
Fat Body—Multifunctional Insect Tissue



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Morphology and Anatomy of the Fat Body
2.1. Trophocytes
2.2. Oenocytes
2.3. Mycetocytes
2.4. Chromatocytes
2.5. Urocytes
2.6. Microbiota of the Fat Body
3. Differentiation of the Fat Body
3.1. Histotypes and Regional and Structural Differences
3.2. Metamorphosis of the Fat Body
4. Role and Metabolism of Lipids
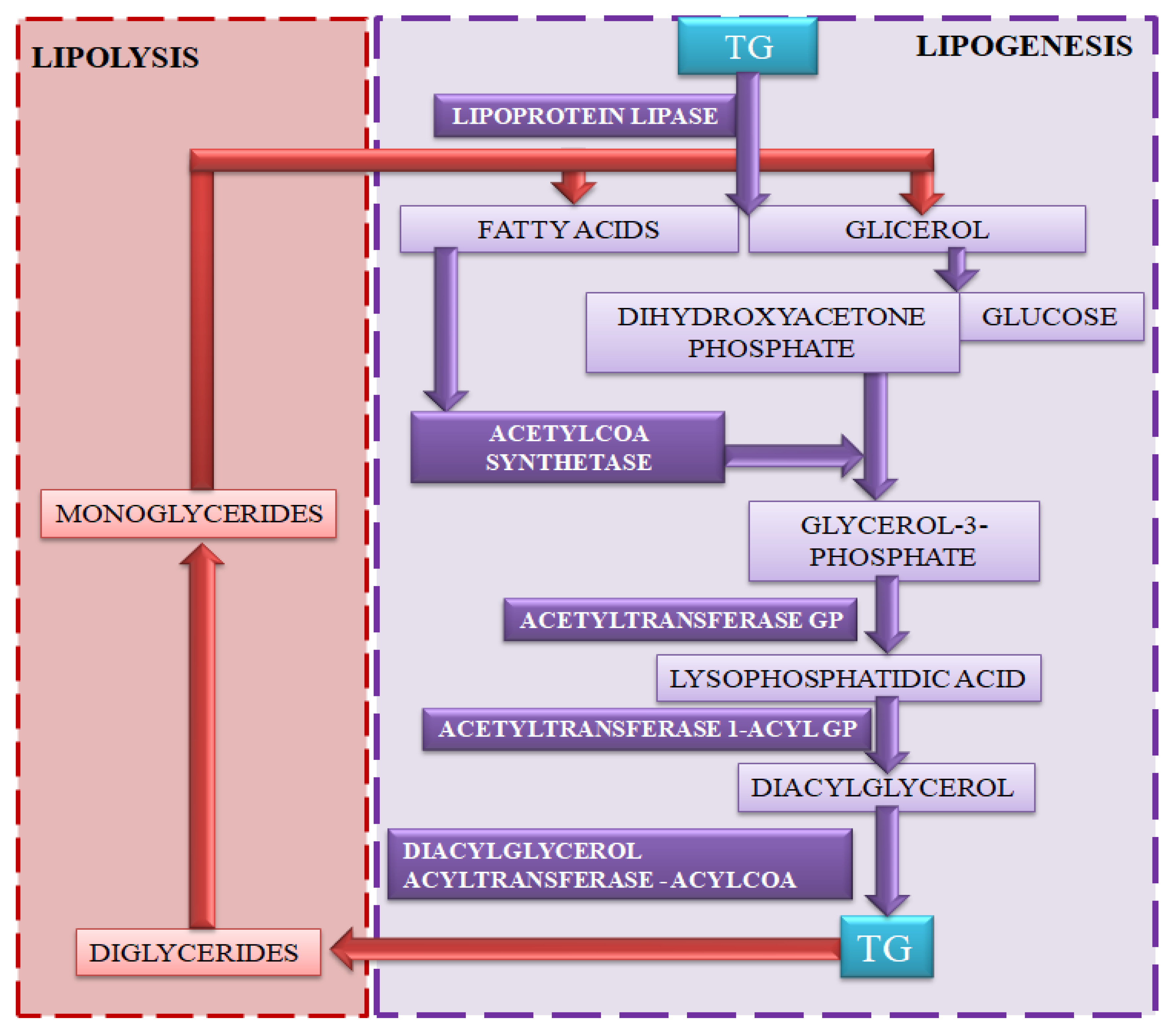
4.1. Lipogenesis
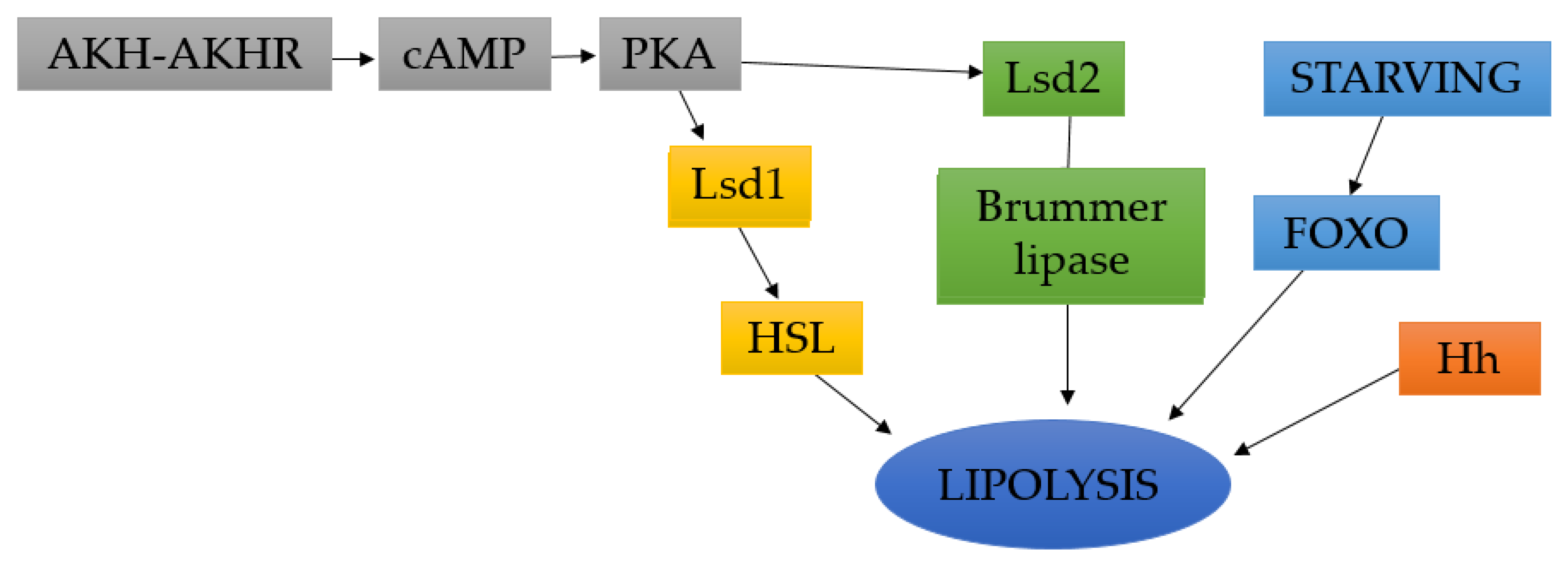
4.2. Lipolysis
4.3. Lipid Metabolism under Various Environmental Conditions
5. Role and Metabolism of Carbohydrates
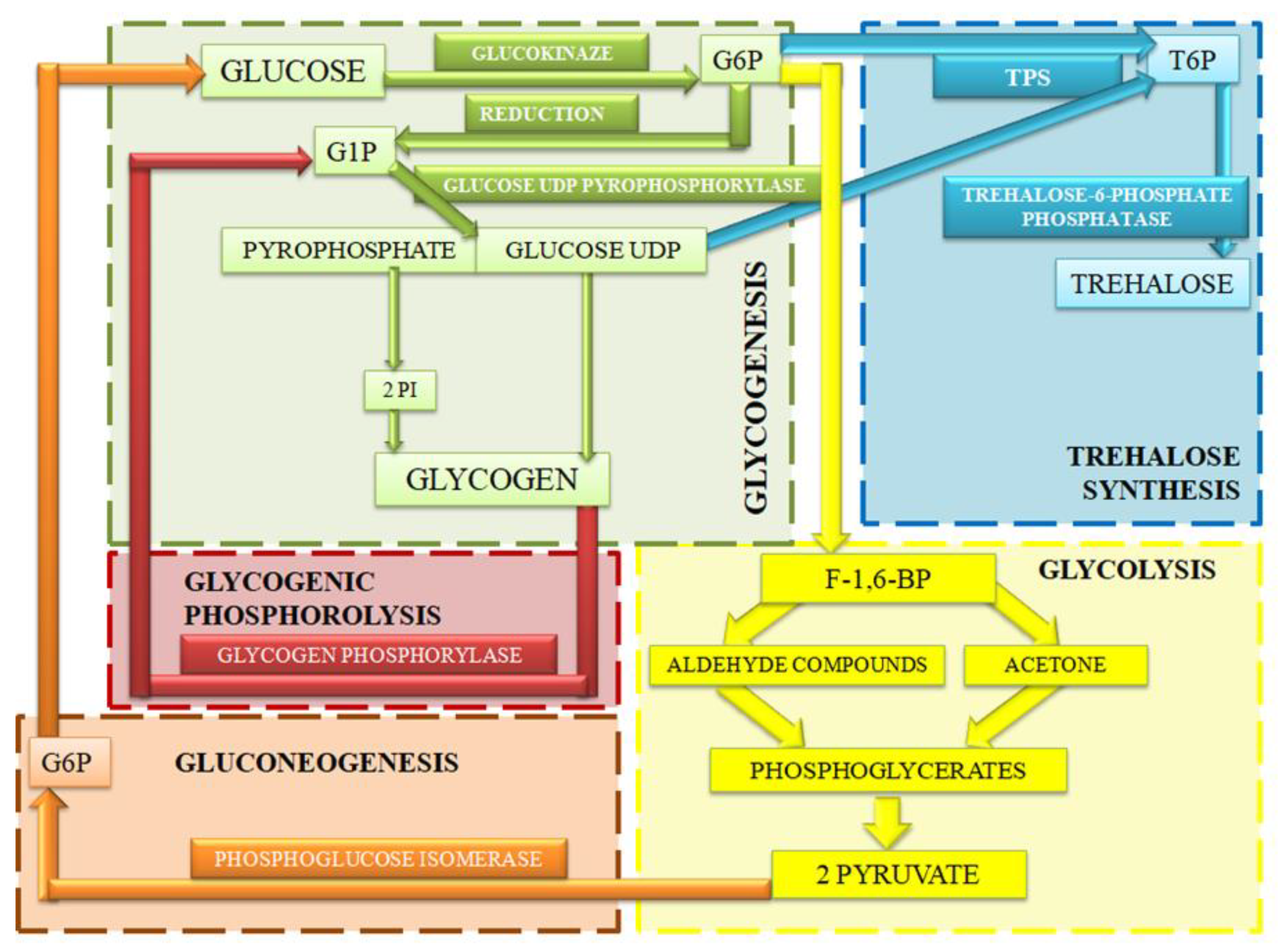
5.1. Glycogenic Phosphorolysis
5.2. Glycogenesis
5.3. Glycolysis
5.4. Gluconeogenesis
5.5. Trehalose Synthesis
6. Role and Metabolism of Proteins
6.1. Urea Cycle
6.2. Proline
6.3. Protein Sensors
7. Additional Metabolic Sensors
8. Hormone Effect on Fat Body
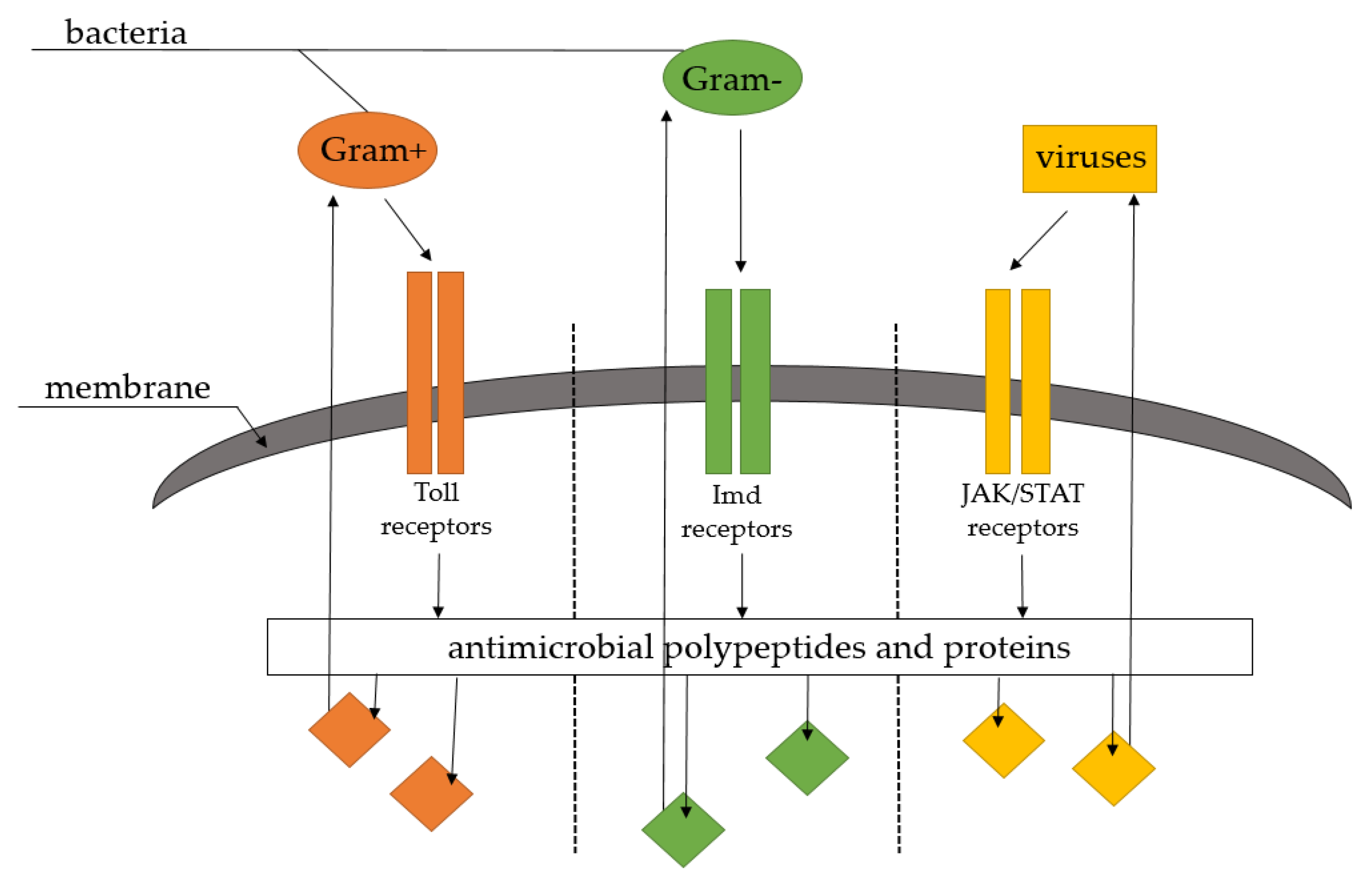
9. Role in Immunity
9.1. Lysozyme
9.2. Lectins
9.3. Cecropins
9.4. Attacin
9.5. Defensin
9.6. Apidicin and Abycin in Apidae Insects
9.7. Reaction to Toxins
9.8. Vitellogenin
9.9. Immunosenescence
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 20E | 20-hydroxyecdysone |
| 93F | proper name of protein |
| ABC | ATP binding cassette |
| AKH-AKHR | adipokinetic hormone—adipokinetic hormone receptor |
| AMPK | 5′ adenosine monophosphate-activated protein kinase |
| APRP | adipokinetic hormone precursor-related peptide |
| Ast-A | allanostatin-A |
| ATG | autophagy-related proteins |
| ATGL | adipose triglyceride lipase |
| Br-C | Broad-Complex |
| CAC | caspase activity cascade |
| cAMP | cyclic adenosine monophosphate |
| CCha2 | CCHamide-2 |
| Ckd2 | cyclin-dependent kinase 2 |
| Ckd4 | cyclin-dependent kinase 4 |
| CRz | co-razonin |
| DG | diglyceride |
| DH-BPAN | diapause hormone-pheromone biosynthesis activating neuropeptide |
| DILP1-8 | insulin-like peptides in Drosophilia melanogaster |
| E93 | ecdysone-induced protein 93 |
| Ecd | ecdysteroid |
| EcR/USP | ecdysone receptor/ultraspiracle protein |
| F-1,6-BP | fructose-1,6-bisphosphate |
| FBS | fat body signal |
| FIT | female-specific independent transformer |
| FOXO | Forkhead box-containing transcription factors |
| G1P | glucose-1-phosphate |
| G3P | glycerol-3-phosphate |
| G6P | glucose-6-phosphate |
| GABA | gamma-aminobutyric acid |
| Gmd | Grindelwald receptor |
| GP | glycogen phosphorylase |
| GST | glutathione-S-transferase |
| HDL | high-density lipoprotein |
| Hh | hedgehog signaling |
| HSL | hormone-sensitive lipase |
| IIS | insulin/IGF signaling |
| ILP6 | insulin-like peptide 6 |
| ILP | insulin-like peptide |
| IP3 | inositol trisphosphate |
| IPC | insulin producing cell |
| JAK/STAT | Janus kinase/signal transducers and activator of transcriptio |
| JH | juvenile hormone |
| Kr-h1 | Krüppel homolog |
| LDL | low-density lipoprotein |
| Lk | leucokin |
| LKB1 | LKB1 kinase |
| Lsd1 | lysine-specific histone demethylase 1A |
| Lsd2 | lysine-specific histone demethylase 1B |
| Lst | limostatin |
| MG | monoglyceride |
| Mmps | metalloproteinase |
| MT | Malpighian tubule system |
| Myc | proper name of gene regulator |
| NPF | neuropeptide F |
| PAMP | pathogen-associated molecular pattern |
| PAT | phosphinothricin acetyltransferase |
| PCD | programmed cell death |
| PGE 2 | prostaglandin receptor |
| PGRP | peptidoglycan recognition protein |
| PKA | protein kinase A |
| PRR | pattern recognition receptor |
| ROS | reactive oxygen species |
| Sir2 | sirtuin homolog |
| sNPF | short neuropeptide F |
| T6P | trehalose-6-phosphate |
| TG | triglyceride |
| TGL | triglyceride lipase |
| Tk | tahykinin |
| Toll | proper name of Toll group receptors |
| TORC1 | transducer of regulated CREB activity 1 |
| TPP | trehalose-6-phosphate phosphatase |
| TPS | trehalose-6-phosphate synthase |
| UDP glucose | uridine diphosphate glucose |
| UGT | UDP gluconosyl transferase |
| Upd2 | unpaired 2 |
| VDVL | very low-density lipoprotein |
References
- ITIS Report: Insecta. Available online: https://www.itis.gov/servlet/SingleRpt/SingleRpt?search_topic=TSN&search_value=99208#null (accessed on 5 May 2021).
- Keeley, L. Biochemistry and physiology of the insect fat body. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Pergamon: New York, NY, USA, 1985; Volume 3, pp. 211–228. [Google Scholar]
- Kritsky, G. The Tears of Re: Beekeeping in Ancient Egypt; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Klatt, B.K.; Holzschuh, A.; Westphal, C.; Clough, Y.; Smit, I.; Pawelzik, E.; Tscharntke, T. Bee pollination improves crop quality, shelf life and commercial value. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132440. [Google Scholar] [CrossRef]
- Law, J.; Wells, M. Insects as biochemical models. J. Biol. Chem. 1989, 264, 16335–16338. [Google Scholar] [CrossRef]
- Manohar, P.; Nachimuthu, R.; Lopes, B.S. The therapeutic potential of bacteriophages targeting gram-negative bacteria using Galleria mellonella infection model. BMC Microbiol. 2018, 18, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Rigueiro, J.; Viney, C.; Llorca, J.; Elices, M. Silkworm silk as an engineering material. J. Appl. 1998, 70, 2439–2447. [Google Scholar] [CrossRef]
- Srivastava, S.K.; Babu, N.; Pandey, H. Traditional insect bioprospecting–As human food and medicine. Indian J. Tradit. Knowl. 2009, 8, 485–494. [Google Scholar]
- Yamamoto, M.; da Silva, C.I.; Augusto, S.C.; Barbosa, A.A.A.; Oliveira, P.E. The role of bee diversity in pollination and fruit set of yellow passion fruit (Passiflora edulis forma flavicarpa, Passifloraceae) crop in Central Brazil. Apidologie 2012, 43, 515–526. [Google Scholar] [CrossRef] [Green Version]
- Bindroo, B.; Manthira Moorthy, S. Genetic divergence, implication of diversity, and conservation of silkworm, Bombyx mori. Int. J. Biodivers. 2014, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chobotow, J.; Strachecka, A. Morfologia i funkcje ciała tłuszczowego owadów z uwzględnieniem pszczoły miodnej Apis mellifera L. Med. Veter. 2013, 69, 712–715. [Google Scholar]
- Beenakkers, A.M.T. The influence of corpus allatum and corpus cardiacum on lipid metabolism in Locusta migratoria. Gen. Comp. Endocrinol. 1969, 13, 492. [Google Scholar]
- Beenakkers, A.; Vanderhorst, D.; Vanmarrewijk, W. Insect flight metabolism. Insect Biochem. 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolezal, T.; Krejcova, G.; Bajgar, A.; Nedbalova, P.; Strasser, P. Molecular regulations of metabolism during immune response in insects. Insect Biochem. Mol. Biol. 2019, 109, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Mirth, C.; Riddiford, L. Size assessment and growth control: How adult size is determined in insects. BioEssays 2007, 29, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Roma, G.C.; Bueno, O.C.; Camargo-Mathias, M.I. Morpho-physiological analysis of the insect fat body: A review. Micron 2010, 41, 395–401. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, V.P.; Cruz-Landim, C. Morphology and function of insect fat body cells: A review. Biociências 2003, 11, 195–205. [Google Scholar]
- Chapman, R.F. The Insect: Structure and Function; Cambridge University Press: Cambridge, UK, 1998. [Google Scholar]
- Gade, G. Regulation of intermediary metabolism and water balance of insects by neuropeptides. Annu. Rev. Entomol. 2004, 49, 93–113. [Google Scholar] [CrossRef]
- Dean, R.; Collins, J.; Locke, M. Structure of the fat body. In Comprehensive Insect Physiology, Biochemistry, and Pharmacology; Pergamon: New York, NY, USA, 1985; pp. 155–210. [Google Scholar]
- Fruttero, L.L.; Leyria, J.; Moyetta, N.R.; Ramos, F.O.; Settembrini, B.P.; Canavoso, L.E. The fat body of the hematophagous insect, Panstrongylus megistus (Hemiptera: Reduviidae): Histological features and participation of the β-chain of ATP synthase in the lipophorin-mediated lipid transfer. J. Insect Sci. 2019, 19, 16. [Google Scholar] [CrossRef]
- Gutierrez, E.; Wiggins, D.; Fielding, B.; Gould, A. Specialized hepatocyte-like cells regulate Drosophila lipid metabolism. Nature 2007, 445, 275–280. [Google Scholar] [CrossRef]
- Vafopoulou-Mandalos, X.; Laufer, H. Tissue-specificity of hemoglobin synthesis: Localization of heme synthesis in the subepidermal fat body of Chironomus thummi (Diptera). Arch. Insect Biochem. Physiol. 1983, 1, 191–197. [Google Scholar] [CrossRef]
- Willott, E.; Bew, L.; Nagle, R.; Wells, M. Sequential structural changes in the fat body of the tobacco hornworm, Manduca sexta, during the fifth larval stadium. Tissue Cell 1988, 20, 635–643. [Google Scholar] [CrossRef]
- Zhang, J.H.; Yu, N.; Xu, X.X.; Liu, Z.W. Community structure, dispersal ability and functional profiling of microbiome existing in fat body and ovary of the brown planthopper, Nilaparvata lugens. Insect Sci. 2019, 26, 683–694. [Google Scholar] [CrossRef]
- Haunerland, N.; Shirk, P. Regional and functional differentiation in the insect fat body. Annu. Rev. Entomol. 1995, 40, 121–145. [Google Scholar] [CrossRef]
- Krupp, J.J.; Levine, J.D. Dissection of oenocytes from adult Drosophila melanogaster. J. Vis. Exp. 2010, 41. [Google Scholar] [CrossRef] [PubMed]
- Rizki, T. Intracellular localization of kynurenine in the fat body of Drosophila. J. Cell Biol. 1961, 9, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Rizki, T.; Rizki, R. Cytodifferentiation in the rosy mutant of Drosophila melanogaster. J. Cell Biol. 1962, 12, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, B.A.; Orr-Weaver, T.L. Endoreplication cell cycles: More for less. Cell 2001, 105, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Yang, X.; Xi, Y. Fat body remodeling and homeostasis control in Drosophila. Life Sci. 2016, 167, 22–31. [Google Scholar] [CrossRef]
- Yamanaka, N.; Rewitz, K.F.; O’Connor, M.B. Ecdysone control of developmental transitions: Lessons from Drosophila research. Annu. Rev. Entomol. 2013, 58, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Bond, N.D.; Nelliot, A.; Bernardo, M.K.; Ayerh, M.A.; Gorski, K.A. βFTZ-F1 and Matrix metalloproteinase 2 are required for fat-body remodeling in Drosophila. Dev. Biol. 2011, 360, 286–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Q.; Liu, Y.; Liu, H.; Li, S. Mmp1 and Mmp2 cooperatively induce Drosophila fat body cell dissociation with distinct roles. Sci. Rep. 2014, 4, 7535. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L. 20-Hydroxyecdysone (20E) primary response gene E93 modulates 20E signaling to promote Bombyx larval-pupal metamorphosis. J. Biol. Chem. 2015, 290, 27370–27383. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lu, Y.X.; Liu, J.; Yang, C.; Feng, Q.L.; Xu, W.H. A regulatory pathway, ecdysonetranscription factor relish-cathepsin L, is involved in insect fat body dissociation. PLoS Genet. 2013, 9, e1003273. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, H.D.; Baehrecke, E.H. Distinct death mechanisms in Drosophila development. Curr. Opin. Cell Biol. 2010, 22, 889–895. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Tian, L.; Guo, X.; Li, K.; Li, J. BmATG5 and BmATG6 mediate apoptosis following autophagy induced by 20-hydroxyecdysone or starvation. Autophagy 2016, 12, 381–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.Y.; Neufeld, T.P. An Atg1/Atg13 complexwithmultiple roles in TOR-mediated autophagy regulation. Mol. Biol. Cell 2009, 20, 2004–2014. [Google Scholar] [CrossRef] [Green Version]
- Delanoue, R.; Slaidina, M.; L’eopold, P. The steroid hormone ecdysone controls systemic growth by repressing dMyc function in Drosophila fat cells. Dev. Cell 2010, 18, 1012–1021. [Google Scholar] [CrossRef] [PubMed]
- Lippai, M.; Csik’os, G.; Maroy, P.; Luk’acsovich, T.; Juh’asz, G.; Sass, M. SNF4Aγ, the Drosophila AMPK γsubunit is required for regulation of developmental and stress-induced autophagy. Autophagy 2008, 4, 476–486. [Google Scholar] [CrossRef]
- Liu, H.; Jia, Q.; Tettamanti, G.; Li, S. Balancing crosstalk between 20-hydroxyecdysone-induced autophagy and caspase activity in the fat body during Drosophila larval-prepupal transition. Insect Biochem. Mol. Biol. 2013, 43, 1068–1078. [Google Scholar] [CrossRef]
- Scott, R.C.; Schuldiner, O.; Neufeld, T.P. Role and regulation of starvation-induced autophagy in the Drosophila fat body. Dev. Cell. 2004, 7, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ding, Y.; Lu, X.; Geng, D.; Li, S.; Raikhel, A.S.; Zou, Z. The ecdysone-induced protein 93 is a key factor regulating gonadotrophic cycles in the adult female mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2021, 118, e2021910118. [Google Scholar] [CrossRef]
- Agrawal, N.; Delanoue, R.; Mauri, A.; Basco, D.; Pasco, M. The Drosophila TNF Eiger is an adipokine that acts on insulin-producing cells to mediate nutrient response. Cell Metab. 2016, 23, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Frei, C.; Galloni, M.; Hafen, E.; Edgar, B.A. The Drosophila mitochondrial ribosomal protein mRpL12 is required for Cyclin D/Cdk4-driven growth. EMBO J. 2005, 24, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Lohnson, M.; Butterworth, F. Maturation and aging of adult fat bodyand oenocytes in Drosophila as revealedby light microscopic morphometry. J. Morphol. 1985, 184, 51–59. [Google Scholar] [CrossRef]
- Li, S.; Yu, X.; Feng, Q. Fat body biology in the last decade. Annu. Rev. Entomol. 2019, 64, 315–333. [Google Scholar] [CrossRef]
- Yuan, D.; Zhou, S.; Liu, S.; Li, K.; Zhao, H.; Long, S.; Liu, H.; Xie, Y.; Su, Y.; Yu, F.; et al. The AMPK-PP2A axis in insect fat body is activated by 20-hydroxyecdysone to antagonize insulin/IGF signaling and restrict growth rate. Proc. Natl. Acad. Sci. USA 2020, 117, 9292–9301. [Google Scholar] [CrossRef]
- Almeida-Oliveira, F.; Tuthill, B.F., II; Gondim, K.C.; Majerowicz, D.; Musselman, L.P. dHNF4 regulates lipid homeostasis and oogenesis in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2021, 133, 103569. [Google Scholar] [CrossRef]
- Canavoso, L.; Jouni, Z.; Karnas, K.; Pennington, J.; Wells, M. Fat metabolism in insects. Annu. Rev. Nutr. 2001, 21, 23–46. [Google Scholar] [CrossRef]
- Gilby, A. Lipids and their metabolism in insects. Annu. Rev. Entomol. 1965, 10, 141–160. [Google Scholar] [CrossRef]
- Patel, R.T.; Soulages, J.L.; Hariharasundaram, B.; Arrese, E.L. Activation of the lipid droplet controls the rate of lipolysis of triglycerides in the insect fat body. J. Biol. Chem. 2005, 280, 22624–22631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasaemle, D. Thematic review series: Adipocyte biology. The perilipin family of structural lipid droplet proteins: Stabilization of lipid droplets and control of lipolysis. J. Lipid Res. 2007, 48, 2547–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronke, S.; Beller, M.; Fellert, S.; Ramakrishnan, H.; Jackle, H.; Kuhnlein, R. Control of fat storage by a Drosophila PAT domain protein. Curr. Biol. 2013, 13, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.; Xiang, Y.; Chen, H.; Liu, Z.; Gronke, S. Opposite and redundant roles of the two Drosophila perilipins in lipid mobilization. J. Cell Sci. 2012, 125, 3568–3577. [Google Scholar] [CrossRef] [Green Version]
- Soulages, J.; Wells, M. Lipophorin: The structure of an insect lipoprotein and its role in lipid transport in insects. Adv. Protein Chem. 1994, 45, 371–415. [Google Scholar] [PubMed]
- Defferrari, M.S.; Orchard, I.; Lange, A.B. Identification of the first insulin-like peptide in the disease vector Rhodnius prolixus: Involvement inmetabolic homeostasis of lipids and carbohydrates. Insect Biochem. Mol. Biol. 2016, 70, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Gronke, S.; Mildner, A.; Fellert, S.; Tennagels, N.; Petry, S. Brummer lipase is an evolutionary conserved fat storage regulator in Drosophila. Cell Metab. 2005, 1, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Gronke, S.; Muller, G.; Hirsch, J.; Fellert, S.; Andreou, A. Dual lipolytic control of body fat storage and mobilization in Drosophila. PLoS Biol. 2007, 5, e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.S.; Liu, Y.; Zhou, S.; Li, K.; Tian, L.; Li, S. 20-Hydroxyecdysone-induced transcriptional activity of FoxO upregulates brummer and acid lipase-1 and promotes lipolysis in Bombyx fat body. Insect Biochem. Mol. Biol. 2013, 43, 829–838. [Google Scholar] [CrossRef]
- Rajan, A.; Perrimon, N. Drosophila cytokine Unpaired 2 regulates physiological homeostasis by remotely controlling insulin secretion. Cell 2012, 151, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Toprak, U.; Hegedus, D.; Doğan, C.; Güney, G. A journey into the world of insect lipid metabolism. Arch. Insect Biochem. Physiol. 2020, 104, e21682. [Google Scholar] [CrossRef]
- Jimenez-Sanchez, M.; Menzies, F.M.; Chang, Y.Y.; Simecek, N.; Neufeld, T.P.; Rubinsztein, D.C. The Hedgehog signalling pathway regulates autophagy. Nat. Commun. 2012, 3, 1200. [Google Scholar] [CrossRef] [Green Version]
- Reis, T.; Van Gilst, M.R.; Hariharan, I.K. A buoyancy-based screen of Drosophila larvae for fat-storage mutants reveals a role for Sir2 in coupling fat storage to nutrient availability. PLoS Genet. 2010, 6, e1001206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugrankar, R.; Liu, Y.L.; Provaznik, J.; Schmitt, S.; Lehmann, M. Lipin is a central regulator of adipos tissue development and function in Drosophila melanogaster. Mol. Cell. Biol. 2011, 31, 1646–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, D. Prostaglandins and other eicosanoids in insects: Biological significance. Annu. Rev. Entomol. 2006, 51, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Mead, J.F. Lipids: Chemistry, Biochemistry, and Nutrition; Plenum Press: New York, NY, USA, 1986. [Google Scholar]
- Athenstaedt, K.; Daum, G. The life cycle of neutral lipids: Synthesis, storage and degradation. Cell. Mol. Life Sci. 2006, 63, 1355–1369. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.Y.; Lucas, H.; Sagili, R.; Cha, D.H.; Lee, J.C. Effect of erythritol on Drosophila suzukii (Diptera: Drosophilidae) in the presence of naturally-occurring sugar sources, and on the survival of Apis mellifera (Hymenoptera: Apidae). J. Econ. Entomol. 2019, 112, 981–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sztalryd, C.; Xu, G.; Dorward, H.; Tansey, J.T.; Contreras, J.A. Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. J. Cell Biol. 2013, 161, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Doğan, C.; Hänniger, S.; Heckel, D.G.; Coutu, C.; Hegedus, D.D.; Crubaugh, L.; Groves, R.L.; Mutlu, D.A.; Suludere, Z.; Bayram, Ş.; et al. Characterization of calcium signaling proteins from the fat body of the Colorado Potato Beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae): Implications for diapause and lipid metabolism. Insect Biochem. Mol. Biol. 2021, 133, 103549. [Google Scholar] [CrossRef]
- Vardanis, A. Glycogen synthesis in the insect fat body. BBA-Spec. Sect. Enzymol. Subj. 1963, 73, 565–573. [Google Scholar] [CrossRef]
- Lockey, K.H. Lipids of the insect cuticle: Origin, composition and function. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1988, 89, 595–645. [Google Scholar] [CrossRef]
- Storey, K.B. Organic solutes in freezing tolerance. Comp. Biochem. Phys. A 1997, 117, 319–326. [Google Scholar] [CrossRef]
- Steele, J. Glycogen-phosphorylase in insects. Insect Biochem. 1982, 12, 131–134. [Google Scholar] [CrossRef]
- Goodman, H.M. Growth hormone and the metabolism of carbohydrate and lipid in adipose tissue. N. Y. Acad. Sci. 1968, 148, 419–440. [Google Scholar] [CrossRef] [PubMed]
- Young, F.G. Glycogen and the metabolism of carbohydrate. Lancet 1936, 231, 237–302. [Google Scholar] [CrossRef]
- Meyer-Fernandes, J.; Clark, C.; Gondim, K.; Wells, M. Fat body fructose-2,6-bisphosphate content and phosphorylase activity correlate with changes in hemolymph glucose concentration during fasting and re-feeding in larval Manduca sexta. Insect Biochem. Mol. Biol. 2001, 31, 165–170. [Google Scholar] [CrossRef]
- Sacktor, B. Biochemical adaptations for flight in the insect. In Biochemical Society Symposium; Portland Press: London, UK, 1976; pp. 111–131. [Google Scholar]
- Stryer, L. Biochemia. Wydawnictwo Naukowe PWN. 2003. Available online: https://idoc.pub/documents/biochemia-stryer-d2nv6pwg304k (accessed on 7 June 2021).
- Exton, J.H. Gluconeogenesis. Metabolism 1972, 21, 945–990. [Google Scholar] [CrossRef]
- Garcia, C.K.; Goldstein, J.L.; Pathak, R.K.; Anderson, R.G.; Brown, M.S. Molecular characterization of a membrane transporter for lactate, pyruvate, and other monocarboxylates: Implications for the Cori cycle. Cell 1994, 76, 865–873. [Google Scholar] [CrossRef]
- Hatting, M.; Tavares, C.D.; Sharabi, K.; Rines, A.K.; Puigserver, P. Insulin regulation of gluconeogenesis. Ann. N. Y. Acad. Sci. 2018, 1411, 21. [Google Scholar] [CrossRef]
- Thompson, S. Trehalose: The insect ‘blood’ sugar. Adv. Insect Physiol. 2003, 31, 205–285. [Google Scholar]
- Friedman, S. Trehalose regulation, one aspect of metabolic homeostasis. Annu. Rev. Entomol. 1978, 23, 389–407. [Google Scholar] [CrossRef]
- Gade, G.; Auerswald, L. Beetles’ choice—Proline for energy output: Control by AKHs. Comp. Biochem. Physiol. B 2002, 132, 117–129. [Google Scholar] [CrossRef]
- Tojo, S.; Betchaku, T.; Ziccardi, V.; Wyatt, G. Fat body protein granules andstorage proteins in the silkmoth, Hyalophora cecropia. J. Cell Biol. 1978, 78, 823–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bursell, E. The role of proline in energy metabolism. In Energy Metabolism in Insects; Pergamon: New York, NY, USA, 1981; pp. 135–154. [Google Scholar]
- Locke, M. The structure and development of the vacuolar system in the fat body of insects. In Insect Ultrastructure; Springer: Boston, MA, USA, 1984; pp. 151–197. [Google Scholar]
- Locke, M.; McMahon, I. The origin and fate of microbodies in the fat body of an insect. J. Cell Bioi. 1971, 48, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Buckner, J. 20-Hydroxyecdysone-induced storage of uric acid in Manduca sexta fat body. Am. Zool. 1980, 20, 939. [Google Scholar]
- Nijhout, H.; Williams, C. Control of moulting and metamorphosis in the tocbacco hornworm, Manduca sexta (L.); growth of the last-instar larva andthe decision to pupate. J. Exp. Biol. 1974, 61, 481–491. [Google Scholar] [CrossRef]
- Scholz, R.W.; Featherston, W.R. Effect of alterations in protein intake on liver xanthine dehydrogenase in the chick. J. Nutr. 1968, 95, 271–277. [Google Scholar] [CrossRef] [Green Version]
- Pimley, R.; Langley, P. Hormone stimulated lipolysis and proline synthesis in the fat body of the adult tsetse fly, Glossina morsitans. J. Insect Physiol. 1982, 28, 781–789. [Google Scholar] [CrossRef]
- Colombani, J.; Raisin, S.; Pantalacci, S.; Radimerski, T.; Montagne, J.; L’eopold, P. A nutrient sensor mechanism controls Drosophila growth. Cell 2013, 114, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Delanoue, R.; Meschi, E.; Agrawal, N.; Mauri, A.; Tsatskis, Y. Drosophila insulin release is triggered by adipose Stunted ligand to brain Methuselah receptor. Science 2016, 353, 1553–1556. [Google Scholar] [CrossRef] [Green Version]
- G’eminard, C.; Rulifson, E.J.; L’eopold, P. Remote control of insulin secretion by fat cells in Drosophila. Cell Metab. 2009, 10, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Sano, H.; Nakamura, A.; Texada, M.J.; Truman, J.W.; Ishimoto, H.; Kamikouchi, A. The nutrient-responsive hormone CCHamide-2 controls growth by regulating insulin-like peptides in the brain of Drosophila melanogaster. PLoS Genet. 2015, 11, e1005209. [Google Scholar]
- Sun, J.; Liu, C.; Bai, X.; Li, X.; Li, J. Drosophila FIT is a protein-specific satiety hormone essential for feeding control. Nat. Commun. 2017, 8, 14161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liu, H.; Liu, S.; Wang, S.; Jiang, R.J.; Li, S. Hormonal and nutritional regulation of insect fat body development and function. Arch. Insect Biochem. Physiol. 2009, 71, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Toprak, U. The role of peptide hormones in insect lipid metabolism. Front. Physiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Gulia, M.; Clark, K.D.; Dhara, A.; Crim, J.W.; Strand, M.R. Two insulin-like peptide family members from the mosquito Aedes aegypti exhibit differential biological and receptor binding activities. Mol. Cell. Endocrinol. 2010, 328, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vroemen, S.F.; Van der Horst, D.J.; Van Marrewijk, W.J. New insights into adipokinetic hormone signaling. Mol. Cell. Endocrinol. 1998, 141, 7–12. [Google Scholar] [CrossRef]
- Arrese, E.L.; Mirza, S.; Rivera, L.; Howard, A.D.; Chetty, P.S.; Soulages, J.L. Expression of lipid storage droplet protein-1 may define the role of AKH as a lipid mobilizing hormone in Manduca sexta. Insect Biochem. Mol. Biol. 2008, 38, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Alves-Bezerra, M.; De Paula, I.F.; Medina, J.M.; Silva-Oliveira, G.; Medeiros, J.S.; Gäde, G. Adipokinetic hormone receptor gene identification and its role in triacylglycerol metabolism in the blood-sucking insect Rhodnius. Insect Biochem. Mol. Biol. 2016, 69, 51–60. [Google Scholar] [CrossRef]
- Milde, J.J.; Ziegler, R.; Wallstein, M. Adipokinetic hormone stimulates neurones in the insect central nervous system. J. Exp. Biol. 1995, 198, 1307–1311. [Google Scholar] [CrossRef]
- Scarborough, R.M.; Jamieson, G.C.; Kalish, F.; Kramer, S.J.; McEnroe, G.A.; Miller, C.A. Isolation and primary structure of two peptides with cardioacceleratory and hyperglycemic activity from the corpora cardiaca of Periplaneta americana. Proc. Natl. Acad. Sci. USA 1984, 81, 5575–5579. [Google Scholar] [CrossRef] [Green Version]
- Socha, R.; Kodrík, D.; Zemek, R. Adipokinetic hormone stimulates insect locomotor activity. Naturwissenschaften 1999, 88, 85–86. [Google Scholar] [CrossRef]
- Strachecka, A.; Chobotow, J.; Paleolog, J.; Łoś, A.; Schulz, M.; Teper, D.; Kucharczyk, H.; Grzybek, M. Insights into the biochemical defence and methylation of the solitary bee Osmia rufa L: A foundation for examining eusociality development. PLoS ONE 2017, 12, e0176539. [Google Scholar]
- Brown, M.R.; Clark, K.D.; Gulia, M.; Zhao, Z.; Garczynski, S.F.; Crim, J.W. An insulin-like peptide regulates egg maturation and metabolism in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2008, 105, 5716–5721. [Google Scholar] [CrossRef] [Green Version]
- Kannan, K.; Fridell, Y.W.C. Functional implications of Drosophila insulin-like peptides in metabolism, aging, and dietary restriction. Front. Physiol. 2013, 4, 288. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, I.; Moto, K.; Sakurai, S.; Iwami, M. A novel member of the bombyxin gene family: Structure and expression of bombyxin G1 gene, an insulin-related peptide gene of the silkmoth Bombyx mori. Dev. Genes Evol. 1998, 208, 407–410. [Google Scholar] [CrossRef]
- Majerowicz, D.; Gondim, K.C. Insect lipid metabolism: Insights into gene expression regulation. In Recent Trends in Gene Expression; Mandal, S.S., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 147–189. [Google Scholar]
- Choi, Y.J.; Lee, G.; Hall, J.C.; Park, J.H. Comparative analysis of Corazonin-encoding genes (Crz’s) in Drosophila species and functional insights into Crz-expressing neurons. J. Comp. Neurol. 2005, 482, 372–385. [Google Scholar] [CrossRef]
- Gospocic, J.; Shields, E.J.; Glastad, K.M.; Lin, Y.; Penick, C.A.; Yan, H. The neuropeptide corazonin controls social behavior and caste identity in ants. Cell 2017, 170, 748–759. [Google Scholar] [CrossRef]
- Kim, Y.J.; Spalovská-Valachová, I.; Cho, K.H.; Zitnanova, I.; Park, Y.; Adams, M.E. Corazonin receptor signaling in ecdysis initiation. Proc. Natl. Acad. Sci. USA 2004, 101, 6704–6709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayler, T.D.; Pacheco, D.A.; Hergarden, A.C.; Murthy, M.; Anderson, D.J. A neuropeptide circuit that coordinates sperm transfer and copulation duration in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 20697–20702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žitòan, D.; Daubnerová, I. Crustacean cardioactive peptide. In Handbook of Hormones; Takei, Y., Ando, H., Tsutsui, K., Eds.; Academic Press: Oxford, UK, 2016; pp. 442–443. [Google Scholar]
- Ren, G.R.; Hauser, F.; Rewitz, K.F.; Kondo, S.; Engelbrecht, A.F.; Didriksen, A.K. CCHamide-2 is an orexigenic brain-gut peptide in Drosophila. PLoS ONE 2015, 10, e0133017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veenstra, J.A.; Agricola, H.J.; Sellami, A. Regulatory peptides in fruit fly midgut. Cell Tissue Res. 2018, 334, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Gliński, Z.; Jarosz, J. Immunobiologia Pszczoły Miodnej; Uniwersytet Przyrodniczy w Lublinie: Lublin, Poland, 1995; ISBN 83-86761-12-1. [Google Scholar]
- Strachecka, A.; Łoś, A.; Filipczuk, J.; Schulz, M. Indywidualne i społeczne mechanizmy odporności pszczoły miodnej. J. Med. Veter. 2018, 74, 426–433. [Google Scholar]
- Hanson, M.A.; Dostálová, A.; Ceroni, C.; Poidevin, M.; Kondo, S. Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. eLife 2019, 8, e44341. [Google Scholar] [CrossRef]
- Hanson, M.A.; Lemaitre, B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr. Opin. Immunol. 2020, 62, 22–30. [Google Scholar] [CrossRef]
- Takov, D.I.; Ostoich, P.V.; Tchorbanov, A.I.; Pilarska, D.K. Order Diptera as a model in the studies of insect immunity: A review. Turk. J. Zool. 2020, 44, 481–489. [Google Scholar] [CrossRef]
- Stroschein-Stevenson, S.L.; Foley, E.; O’Farrell, P.H.; Johnson, A.D. Identification of Drosophila gene products required for phagocytosis of Candida albicans. PLoS Biol. 2006, 4, e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, S.A.; Wasserman, S.A. Conventional and non-conventional Drosophila Toll signaling. Dev. Comp. Immunol. 2014, 42, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Manniello, M.D.; Moretta, A.; Salvia, R.; Scieuzo, C.; Lucchetti, D.; Vogel, H.; Sgambato, A.; Falabella, P. Insect antimicrobial peptides: Potential weapons to counteract the antibiotic resistance. Cell. Mol. Life Sci. 2021, 78, 4259–4282. [Google Scholar]
- Myllymaki, H.; Valanne, S.; Rämet, M. The Drosophila Imd signaling pathway. J. Immunol. 2014, 192, 3455–3462. [Google Scholar] [CrossRef] [Green Version]
- Nunes, C.; Sucena, É.; Koyama, T. Endocrine regulation of immunity in insects. FEBS J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.J.; Zhan, M.Y.; Pan, Y.M.; Liu, S.; Yang, P.J. Immune functions of insect βGRPs and their potential application. Dev. Comp. Immunol. 2017, 83, 80–88. [Google Scholar] [CrossRef]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grizanova, E.V.; Coates, C.J.; Butt, T.M.; Dubovskiy, I.M. RNAi-mediated suppression of insect metalloprotease inhibitor (IMPI) enhances Galleria mellonella susceptibility to fungal infection. Dev. Comp. Immunol. 2021, 122, 104126. [Google Scholar] [CrossRef] [PubMed]
- Gajda, E.; Bugla-Płoskońska, G. Lizozym-występowanie w przyrodzie, właściwości biologiczne i możliwości zastosowań. Adv. Clin. Exp. Med. 2014, 68, 1501–1515. [Google Scholar]
- Jollès, J.; Schoentgen, F.; Croizier, G.; Croizier, L.; Jollès, P. Insect lysozymes from three species of Lepidoptera: Their structural relatedness to the C (chicken) type lysozyme. J. Mol. Evol. 1979, 14, 267–271. [Google Scholar] [CrossRef]
- Samakovlis, C.; Kimbrell, D.A.; Kylsten, P.; Engström, Å.; Hultmark, D. The immune response in Drosophila: Pattern of cecropin expression and biological activity. EMBO J. 1990, 9, 2969–2976. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Cociancich, S.; Dimarcq, J.L.; Lambert, J.; Reichhart, J.M.; Hoffmann, D.; Hoffmann, J.A. Insect immunity. Isolation from a coleopteran insect of a novel inducible antibacterial peptide and of new members of the insect defensin family. J. Biol. Chem. 1991, 36, 24520–24525. [Google Scholar] [CrossRef]
- Darkin-Rattray, S.J.; Gurnett, A.M.; Myers, R.W.; Dulski, P.M.; Crumley, T.M.; Allocco, J.J.; Schmatz, D.M. Apicidin: A novel antiprotozoal agent that inhibits parasite histone deacetylase. Proc. Natl. Acad. Sci. USA 1996, 93, 13143–13147. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Ahn, S.J.; Vogel, H.; Heckel, D.G. Comparative analysis of the UDP-glycosyltransferase multigene family in insects. Insect Biochem. Mol. Biol. 2012, 42, 133–147. [Google Scholar] [CrossRef]
- Ketterman, A.J.; Saisawang, C.; Wongsantichon, J. Insect glutathione transferases. Drug Metab. Rev. 2011, 43, 253–265. [Google Scholar] [CrossRef]
- Feyereisen, R. Arthropod CYPomes illustrate the tempo and mode in P450 evolution. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 19–28. [Google Scholar] [CrossRef]
- Grosso, C.G.; Blariza, M.J.; Mougabure-Cueto, G.; Picollo, M.I.; Garcia, B.A. Identification of three cytochrome P450 genes in the Chagas’ disease vector Triatoma infestans: Expression analysis in deltamethrin susceptible and resistant populations. Infect. Genet. Evol. 2016, 44, 459–470. [Google Scholar] [CrossRef]
- Pan, M.L.; Bell, W.J.; Telfer, W.H. Vitellogenic blood protein synthesis by insect fat body. Science 1969, 165, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.C.; Lindström, I.; Lee, J.Y.; Faye, I. Structure and expression of the attacin genes in Hyalophora cecropia. Eur. J. Biochem. 1991, 196, 247–254. [Google Scholar] [CrossRef]
- Valle, D. Vitellogenesis in insects and other groups: A review. Mem. Inst. Oswaldo Cruz 1993, 88, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.; Yang, Y.; Kumar, S.; Lee, D.W.; Bajracharya, P.; Calkins, T.L.; Pietrantonio, P.V. Characterization of the first insect prostaglandin (PGE2) receptor: MansePGE2R is expressed in oenocytoids and lipoteichoic acid (LTA) increases transcript expression. Insect Biochem. Mol. Biol. 2020, 117, 103290. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Stanley, D. Eicosanoid Signaling in Insect Immunology: New Genes and Unresolved Issues. Genes 2021, 12, 211. [Google Scholar] [CrossRef] [PubMed]
- Aw, D.; Silva, A.B.; Palmer, D.B. Immunosenescence: Emerging challenges for an ageing population. Immunology 2007, 120, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Al Baki, M.A.; Lee, J.; Seo, D.Y.; Lee, D.; Kim, Y. The first report of prostacyclin and its physiological roles in insects. Gen. Comp. Endocrinol. 2021, 301, 113659. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skowronek, P.; Wójcik, Ł.; Strachecka, A. Fat Body—Multifunctional Insect Tissue. Insects 2021, 12, 547. https://doi.org/10.3390/insects12060547
Skowronek P, Wójcik Ł, Strachecka A. Fat Body—Multifunctional Insect Tissue. Insects. 2021; 12(6):547. https://doi.org/10.3390/insects12060547
Chicago/Turabian StyleSkowronek, Patrycja, Łukasz Wójcik, and Aneta Strachecka. 2021. "Fat Body—Multifunctional Insect Tissue" Insects 12, no. 6: 547. https://doi.org/10.3390/insects12060547
APA StyleSkowronek, P., Wójcik, Ł., & Strachecka, A. (2021). Fat Body—Multifunctional Insect Tissue. Insects, 12(6), 547. https://doi.org/10.3390/insects12060547