 | 0.95 ± 0.05 (5) a | 20.9 ± 0.9 (5) a | 325.6 ± 44.4 (5) a | 0.59 ± 0.06 (5) a |
| L. repens3 | 0.85 ± 0.06 (5) ab | 21.1 ± 0.3 (5) a | 25 ± 4.1 (5) c | 0.82 ± 0.04 (5) a |
| L. palustris3 | 0.45 ± 0.09 (5) bc | 21.6 ± 1.3 (5) a | 128.5 ± 22.5 (4) b | 0.68 ± 0.10 (5) a |
| Epilobium ciliatum subsp. ciliatum 3 | 0.85 ± 0.06 (5) ab | 19.2 ± 0.3 (5) a | 161.2 ± 27.5 (5) ab | 0.43 ± 0.11 (5) a |
| E. canum3 | 0 (5) | |||
| Clarkia amoena3 | 0.25 ± 0 (5) c | 20 ± 1.2 (5) a | ||
| Oenothera elata subsp. hookeri 3 | 0 (5) |
| Test-Plant | Number of Eggs Per Test Plant 4 | Range |
|---|---|---|
| Ludwigiahexapetala1 | 44.1 ± 8.8 (15) abcde | 0–96 |
| L. peploides subsp. peploides 1 | 72.4 ± 15.9 (5) abd | 31–125 |
| L. peploides subsp. montevidensis 1 | 6 ± 3.7 (5) ce | 0–15 |
| L. decurrens2 | 3.8 ± 3.1 (5) de | 0–16 |
| L. polycarpa3 | 28.4 ± 13 (5) abcde | 0–66 |
| L. repens3 | 58 ± 14.4 (5) abcde | 23–90 |
| L. palustris3 | 45.8 ± 18.3 (5) bcde | 7–106 |
| Epilobium ciliatum subsp. ciliatum 3 | 43 ± 17.0 (5) abcde | 20–110 |
| E. canum3 | 2.4 ± 2.4 (5) bcde | 0–12 |
| Clarkia amoena3 | 171.6 ± 53.3 (5) a | 96–381 |
| Oenothera elata subsp. hookeri 3 | 69.2 ± 15.0 (5) abc | 44–124 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddy, A.M.; Pratt, P.D.; Grewell, B.J.; Harms, N.E.; Cibils-Stewart, X.; Cabrera Walsh, G.; Faltlhauser, A. Biological and Host Range Characteristics of Lysathia flavipes (Coleoptera: Chrysomelidae), a Candidate Biological Control Agent of Invasive Ludwigia spp. (Onagraceae) in the USA. Insects 2021, 12, 471. https://doi.org/10.3390/insects12050471
Reddy AM, Pratt PD, Grewell BJ, Harms NE, Cibils-Stewart X, Cabrera Walsh G, Faltlhauser A. Biological and Host Range Characteristics of Lysathia flavipes (Coleoptera: Chrysomelidae), a Candidate Biological Control Agent of Invasive Ludwigia spp. (Onagraceae) in the USA. Insects. 2021; 12(5):471. https://doi.org/10.3390/insects12050471
Chicago/Turabian StyleReddy, Angelica M., Paul D. Pratt, Brenda J. Grewell, Nathan E. Harms, Ximena Cibils-Stewart, Guillermo Cabrera Walsh, and Ana Faltlhauser. 2021. "Biological and Host Range Characteristics of Lysathia flavipes (Coleoptera: Chrysomelidae), a Candidate Biological Control Agent of Invasive Ludwigia spp. (Onagraceae) in the USA" Insects 12, no. 5: 471. https://doi.org/10.3390/insects12050471
APA StyleReddy, A. M., Pratt, P. D., Grewell, B. J., Harms, N. E., Cibils-Stewart, X., Cabrera Walsh, G., & Faltlhauser, A. (2021). Biological and Host Range Characteristics of Lysathia flavipes (Coleoptera: Chrysomelidae), a Candidate Biological Control Agent of Invasive Ludwigia spp. (Onagraceae) in the USA. Insects, 12(5), 471. https://doi.org/10.3390/insects12050471