
Endophytic Isaria javanica pf185 Persists after Spraying and Controls Myzus persicae (Hemiptera: Aphididae) and Colletotrichum acutatum (Glomerellales: Glomerellaceae) in Pepper

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Insects and Plants
2.3. Conidial Suspensions
2.4. Experimental Set-Up
2.5. Seedling Inoculation and Management
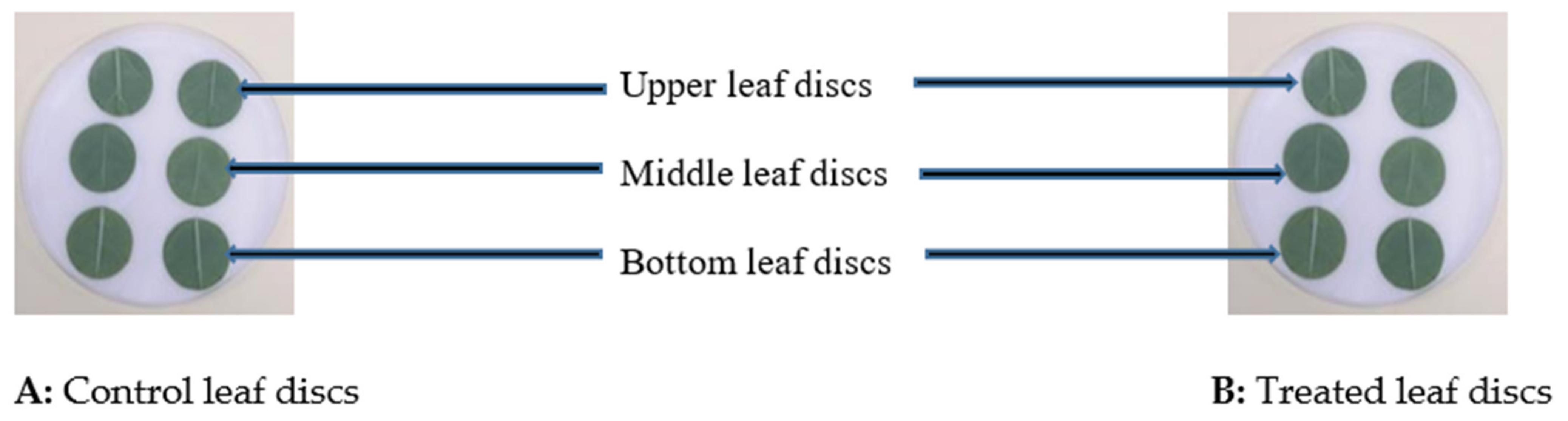
2.6. Leaf Sampling for I. Javanica pf185 Biocontrol Assessments and its Persistence via Isolation
2.7. Effect of I. Javanica pf185 on M. Persicae
2.8. Effect of I. Javanica pf185 on C. Acutatum
2.9. Examining the Endophytic Presence of I. Javanica pf185
- (a)
- Samples of leaf discs were washed under tap water to remove dust and rinsed three times with distilled water. To suppress microorganisms on the leaf surface, the samples were surface-sterilized with 70% ethanol for 30–60 s, followed by a 5% sodium hypochlorite bath for 3 min. The leaves were finally rinsed five times with sterile distilled water and left on sterile tissue paper [37] to dry. Three dried leaf discs from the top, medium, and bottom parts of plants (i.e., nine leaf discs) were each cut using a 0.8 cm-diameter cork borer. Then, the leaf discs were cut with a sterile blade into 1 × 20 mm segments to avoid inoculating new microorganisms. Each leaf segment from the same plant was placed onto a PDA medium supplemented with 500 mg/l chloramphenicol (PDA-C) to suppress bacterial growth [38]. To confirm success of the sterilization, the final rinse with 50 µL distilled water was placed on a PDA medium as a negative control. All plates were incubated in the dark at 25 °C and daily examined for four weeks because growth rate is not identical between taxonomic groups. The success of the leaf piece sterilization was confirmed on Control PDA plates from which no fungal or bacterial growth was observed.
- (b)
- Fungal spores that grew from sterilized leaf surface were extracted from 4 week-old PDA plates by adding 10 mL sterile 0.05% Tween 80® solution, and the spores were filtered through four screens made of cheesecloth to remove hyphae and other particles. The mixture was then vortexed for 3 min to get a homogenous mixture of spores. To start a single spore culture of the different harvested species, using a hemocytometer, each suspension was adjusted to 103/mL as described by Han et al. [34]. Then, the underside of each Petri dish containing fresh PDA was marked at 1 cm × 1 cm with a pen. Through a micropipette 1 μL drop of the suspension was placed above the PDA at the middle of each square and allowed to grow. Between 80–100 drops were placed on each PDA plate weekly to identify the maximum of taxonomic groups present.
- (c)
- The developed fungal colonies were purified through successive transplanting and single spore techniques. After slide culturing, the purified endophytic fungi were identified according to the fungal morphological and micromorphological characteristics as described [39,40,41]. At the end, only the white colonies obtained had been considered for this experiment to eliminate ones that did not resemble I. javanica pf185, which is originally white. The different taxonomic groups isolated were stored at 5 °C in a solution of 40% glycerol and water. Similarly, the endophytic colonization frequency (Endo CF) of the different fungi was calculated individually for the collected samples.
2.10. Weekly Ectophytic Presence of I. Javanica pf185 after Spraying
- (a)
- From leaf samples collected, respectively, at the top, middle, and bottom of each plant, 0.5 cm-diameter leaf discs were cut (three discs from each plant part). The discs were then placed in a tube that contained five milliliters of 0.05% Tween 80® solution and vortexed for 1 min to promote detachment of the adhered spores. Ten milliliters of this solution were transferred onto PDA plates (three plates per sample), which were sealed and incubated at 25 °C. Three days after incubation, the average number of colony-forming units (CFUs) was counted and estimated as CFUs/mm2 per plant. All colonies of I. javanica were microscopically observed and counted to estimate ectophytic colonization frequency (Ecto CF) on the leaf surface per week using the formula:
- (b)
- Then, the spore concentration of the same leaf washed solution was determined through a hemocytometer. The average spore concentration of mixture from each sterilized leaf surface was estimated in conidia/mm3 with three replications from top, middle, and bottom canopy per single plant. All CFUs present on the medium were counted.
- (c)
- Finally, the three leaf discs were vortexed, removed from each tube and placed on filter paper moistened with distilled water in Petri dishes and allowed to dry under a laminar cabinet. After drying, the number of spores remaining adhered to the leaf blade was carefully counted using a stereoscopic electric microscope (LEICA DMRE). By leaf disc, the number of spores in three random microscopic fields were counted. By plant, nine microscopic fields or observations were taken into account per week. The average number of spores adhered to each leaf was estimated in conidia/ mm2. All the spores observed under the microscope were recorded.
2.11. Processing of Data
2.12. Data Analyses
3. Results
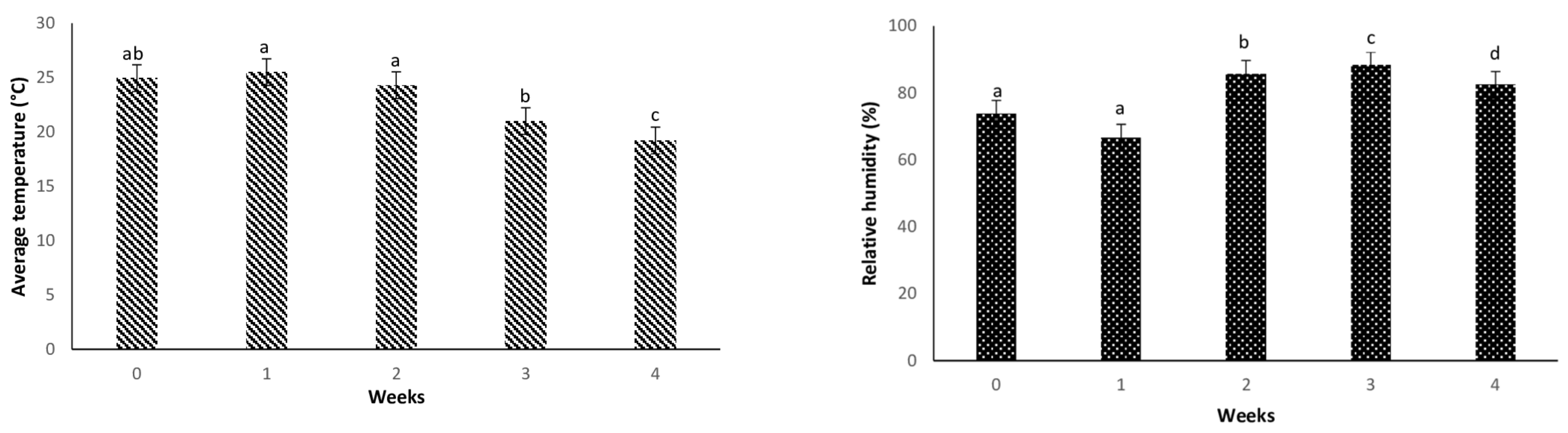
3.1. Weather Patterns
3.2. Weekly Ectophytic Presence of I. Javanica pf185 after Treatment
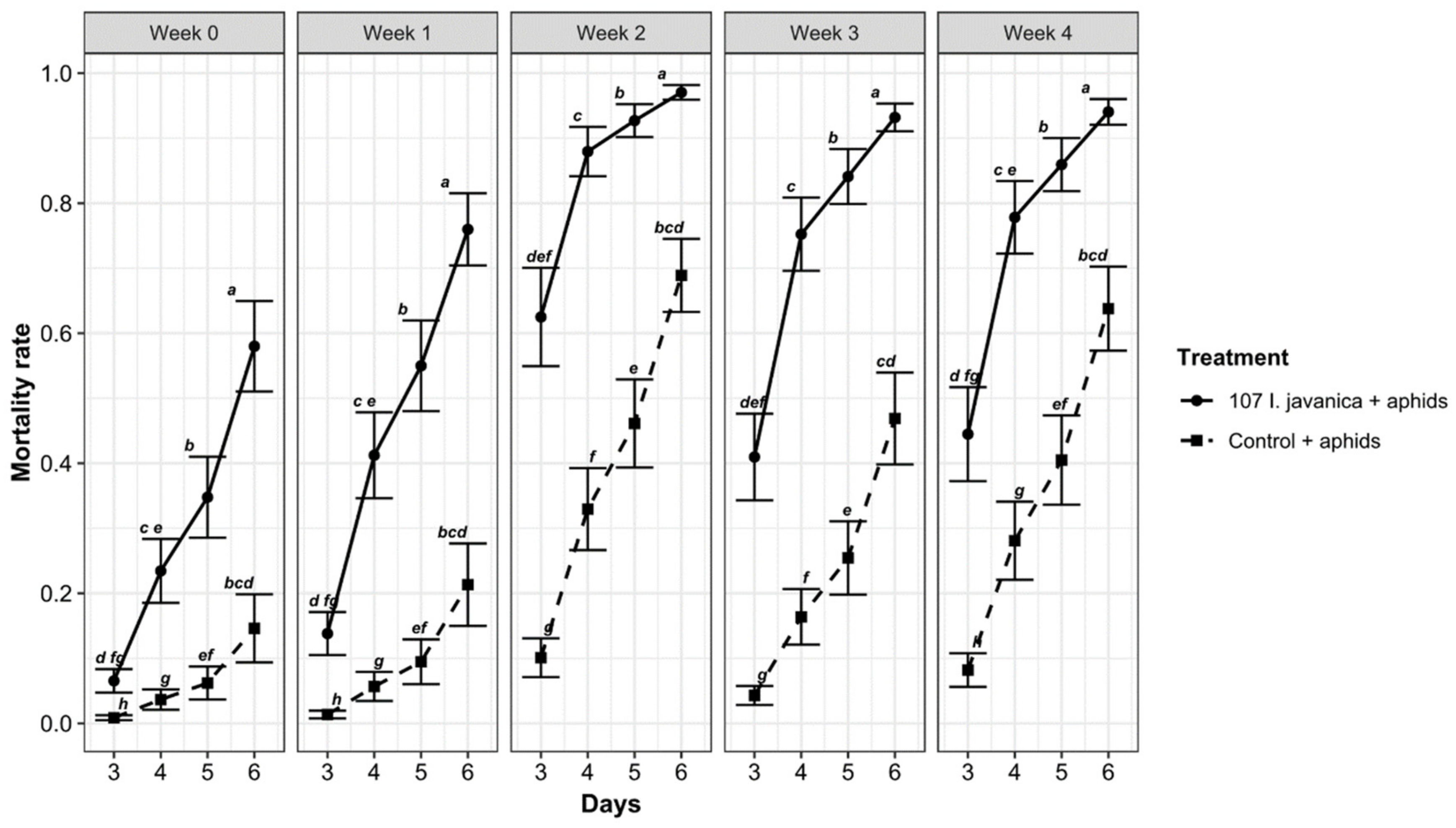
3.3. Daily Based Study of Insecticidal Activities on Fungus-Treated and Controlled Leaves per Week
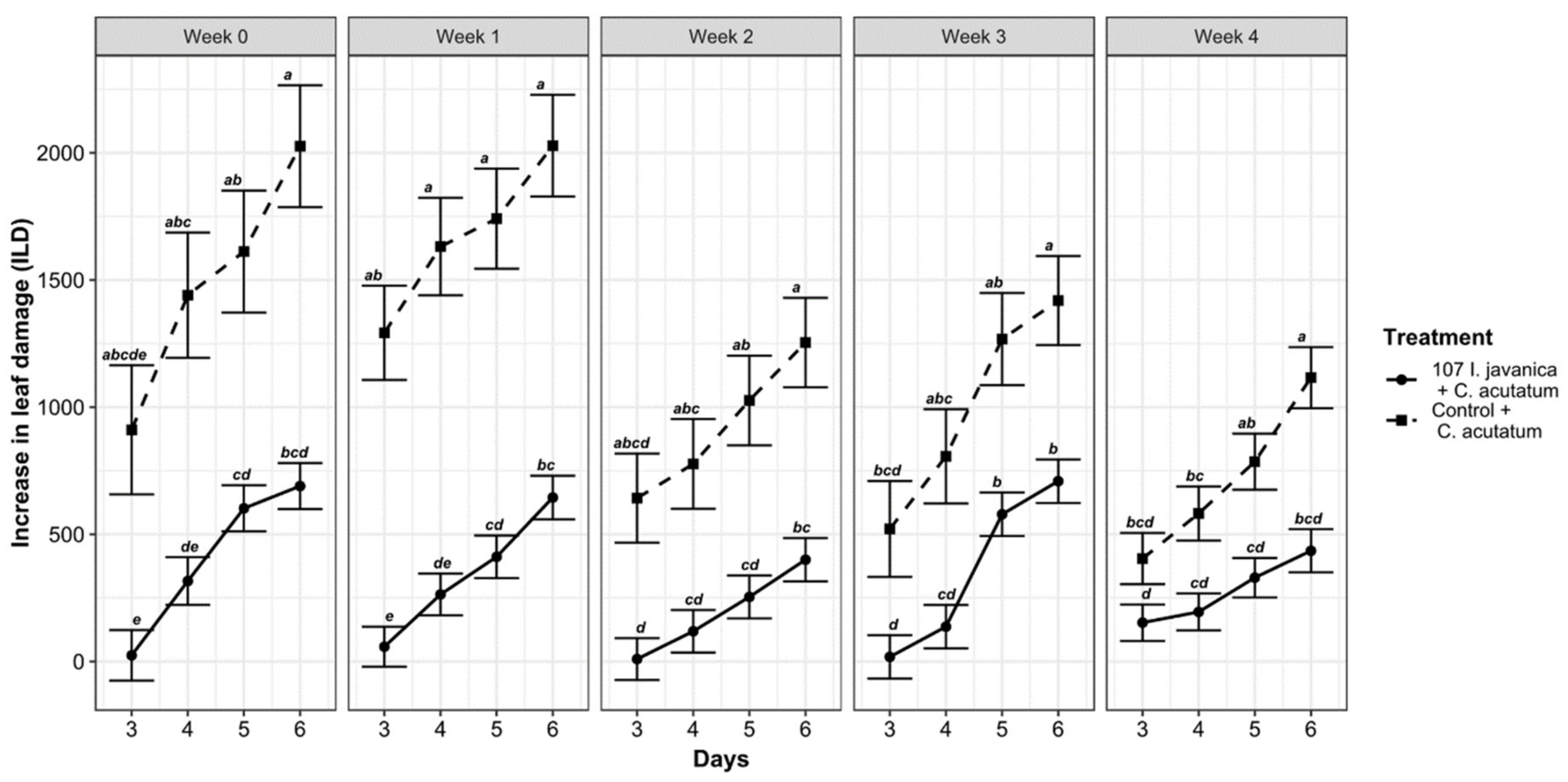
3.4. Daily Based Study of the Antifungal Activities on Fungus-Treated and Controlled Leaves per Week
3.5. Suspension Biocontrol Efficacy Per Week
3.6. Weekly Based Study of Correlations Between Biocontrol Efficacy and Weather Patterns
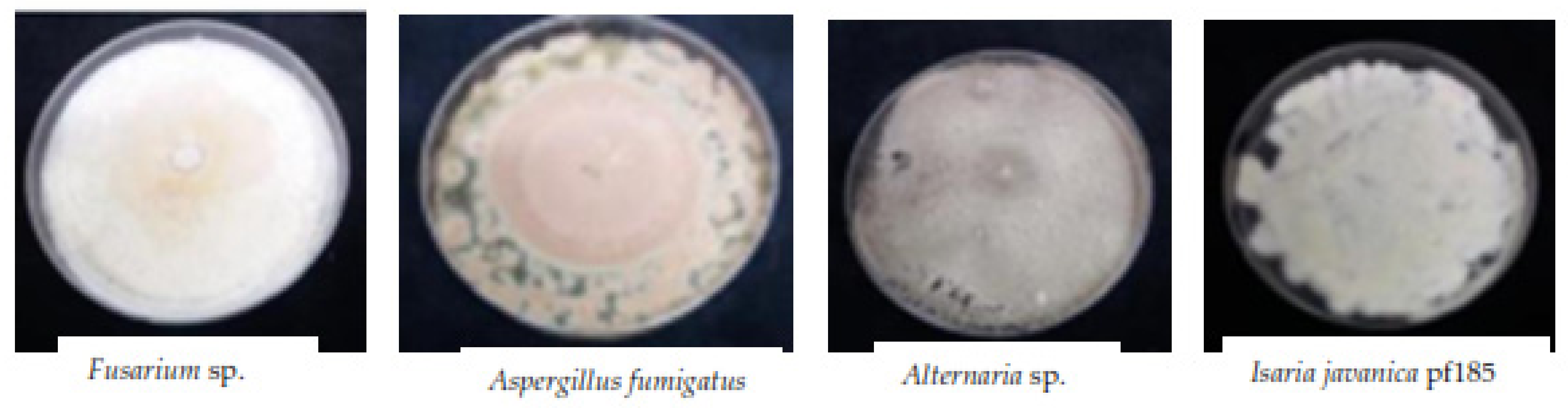
3.7. Endophytic Fungi Weekly Isolated from Leaf Tissues of Treated Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chunthawodtiporn, J.; Hill, T.; Stoffel, K.; Van Deynze, A. Quantitative Trait Loci controlling fruit size and other horticultural traits in Bell pepper (Capsicum annuum). Plant Genome 2018, 11, 160125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singletary, K.R. Pepper: Overview of potential health benefits. Nutr. Today 2011, 46, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Pawar, S.S.; Bharude, N.V.; Sonone, S.S.; Deshmukh, R.S.; Raut, A.K.; Umarkar, A.R. Chillies as food, spice and medicine: A perspective. Int. J. Pharm. Biol. Sci. 2011, 1, 311–318. [Google Scholar]
- Jagtap, P.P.; Shingane, U.S.; Kulkarni, K.P. Economics of chilli production in India. AJBAS 2012, 5, 161–164. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops—An Identification and Information Guide; The Natural History Museum; John Wiley and Sons, Ltd.: New York, NY, USA, 2000. [Google Scholar]
- Cahon, T.; Caillon, R.; Pincebourde, S. Do aphids alter leaf Surface temperature patterns during early infestation? Insects 2018, 9, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khamraev, A.S.; Davenport, C.F. Identification and Control of Agricultural Plant Pests and Diseases in Khorezm and Republic of Karakalpakstan, Uzbekistan (8) ZEF Work Papers for Sustainable Development in Central Asia; ZEF Bonn: Bonn, Germany, 2004; p. 132. [Google Scholar]
- Kmiec, K.; Rubinowska, K.; Michałek, W.; Sytykiewicz, H. The effect of galling aphids feeding on photosynthesis photochemistry of elm trees (Ulmus sp.). Photosynthetic 2018, 56, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Heimoana, S.C. The Effects of Aphids (Aphis gossypii) Glover on Photosynthesis in Cotton (Gossypium Hirsutum). Ph.D. Thesis, School of Agriculture and Food Sciences, The University of Queensland, Queensland, Australia, 2012; 259p. [Google Scholar]
- Mibey, R.K. 1.2.2.2 Sooty Moulds; Yair, B.-D., Chris, J.H., Eds.; World Crop Pests; Elsevier: Amsterdam, The Netherlands, 1997; pp. 275–290. [Google Scholar]
- Hughes, S.J. Sooty moulds. Mycologia 1976, 68, 693–820. [Google Scholar] [CrossRef]
- Muhammad, R.; Arshad, M.; Muhammad, A.; Muhammad, I.; Muhammad, A.; Safraz, A.S. Losses in yield and yield components caused by aphids to late sown Brassica napus L., Brassica juncea L., Brassica carrinata A. Braun at Multan, Punjab (Pakistan). Pak. J. Bot. 2011, 1, 319–324. [Google Scholar]
- Singh, N.; Dhiman, S. Quality and quantity loss by aphid infestation in vegetables grown under protected cultivation in Ladakh region. Def. Life Sci. J. 2018, 1, 71–74. [Google Scholar] [CrossRef]
- Reetsang, L.M.; Motshwari, O.; Aobakwe, B.N.T. Economic injury level and yield loss assessment caused by cabbage aphid (Brevicoryne brassicae) on rape (Brassica napus). J. Entomol. Res. 2015, 39, 101–104. [Google Scholar]
- Bailey, J.A.; Jeger, M.J. Colletotrichum: Biology, Pathology and Control; Commonwealth Mycological Institute: Wallingford, UK, 1992; p. 388. [Google Scholar]
- Cannon, P.F.; Bridge, P.D.; Monte, E. Linking the Past, Present, and Future of Colletotrichum Systematics. In Colletotrichum: Host Specificity, Pathology, and Host-Pathogen Interaction; Prusky, D., Freeman, S., Dickman, M., Eds.; APS Press: St. Paul, MN, USA, 2000; pp. 1–20. [Google Scholar]
- Lewis Ivey, M.L.; Nava-Diaz, C.; Miller Sally, A. Identification and management of Colletotrichum acutatum on immature bell peppers. Plant Dis. 2004, 88, 1198–1204. [Google Scholar] [CrossRef] [Green Version]
- Manandhar, J.B.; Hartman, G.L.; Wang, T.C. Anthracnose development on pepper fruits inoculated with Colletotrichum gloeosporioides. Plant Dis. 1995, 79, 380–383. [Google Scholar] [CrossRef]
- Sharma, P.N.; Kaur, M.; Sharma, O.P.; Sharma, P.; Pathania, A. Morphological, pathological and molecular variability in Colletotrichum capsici, the cause of fruit rot of chillies in the subtropical region of north-western India. J. Phytopathol. 2005, 4, 232–237. [Google Scholar] [CrossRef]
- Pakdeevaraporn, P.; Wasee, S.; Taylor, P.W.J.; Mongkolporn, O. Inheritance of resistance to anthracnose caused by Colletotrichum capsici in Capsicum. Plant Breed. 2005, 2, 206–208. [Google Scholar] [CrossRef]
- Than, P.P.; Jeewon, R.; Hyde, K.D.; Pongsupasamit, S.; Mongkolporn, O.; Taylor, P.W.J. Characterization and pathogenicity of Colletotrichum species associated with anthracnose disease on chilli (Capsicum spp.) in Thailand. Plant Pathol. 2008, 3, 562–572. [Google Scholar] [CrossRef]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Jang, M.R.; Moon, H.K.; Kim, T.R.; Yuk, D.H.; Kim, J.H.; Park, S.G. Dietary risk assessment for pesticide residues of vegetables in Seoul, Korea. Korean J. Nutr. 2010, 43, 404–412. [Google Scholar] [CrossRef]
- Lakhani, L.; Soni, D.; Alune, B. Dangers of pesticides on wildlife ecology. IJRG 2015, 9, 1–3. [Google Scholar]
- Witczak, A.; Abdel-Gawad, H. Assessment of health risk from organochlorine pesticides residues in high-fat spreadable foods produced in Poland. J. Environ. Sci. Health Part B 2014, 49, 917–928. [Google Scholar] [CrossRef]
- Chourasiya, S.; Khillare, P.S.; Jyethi, D.S. Health risk assessment of organochlorine pesticide exposure through dietary intake of vegetables grown in the periurban sites of Delhi, India. Environ. Sci. Pollut. Res. Int. 2015, 22, 5793–5806. [Google Scholar] [CrossRef]
- Reiler, E.; Jørs, E.; Bælum, J.; Huici, O.; Alvarez Caero, M.M.; Cedergreen, N. The influence of tomato processing on residues of organochlorine and organophosphate insecticides and their associated dietary risk. Sci. Total Environ. 2015, 527, 262–269. [Google Scholar] [CrossRef]
- Peres, N.A.R.; de Souza, N.L.; Peever, T.L.; Timmer, L.W. Benomyl sensitivity of isolates of Colletotrichum acutatum and C. gloeosporioides from Citrus. Plant Dis. 2004, 88, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Jallow, M.F.; Awadh, D.G.; Albaho, M.S.; Devi, V.Y.; Thomas, B.M. Pesticide risk behaviors and factors influencing pesticide use among farmers in Kuwait. Sci. Total Environ. 2017, 574, 490–498. [Google Scholar] [CrossRef]
- Gemechu, K.; Endashaw, B.; Emana, G.; Muhammad, I.; Tebkew, D.; Bayeh, M.; Kifle, D. Breeding food legumes for Resistance to storage insect pests: Potential and limitations. Sustainability 2011, 3, 1399–1415. [Google Scholar]
- Lingappa, S.; Saxena, H.; Devi Vimala, P.S. Role of biocontrol agents in management of Helicoverpa armigera (Hubner). In Recent Advances in Helicoverpa armigera Management; Saxena, H., Rai, A., Ahmad, R., Gupta, S., Eds.; Indian Society of Pulses Research and Development, IIPR: Kanpur, India, 2005; pp. 159–184. [Google Scholar]
- Kang, B.R.; Han, J.H.; Kim, J.J.; Kim, Y.C. Dual Biocontrol potential of the entomopathogenic fungus, Isaria javanica, for both aphids and plant fungal pathogens. Mycobiology 2018, 4, 440–447. [Google Scholar] [CrossRef] [Green Version]
- Giné, A.; Carrasquilla, M.; Martínez-Alonso, M.; Gaju, N.; Sorribas, F.J. Characterization of soil suppressiveness to root-knot nematodes in organic horticulture in plastic greenhouse. Front. Plant Sci. 2016, 7, 164. [Google Scholar] [CrossRef] [Green Version]
- Han, J.-H.; Jin, B.R.; Kim, J.-J.; Lee, S.-Y. Virulence of entomopathogenic fungi Metarhizium anisopliae and Paecilomyces fumusorosea for the microbial control of Spodoptera exigua. Microbiology 2014, 4, 385–390. [Google Scholar]
- Bocco, R.; Gandonou, C.; Amoussou, P.-L.; Togola, A.; Dieng, I.; Ndjiondjop, M.N.; Seck, P.A.; Tamo, M. Rapid phenotyping for identification of rice resistant varieties to Diopsis apicalis (Diptera: Diopsidae) Dalman. Cogent Biol. 2019, 5, 1649851. [Google Scholar]
- Rural Development Administration (RDA). Cultivation Technology in Pepper (Agricultural Technique Guide); RDA: Jeonju, Korea, 2017; pp. 83–127.
- Petrini, O. Taxonomy of endophytic fungi of aerial plant tissues. In Microbiology of the Phyllosphere; Fokkema, N.J., van den Heuvel, J., Eds.; Cambridge University Press: Cambridge, UK, 1986; pp. 175–187. [Google Scholar]
- Giridharan, P.; Verekar, S.A.; Gohil, A.R.; Mishra, P.D.; Khanna, A.; Deshmukh, S.K. Antiproliferative Activity of Hamigerone and Radicinol Isolated from Bipolaris papendorfii. Biomed. Res. Int. 2014, 2014, 890904. [Google Scholar] [CrossRef]
- Booth, C. Fusarium. Laboratory Guide to the identification of the Major Species; Commonwealth Mycological Institute: Kew, UK, 1977; 58p. [Google Scholar]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; Macmillan Publishing Co.: New York, NY, USA, 1986; 218p. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. Fusarium Laboratory Manual; SB741.F9L47; Blackwell Publishing: Hoboken, NJ, USA, 2006; 388p, Available online: www.blackwellprofessional.com (accessed on 17 May 2018).
- Püntener, W. Manual for Field Trials in Plant Protection, 2nd ed.; Agricultural Division, Ciba-Geigy Limited: Basle, Switzerland, 1981. [Google Scholar]
- Henderson, C.F.; Tilton, E.W. Test with acaricides against the brown wheat mites. J. Econ. Entomol. 1955, 48, 157–161. [Google Scholar] [CrossRef]
- VSN International GenStat for Windows, 15th ed.; VSN International: Hemel Hempstead, UK, 2012; Available online: GenStat.co.uk (accessed on 22 May 2013).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-137. 2018. Available online: https://CRAN.R-project.org/package=nlme (accessed on 11 September 2019).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Brooks, E.M.; Kristensen, K.; van Benthem, J.K.; Magnusson, A.; Berg, W.C.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, M.B. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 2, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B.; R Development Core Team. bbmle: Tools for General Maximum Likelihood Estimation. R Package Version 1.0.20. 2017. Available online: https://CRAN.R-project.org/package=bbmle (accessed on 11 September 2019).
- Lenth, R.; emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.3.0. 2019. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 11 September 2019).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 11 September 2019).
- Karlsson, I.; Friberg, H.; Steinberg, C.; Persson, P. Fungicide effects on fungal community composition in the wheat phyllosphere. PLoS ONE 2014, 9, e111786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Last, F.T. Seasonal incidence of Sporobolomyces on cereal leaves. Trans. Br. Mycol. Soc. 1955, 38, 221–239. [Google Scholar] [CrossRef]
- Blixt, E.; Olson, Å.; Lindahl, B.; Djurle, A.; Yuen, J. Spatiotemporal variation in the fungal community associated with wheat leaves showing symptoms similar to Stagonospora nodorum blotch. Eur. J. Plant Pathol. 2010, 126, 373–386. [Google Scholar] [CrossRef]
- Southwell, R.J.; Brown, J.F.; Welsby, S.M. Microbial interactions on the phylloplane of wheat and barley after applications of mancozeb and triadimefon. Australas. Plant Pathol. 1999, 28, 139–148. [Google Scholar] [CrossRef]
- Boomsma, J.; Jensen, A.; Meyling, N.; Eilenberg, J. Evolutionary interaction networks of insect pathogenic fungi. Environ. Entomol. 2014, 59, 467–485. [Google Scholar] [CrossRef]
- Gouli, V.; Gouli, S.; Marcelino, A.P.; Skinner, M.; Parker, B.L. Entomopathogenic fungi associated with invasive insect pests in Northeastern forests of the USA. Insects 2014, 4, 631–645. [Google Scholar] [CrossRef]
- Luo, S.; He, M.; Cao, Y.; Yuxian, X. The tetraspanin gene MaPls1 contributes to virulence by affecting germination, appressorial function and enzymes for cuticle degradation in the entomopathogenic fungus, Metarhizium acridum. Environ. Microbiol. 2014, 11, 2966–2979. [Google Scholar]
- Mantzoukas, S.; Lagogiannis, I. Endophytic colonization of pepper (Capsicum annum) controls aphids (Myzus persicae Sulzer). Appl. Sci. 2019, 9, 2239. [Google Scholar] [CrossRef] [Green Version]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Stinson, M.; Ezra, D.; Hess, W.M.; Sears, J.; Strobel, G. An endophytic Gliocladium sp. of Eucryphia cordifolia producing selective volatile antimicrobial compounds. Plant Sci. 2003, 165, 913–922. [Google Scholar] [CrossRef]
- Lee, S.O.; Kim, H.Y.; Choi, G.J.; Lee, H.B.; Jang, K.S.; Choi, Y.H.; Kim, J.C. Mycofumigation with Oxyporus latemarginatus EF069 for control of postharvest apple decay and Rhizoctonia root rot on moth orchid. J. Appl. Microbiol. 2009, 106, 1213–1219. [Google Scholar] [CrossRef]
- Sherameti, I.; Shahollari, B.; Venus, Y.; Altschmied, L.; Varma, A.; Oelmuller, R. The endophytic fungus Piriformospora indica stimulates the expression of nitrate reductase and the starch-degrading enzyme glucan-water dikinase in tobacco and Arabidopsis roots through a homeodomain transcription factor that binds to a conserved motif in their promoters. J. Biol. Chem. 2005, 280, 26241–26247. [Google Scholar]
- Bailey, B.A.; Bae, H.; Strem, M.D.; Robert, D.P.; Thomas, S.E.; Crozier, J.; Samuels, G.J.; Choi, I.Y.; Holmes, K.A. Fungal and plant gene expression during the colonization of cacao seedlings by endophytic isolates of four Trichoderma species. Planta 2006, 224, 1449–1464. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Q.; Gao, Y.Q.; Tang, J.J.; Zhang, A.L.; Gao, J.M. Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J. Agric. Food Chem. 2014, 16, 3584–3590. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Han, J.-H.; Kang, B.R.; Kim, Y.C. Dubityl succinate, produced by an insect-pathogenic fungus, Isaria javanica pf185, is a metabolite that controls of aphids and fungal disease, anthracnose. Pest Manag. Sci. 2019, 75, 852–858. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I. Fungal endophytes and their interaction with plant pathogens. Span. J Agric. Res. 2008, 6, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Pessarakli, A.H. A review on biological control of fungal plant pathogens using microbial antagonists. J. Biol. Sci. 2010, 4, 273–290. [Google Scholar]
- Berthelot, C.; Leyval, C.; Foulon, j.; Chalot, M.; Blaudez, D. Plant growth promotion, metabolite production and metal tolerance of dark septate endophytes isolated from metal-polluted poplar phytomanagement sites. FEMS Ecol. 2016, 2, fiw144. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Hao, C.; Liang, L.; Ma, R.-Y. Effects of temperature and relative humidity on conidial germination of Isaria fumosorosea (Hypocreales: Cordycipitaceae) IF-1106 and pathogenicity of the fungus against Bemisia tabaci (Homoptera: Aleyrodidae). Mycosystema 2014, 3, 668–679. [Google Scholar]
- Luz, C.; Rocha, L.F.N.; Nery, G.V. Detection of entomopathogenic fungi in peridomestic triatomine-infested areas in central Brazil and fungal activity against Triatoma infestans (Klug) (Hemiptera: Reduviidae). Neotrop. Entomol. 2004, 6, 783–791. [Google Scholar] [CrossRef]
- Sapna, M.; Peeyush, K.; Anushree, M. Effect of temperature and humidity on pathogenicity of native Beauveria bassiana isolate against Musca domestica L. J. Parasit. Dis. 2015, 4, 697–704. [Google Scholar]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The status of biological control and recommendations for improving uptake for the future. BioControl 2017, 1, 155–167. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeks | %Ecto I. javanica | CFU/mm2 | Adhered Spores/mm2 | Density w_w/mm2 | |||
|---|---|---|---|---|---|---|---|
| Control | Treated | Control | Treated | Control | Treated | ||
| 0 | 23.26 | 0.5 ± 0.2 b | 2.7 ± 0.6 a | 101.4 ± 12.7 b | 212.5 ± 9.3 a | 192.9 ± 36.1 b | 964.5 ± 28.5 a |
| 1 | 46.51 | 0.2 ± 0.1 a | 1.1 ± 0.5 a | 99.0 ± 6.0 a | 117.7 ± 10.3 a | 231.5 ± 28.5 b | 501.5 ± 38.2 a |
| 2 | 20.93 | 4.9 ± 2.0 b | 17.1 ± 6.2 a | 38.8 ± 5.9 a | 54.5 ± 6.9 a | 424.4 ± 32.5 b | 733.0 ± 36.1 a |
| 3 | 4.65 | 9.3 ±4.0 a | 10.5 ± 2.0 a | 137.5 ± 13.7 b | 180.7 ± 12.1 a | 347.2 ± 27.0 a | 578.7 ± 46.8 a |
| 4 | 4.65 | 5.6 ± 1.5 b | 16.6 ± 4.2 a | 98.0 ± 8.3 a | 117.2 ± 11.1 a | 463.0 ± 50.2 a | 733.0 ± 46.8 a |
| Weeks | Treatment | Mean (%) | SE | CV | Min | Max |
|---|---|---|---|---|---|---|
| 0. | I. javanica pf185 + Aphid | 25.56 | 4.08 | 135.37 | 0.00 | 100.00 |
| 0 | Control + Aphid | 8.61 | 1.47 | 144.95 | 0.00 | 40.00 |
| 1 | I. javanica pf185 + Aphid | 42.78 | 4.48 | 88.88 | 0.00 | 100.00 |
| 1 | Control + Aphid | 11.94 | 2.23 | 158.13 | 0.00 | 80.00 |
| 2 | I. javanica pf185 + Aphid | 83.33 | 3.04 | 30.94 | 0.00 | 100.00 |
| 2 | Control + Aphid | 40.56 | 2.56 | 53.62 | 0.00 | 100.00 |
| 3 | I. javanica pf185 + Aphid | 73.33 | 3.47 | 40.17 | 0.00 | 100.00 |
| 3 | Control + Aphid | 22.22 | 2.33 | 88.7 | 0.00 | 80.00 |
| 4 | I. javanica pf185 + Aphid | 80.28 | 3.27 | 34.61 | 0.00 | 100.00 |
| 4 | Control + Aphid | 28.89 | 2.85 | 83.70 | 0.00 | 80.00 |
| Weeks | Treatment | Mean (%) | SE | CV | Min | Max |
|---|---|---|---|---|---|---|
| 0 | I. javanica pf185 + C. acutatum | 525 | 52.991 | 85.647 | 0 | 1600 |
| 0 | Control + C. acutatum | 1173.91 | 121.998 | 86.326 | 200 | 4800 |
| 1 | I. javanica pf185 + C. acutatum | 419.44 | 42.636 | 86.252 | 0 | 1200 |
| 1 | Control + C. acutatum | 1500 | 88.103 | 49.839 | 200 | 3800 |
| 2 | I. javanica pf185 + C. acutatum | 225 | 36.673 | 138.301 | 0 | 1200 |
| 2 | Control + C. acutatum | 863.89 | 74.622 | 73.295 | 0 | 3000 |
| 3 | I. javanica pf185 + C. acutatum | 369.44 | 48.878 | 112.261 | 0 | 2000 |
| 3 | Control + C. acutatum | 983.33 | 88.965 | 76.768 | 0 | 3400 |
| 4 | I. javanica pf185 + C. acutatum | 279.37 | 32.804 | 93.201 | 0 | 1000 |
| 4 | Control + C. acutatum | 733.33 | 56.085 | 64.895 | 0 | 2000 |
| Weeks | Insecticidal Efficacy (%) | Antifungal Efficacy (%) |
|---|---|---|
| 0 | 20.43 | 26.07 |
| 1 | 39.82 | 38.01 |
| 2 | 72.32 | 53.35 |
| 3 | 66.43 | 29.08 |
| 4 | 70.04 | 41.81 |
| Variables | Insecticidal | Antifungal | Humidity | Temperature |
|---|---|---|---|---|
| Insecticidal | 1 | |||
| Antifungal | 0.638 | 1 | ||
| Humidity | 0.788 | 0.248 | 1 | |
| Temperature | −0.651 | 0.002 | −0.648 | 1 |
| Weeks | Plants | Isolated | I. javanica pf185 | Verticillium sp. | Alternaria sp. | Unknown | Aspergillus fumigatus | Acremonium sp. | Fusarium sp. | Cladosporium sphaerospermum |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | TP1 | 73 | 73 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| TP2 | 53 | 34 | 2 | 2 | 15 | 0 | 0 | 0 | 0 | |
| TP3 | 32 | 1 | 0 | 0 | 0 | 31 | 0 | 0 | 0 | |
| 1 | TP1 | 54 | 44 | 0 | 6 | 1 | 0 | 3 | 0 | 0 |
| TP2 | 20 | 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| TP3 | 51 | 51 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 2 | TP1 | 30 | 0 | 0 | 0 | 0 | 30 | 0 | 0 | 0 |
| TP2 | 37 | 22 | 2 | 0 | 1 | 8 | 0 | 4 | 0 | |
| TP3 | 19 | 1 | 9 | 0 | 0 | 9 | 0 | 0 | 0 | |
| 3 | TP1 | 33 | 15 | 0 | 16 | 0 | 0 | 0 | 0 | 2 |
| TP2 | 35 | 35 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| TP3 | 36 | 35 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |
| 4 | TP1 | 10 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 |
| TP2 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | |
| TP3 | 14 | 2 | 0 | 0 | 0 | 0 | 12 | 0 | 0 | |
| Endo CF | 65.68 | 2.56 | 6.71 | 3.35 | 15.38 | 2.96 | 0.99 | 2.37 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocco, R.; Lee, M.; Kim, D.; Ahn, S.; Park, J.-W.; Lee, S.-Y.; Han, J.-H. Endophytic Isaria javanica pf185 Persists after Spraying and Controls Myzus persicae (Hemiptera: Aphididae) and Colletotrichum acutatum (Glomerellales: Glomerellaceae) in Pepper. Insects 2021, 12, 631. https://doi.org/10.3390/insects12070631
Bocco R, Lee M, Kim D, Ahn S, Park J-W, Lee S-Y, Han J-H. Endophytic Isaria javanica pf185 Persists after Spraying and Controls Myzus persicae (Hemiptera: Aphididae) and Colletotrichum acutatum (Glomerellales: Glomerellaceae) in Pepper. Insects. 2021; 12(7):631. https://doi.org/10.3390/insects12070631
Chicago/Turabian StyleBocco, Roland, Moran Lee, Dayeon Kim, Seongho Ahn, Jin-Woo Park, Sang-Yeob Lee, and Ji-Hee Han. 2021. "Endophytic Isaria javanica pf185 Persists after Spraying and Controls Myzus persicae (Hemiptera: Aphididae) and Colletotrichum acutatum (Glomerellales: Glomerellaceae) in Pepper" Insects 12, no. 7: 631. https://doi.org/10.3390/insects12070631
APA StyleBocco, R., Lee, M., Kim, D., Ahn, S., Park, J. -W., Lee, S. -Y., & Han, J. -H. (2021). Endophytic Isaria javanica pf185 Persists after Spraying and Controls Myzus persicae (Hemiptera: Aphididae) and Colletotrichum acutatum (Glomerellales: Glomerellaceae) in Pepper. Insects, 12(7), 631. https://doi.org/10.3390/insects12070631