
Mitochondrial DNA Variation in Peruvian Honey Bee (Apis mellifera L.) Populations Using the tRNAleu-cox2 Intergenic Region

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Sites
2.2. Mitochondrial DNA Analysis
2.3. Genetic Diversity and Phylogenetic Analysis
3. Results
3.1. New Haplotype and Variants
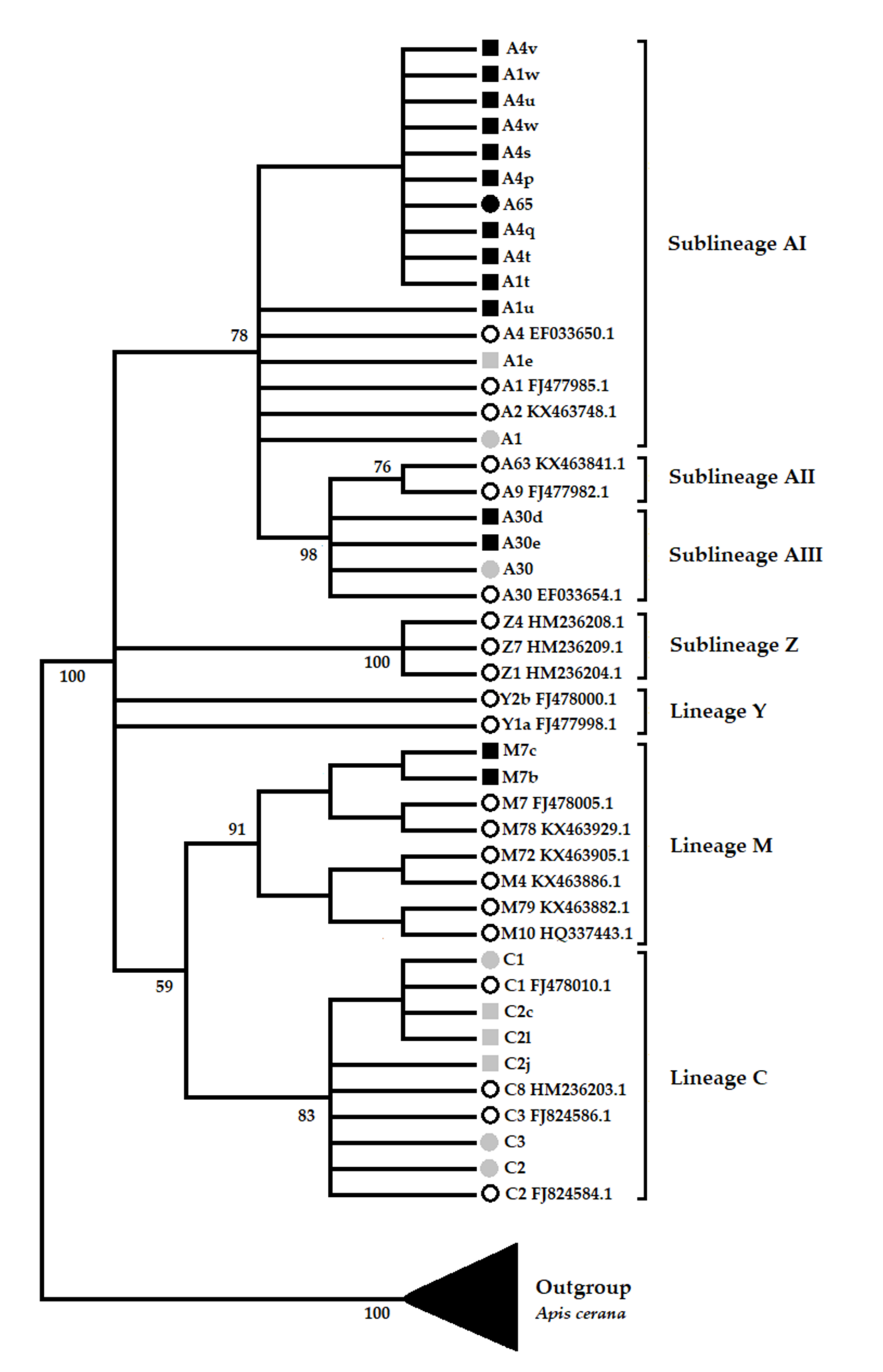
3.2. Phylogenetics Analysis of New Haplotypes and Variants
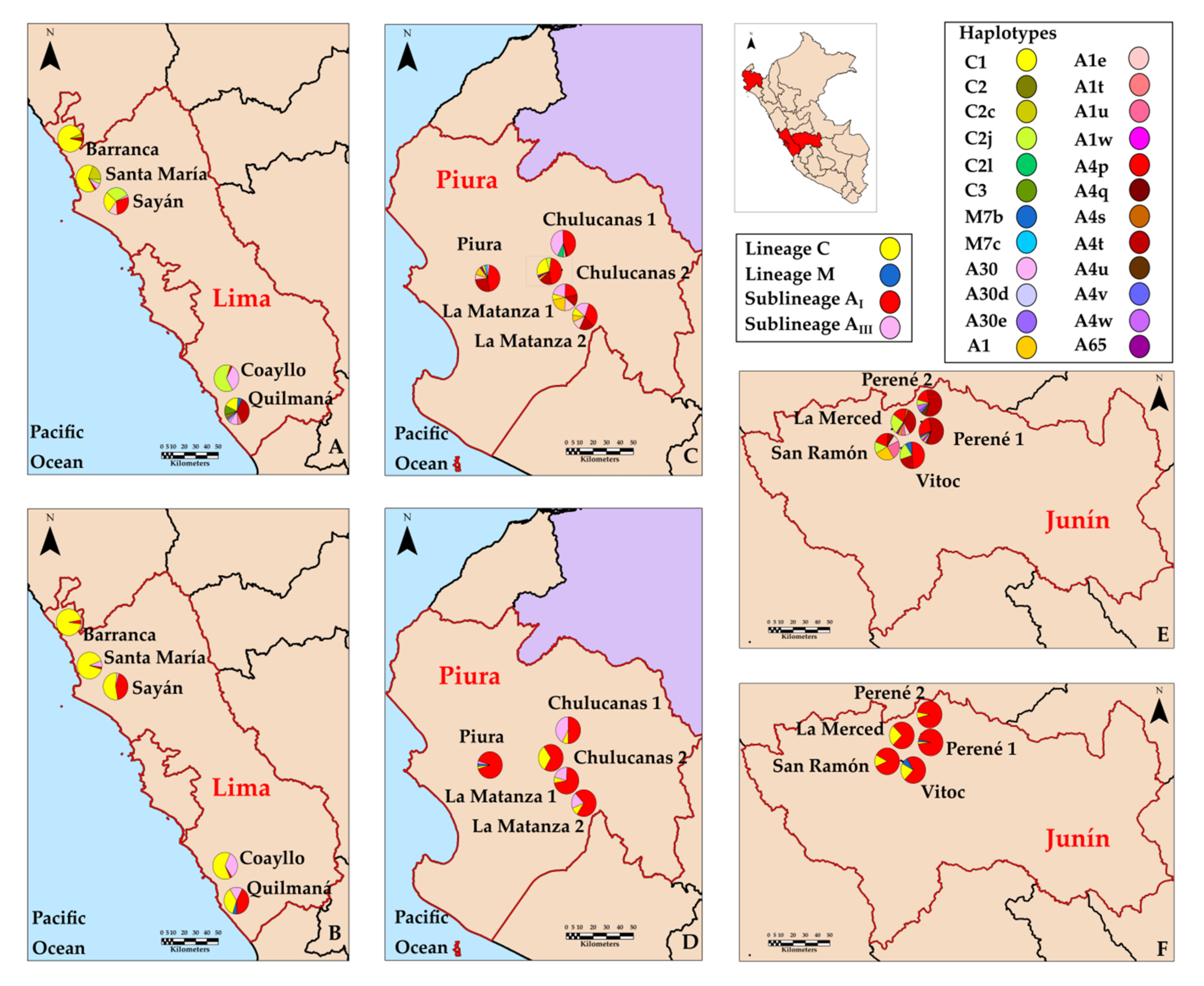
3.3. Distribution of Haplotypes
3.4. Diversity Measures
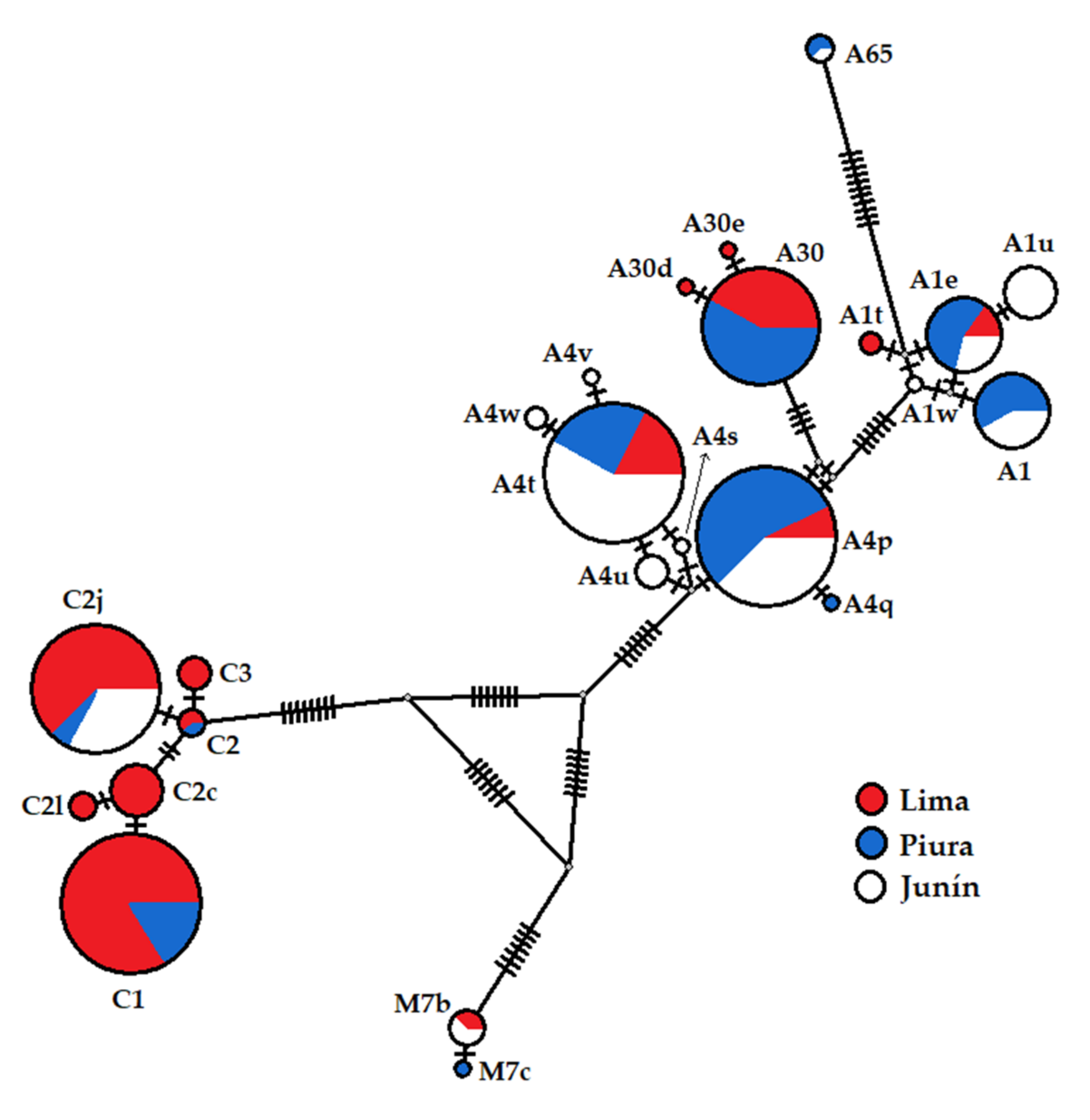
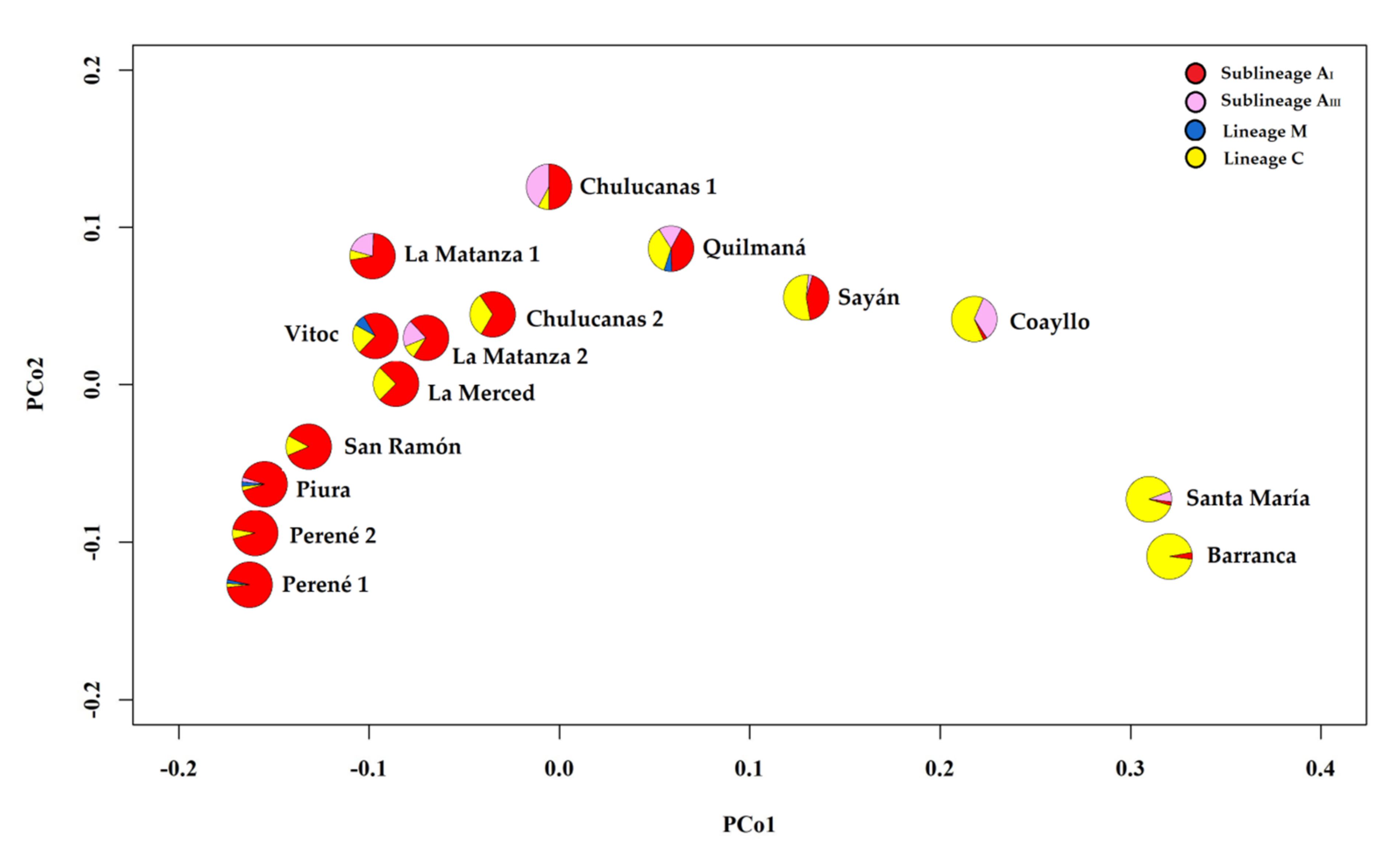
3.5. Relationships among Haplotypes and Lineages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hung, K.-L.J.; Kingston, J.M.; Albrecht, M.; Holway, D.A.; Kohn, J.R. The worldwide importance of honey bees as pollinators in natural habitats. Proc. R. Soc. B 2018, 285, 20172140. [Google Scholar] [CrossRef] [Green Version]
- Lautenbach, S.; Seppelt, R.; Liebscher, J.; Dormann, C.F. Spatial and temporal trends of global pollination benefit. PLoS ONE 2012, 7, e35954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristov, P.; Neov, B.; Shumkova, R.; Palova, N. Significance of Apoidea as main pollinators. Ecological and economic impact and implications for human nutrition. Diversity 2020, 12, 280. [Google Scholar] [CrossRef]
- Ruttner, F. Biogeography and Taxonomy of Honey Bees; Springer: Berlin, Germany, 1988. [Google Scholar]
- Chen, C.; Liu, Z.; Pan, Q.; Chen, X.; Wang, H.; Guo, H.; Liu, S.; Lu, H.; Tian, S.; Li, R.; et al. Genomic analyses reveal demographic history and temperate adaptation of the newly discovered honey bee subspecies Apis mellifera sinisxinyuan n. ssp. Mol. Biol. Evol. 2016, 33, 1337–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilyasov, R.A.; Lee, M.-I.; Takahashi, J.-I.; Kwon, H.W.; Nikolenko, A.G. A revision of subspecies structure of Western honey bee Apis mellifera. Saudi J. Biol. Sci. 2020, 27, 3615–3621. [Google Scholar] [CrossRef]
- Franck, P.; Garnery, L.; Loiseau, A.; Oldroyd, B.P.; Hepburn, H.R.; Solignac, M.; Cornuet, J.M. Genetic diversity of the honey bee in Africa: Microsatellite and mitochondrial data. Heredity 2001, 86, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Tihelka, E.; Cai, C.; Pisani, D.; Donoghue, P.C.J. Mitochondrial genomes illuminate the evolutionary history of the Western honey bee (Apis mellifera). Sci. Rep. 2020, 10, 13515. [Google Scholar] [CrossRef]
- Alburaki, M.; Moulin, S.; Legout, H.; Alburaki, A.; Garnery, L. Mitochondrial structure of Eastern honeybee populations from Syria, Lebanon and Iraq. Apidologie 2011, 42, 628–641. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.C.; Rinderer, T.E.; Sheppard, W.S. Further characterization of honey bees from the Iberian Peninsula by allozyme, morphometric and mtDNA haplotype analyses. J. Apic. Res. 2006, 45, 188–196. [Google Scholar] [CrossRef]
- Henriques, D.; Chávez-Galarza, J.; Teixeira, J.S.G.; Ferreira, H.; Neves, C.J.; Francoy, T.M.; Pinto, M.A. Wing Geometric morphometrics of workers and drones and single nucleotide polymorphisms provide similar genetic structure in the Iberian honey bee (Apis mellifera iberiensis). Insects 2020, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.; Page, R.E.; Crosland, M.W. Clinal variation and selection of MDH allozymes in honey bee populations. Experientia 1994, 50, 867–871. [Google Scholar] [CrossRef]
- Smith, D.R.; Glenn, T.C. Allozyme polymorphisms in Spanish honeybees (Apis mellifera iberica). J. Hered. 1995, 86, 12–16. [Google Scholar] [CrossRef]
- Del Lama, M.A.; Souza, R.O.; Durán, X.A.A.; Soares, A.E.E. Clinal variation and selection on MDH allozymes in honeybees in Chile. Hereditas 2004, 140, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.R.; Palopoli, M.F.; Taylor, B.R.; Garnery, L.; Cornuet, J.-M.; Solignac, M.; Brown, W.M. Geographical overlap of two mitochondrial genomes in Spanish honeybees (Apis mellifera iberica). J. Hered. 1991, 82, 96–100. [Google Scholar] [CrossRef]
- Franck, P.; Garnery, L.; Solignac, M.; Cornuet, J.M. The origin of west European subspecies of honeybees (Apis mellifera): New insights from microsatellite and mitochondrial data. Evolution 1998, 52, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Galarza, J.; Garnery, L.; Henriques, D.; Neves, C.J.; Loucif-Ayad, W.; Jonhston, J.S.; Pinto, M.A. Mitochondrial DNA variation of Apis mellifera iberiensis: Further insights from a large-scale study using sequence data of the tRNA leu-cox2 intergenic region. Apidologie 2017, 48, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Techer, M.A.; Clémencet, J.; Simiand, C.; Preeaduth, S.; Azali, H.A.; Reynaud, B.; Hélène, D. Large-scale mitochondrial DNA analysis of native honey bee Apis mellifera populations reveals a new African subgroup private to the South West Indian Ocean islands. BMC Genet. 2017, 18, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cánovas, F.; De la Rúa, P.; Serrano, J.; Galián, J. Microsatellite variability reveals beekeeping influences on Iberian honeybee populations. Apidologie 2011, 42, 235–251. [Google Scholar] [CrossRef] [Green Version]
- Alburaki, M.; Bertrand, B.; Legout, H.; Moulin, S.; Alburaki, A.; Sheppard, W.S.; Garnery, L. A fifth major genetic group among honeybees revealed in Syria. BMC Genet. 2013, 14, 117. [Google Scholar] [CrossRef] [Green Version]
- Wallberg, A.; Han, F.; Wellhagen, G.; Dahle, B.; Kawata, M.; Haddad, N.; Paulino, Z.L.; Allsopp, M.H.; Kandemir, I.; De La Rúa, P.; et al. A worldwide survey of genome sequence variation provides insight into the evolutionary history of the honeybee Apis mellifera. Nat. Genet. 2014, 46, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Galarza, J.; Henriques, D.; Johnston, J.S.; Acevedo, J.C.; Patton, J.C.; Muñoz, I.; De La Rúa, P.; Pinto, M.A. Signatures of selection in the Iberian honey bee (Apis mellifera iberiensis) revealed by a genome scan analysis of single nucleotide polymorphisms. Mol. Ecol. 2013, 22, 5890–5907. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Galarza, J.; Henriques, D.; Johnston, J.S.; Carneiro, M.; Rufino, J.; Patton, J.C.; Pinto, M.A. Revisiting the Iberian honey bee (Apis mellifera iberiensis) contact zone: Maternal and genome-wide nuclear variations provide support for secondary contact from historical refugia. Mol. Ecol. 2015, 24, 2973–2992. [Google Scholar] [CrossRef] [PubMed]
- Meixner, M.D.; Pinto, M.A.; Bouga, M.; Kryger, P.; Ivanova, E.; Fuchs, S. Standard methods for characterizing subspecies and ecotypes of Apis mellifera. J. Apic. Res. 2013, 52, 1–27. [Google Scholar] [CrossRef]
- Muñoz, I.; Pinto, M.A.; De la Rúa, P. Effects of queen importation on the genetic diversity of Macaronesian island honey bee populations (Apis mellifera linneaus 1758). J. Apic. Res. 2014, 53, 296–302. [Google Scholar] [CrossRef]
- Bertrand, B.; Alburaki, M.; Legout, H.; Moulin, S.; Mougel, F.; Garnery, L. MtDNA COI-COII marker and drone congregation area: An efficient method to establish and monitor honeybee (Apis mellifera L.) conservation centres. Mol. Ecol. Resour. 2015, 15, 673–683. [Google Scholar] [CrossRef]
- De Jaime, J.M. Sobre la primicia hispana en cuanto a los envíos de abejas europeas a América. ILUIL 2003, 26, 595–612. [Google Scholar]
- Padilla, F.; Puerta, F.; Flores, J.M.; Bustos, M. Bees, apiculture and the new world. Arch. Zootec. 1992, 41, 563–567. [Google Scholar]
- Taylor, O.R. The past and possible future spread of Africanized honey bees in the Americas. Bee World 1977, 31, 15–24. [Google Scholar] [CrossRef]
- Pérez-Castro, E.; May-Itzá, W.; Quezada-Euán, J. Thirty years after: A survey on the distribution and expansion of Africanized honey bees (Apis mellifera) in Peru. J. Apic. Res. 2002, 41, 69–73. [Google Scholar] [CrossRef]
- Quezada-Euán, J.; Pérez-Castro, E.; May-Itzá, W. Hybridization between European and African-derived honeybee populations (Apis mellifera) at different altitudes in Perú. Apidologie 2003, 34, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Garnery, L.; Cornuet, J.M.; Solignac, M. Evolutionary history of the honey bee Apis mellifera inferred from mitochondrial DNA analysis. Mol. Ecol. 1992, 1, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Cánovas, F.; De la Rúa, P.; Serrano, J.; Galián, J. Geographical patterns of mitochondrial DNA variation in Apis mellifera iberiensis (Hymenoptera: Apidae). J. Zoolog. Syst. Evol. Res. 2008, 46, 24–30. [Google Scholar] [CrossRef]
- Pinto, M.A.; Muñoz, I.; Chávez-Galarza, J.; De la Rúa, P. The Atlantic side of the Iberian Peninsula: A hot-spot of novel African honey bee maternal diversity. Apidologie 2012, 43, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Rortais, A.; Arnold, G.; Alburaki, M.; Legout, H.; Garnery, L. Review of the DraI COI-COII test for the conservation of the black honeybee (Apis mellifera mellifera). Conser. Genet. Resour. 2011, 3, 383–391. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Garnery, L.; Solignac, M.; Celebrano, G.; Cornuet, J.M. A simple test using restricted PCR-amplified mitochondrial DNA to study the genetic structure of Apis mellifera L. Experientia 1993, 49, 1016–1021. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Phylogeny Inference Package (PHYLIP); Version 3.65c; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Collet, T.; Ferreira, K.M.; Arias, M.C.; Soares, A.E.E.; Del Lama, M.A. Genetic structure of Africanized honeybee populations (Apis mellifera L.) from Brazil and Uruguay viewed through mitochondrial DNA COI–COII patterns. Heredity 2006, 97, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, I.; Dall’Olio, R.; Lodesani, M.; De la Rúa, P. Population genetic structure of Coastal Croatian honeybees. Apidologie 2009, 40, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, I.; De la Rúa, P. Temporal analysis of genetic diversity in a honey bee mating area of an island population (La Palma, Canary Islands, Spain). J. Apic. Sci. 2013, 56, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.E.; Oldroyd, B.P.; Javier, J.; Quezada-Euán, G.; Rinderer, T.E. Origin of honeybees (Apis mellifera L.) from the Yucatan peninsula inferred from mitochondrial DNA analysis. Mol. Ecol. 2001, 10, 1347–1355. [Google Scholar] [CrossRef]
- Branchiccela, B.; Aguirre, C.; Parra, G.; Estay, P.; Zunino, P.; Antúnez, K. Genetic changes in Apis mellifera after 40 years of Africanization. Apidologie 2014, 45, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Segura, J.A.L. Highly polymorphic DNA markers in an Africanized honey bee population in Costa Rica. Gen. Mol. Biol. 2000, 23, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Abrahamovich, A.H.; Atela, O.; De la Rúa, P.; Galián, J. Assessment of the mitochondrial origin of honey bees from Argentina. J. Apic. Res. 2007, 46, 191–194. [Google Scholar] [CrossRef]
- Szalanski, L.; Magnus, R. Mitochondrial DNA characterization of Africanized honey bee (Apis mellifera L.) populations from the USA. J. Apic. Res. 2010, 49, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Tibatá, V.M.; Arias, E.; Corona, M.; Ariza Botero, F.; Figueroa-Ramírez, J.; Junca, H. Determination of the Africanized mitotypes in populations of honey bees (Apis mellifera L) of Colombia. J. Apic. Res. 2018, 57, 219–227. [Google Scholar] [CrossRef]
- Magnus, R.; Tripodi, A.D.; Szalanski, A.L. Mitochondrial DNA diversity of honey bees, Apis mellifera L. (Hymenoptera: Apidae) from queen breeders in the United States. J. Apicul. Sci. 2011, 55, 5–14. [Google Scholar]
- Vidal, M.C.; Garnery, L.; Legoux, H. Analysis of the Genetic Structure of bees in Chile: Identification of Africanized Bees using Mitochondrial DNA. In Proceedings of the 1st Symposium Apimondia on Organic Agriculture, Sunny Beach, Bulgaria, 27–29 August 2010. [Google Scholar]
- Moritz, R.F.A.; Härtel, S.; Neumann, P. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Écoscience 2005, 12, 289–301. [Google Scholar] [CrossRef]
- Kent, R.B. The African honeybee in Peru: An insect invader and its impact on beekeeping. Appl. Geogr. 1989, 9, 237–257. [Google Scholar] [CrossRef]
- Henriques, D.; Wallberg, A.; Chávez-Galarza, J.; Johnston, J.S.; Webster, M.T.; Pinto, M.A. Whole genome SNP-associated signatures of local adaptation in honeybees of the Iberian Peninsula. Sci. Rep. 2018, 8, 11145. [Google Scholar] [CrossRef] [Green Version]
- Franck, P.; Garnery, L.; Celebrano, G.; Solignac, M.; Cornuet, J.M. Hybrid origins of honey bees from Italy (Apis mellifera ligustica) and sicily (A. m. sicula). Mol. Ecol. 2000, 9, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Garnery, L.; Franck, P.; Baudry, E.; Vautrin, D.; Cornuet, J.-M.; Solignac, M. Genetic diversity of the west European honey bee (Apis mellifera mellifera and A. m. iberica). I. Mitochondrial DNA. Genet. Sel. Evol. 1998, 30, S31–S47. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Garnery, L.; Solignac, M. Putative origin and function of the intergenic region between COI and COII of Apis mellifera L. mitochondrial DNA. Genetics 1991, 128, 393–403. [Google Scholar] [CrossRef]
- Kono, Y.; Kohn, J.R. Range and frequency of Africanized honey bees in California (USA). PLoS ONE 2015, 10, e0137407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, R.M.; Wallberg, A.; Simões, Z.L.P.; Lawson, D.J.; Webster, M.T. Genome-wide analysis of admixture and adaptation in the Africanized honey bee. Mol. Ecol. 2017, 26, 3603–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, J.E.; Piganeau, G.; Eyre-Walker, A. The rate of adaptive evolution in animal mitochondria. Mol. Ecol. 2016, 25, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Sampling Sites | N | Na | Np | Ne | uh |
|---|---|---|---|---|---|---|
| Lima | Barranca | 42 | 3 | 1 | 1.215 | 0.181 |
| Coayllo | 38 | 3 | 1 | 1.936 | 0.496 | |
| Quilmaná | 36 | 9 | 1 | 4.909 | 0.819 | |
| Santa María | 39 | 5 | 1 | 2.164 | 0.552 | |
| Sayán | 35 | 5 | 1 | 3.840 | 0.761 | |
| Piura | Chulucanas 1 | 38 | 5 | 1 | 2.597 | 0.632 |
| Chulucanas 2 | 31 | 6 | 1 | 3.302 | 0.720 | |
| La Matanza 1 | 14 | 6 | 1 | 5.444 | 0.879 | |
| La Matanza 2 | 52 | 6 | 1 | 4.777 | 0.806 | |
| Piura | 31 | 8 | 1 | 3.469 | 0.735 | |
| Junín | La Merced | 32 | 8 | 1 | 4.414 | 0.798 |
| Perené 1 | 36 | 7 | 1 | 2.374 | 0.595 | |
| Perené 2 | 29 | 6 | 1 | 2.572 | 0.633 | |
| San Ramón | 35 | 7 | 1 | 5.258 | 0.834 | |
| Vitoc | 24 | 4 | 1 | 2.909 | 0.685 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chávez-Galarza, J.; López-Montañez, R.; Jiménez, A.; Ferro-Mauricio, R.; Oré, J.; Medina, S.; Rea, R.; Vásquez, H. Mitochondrial DNA Variation in Peruvian Honey Bee (Apis mellifera L.) Populations Using the tRNAleu-cox2 Intergenic Region. Insects 2021, 12, 641. https://doi.org/10.3390/insects12070641
Chávez-Galarza J, López-Montañez R, Jiménez A, Ferro-Mauricio R, Oré J, Medina S, Rea R, Vásquez H. Mitochondrial DNA Variation in Peruvian Honey Bee (Apis mellifera L.) Populations Using the tRNAleu-cox2 Intergenic Region. Insects. 2021; 12(7):641. https://doi.org/10.3390/insects12070641
Chicago/Turabian StyleChávez-Galarza, Julio, Ruth López-Montañez, Alejandra Jiménez, Rubén Ferro-Mauricio, Juan Oré, Sergio Medina, Reyna Rea, and Héctor Vásquez. 2021. "Mitochondrial DNA Variation in Peruvian Honey Bee (Apis mellifera L.) Populations Using the tRNAleu-cox2 Intergenic Region" Insects 12, no. 7: 641. https://doi.org/10.3390/insects12070641
APA StyleChávez-Galarza, J., López-Montañez, R., Jiménez, A., Ferro-Mauricio, R., Oré, J., Medina, S., Rea, R., & Vásquez, H. (2021). Mitochondrial DNA Variation in Peruvian Honey Bee (Apis mellifera L.) Populations Using the tRNAleu-cox2 Intergenic Region. Insects, 12(7), 641. https://doi.org/10.3390/insects12070641