3.1. Taxonomic Summary and Genus Diagnosis
Order Entomobryomorpha Börner, 1913 [
35]
Superfamily Entomobryoidea Womersley, 1934 [
21]
Family Orchesellidae Börner, 1906 [
10], sensu Zhang et al. 2019 [
3]
Subfamily Heteromurinae Absolon and Ksenemann, 1942 [
36], sensu Zhang and Deharveng, 2015 [
1]
Tribe Heteromurini Absolon and Ksenemann, 1942 [
36], sensu Zhang et al. 2020 [
8]
Genus
Falcomurus Mandal, 2018 [
15]
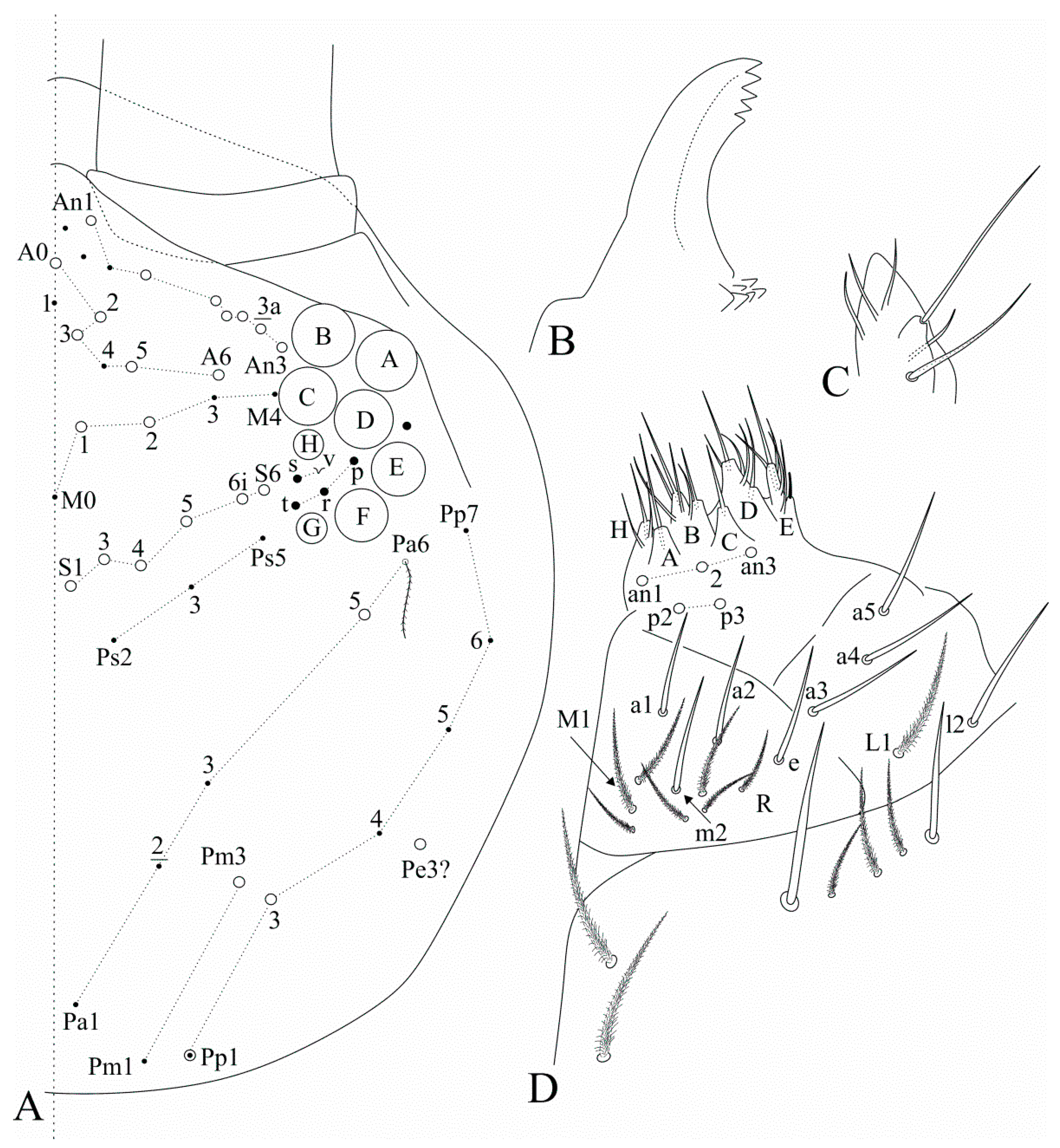
Diagnosis: Habitus similar to
Dicranocentrus (
Figure 1A). Apically rounded or truncate scales present at least over Ant. I, head, trunk and ventral furca. Ant. I subdivided in Ia and Ib, Ant. II subdivided or undivided, Ant. III and IV annulated. Dorsal head with at least
A0,
A2–
3,
A6,
M1–
2,
S1,
S3–
6 (including
S6i),
Pa5,
Pm3 and
Pp3 mac;
A1,
M3–
4,
S0,
S2 mac always absent. Prelabral chaetae normal (not bifurcate) and smooth, at least the labral
a1 chaeta enlarged. Eyes 8 + 8, postantennal organ absent. Tergal S-chaetotaxy from Th. II to Abd. V as 2,2|1,3,3,+,3 for sens, and 1,0|1,0,1,0,0 for ms. Trunk dorsal macrochaetotaxy reduced, Th. II with at least
a5,
m4–
4i,
p1–
3 (including
p2e) mac; Th. III main mac as
p1–
3 (including
p2e) and
a5; Abd. I devoid of mac; Abd. II mac as
m3–
3e,
m5; Abd. III main mac as
m3 and
pm6,
p6 mac possibly present in all species; Abd. IV with 2–3 central mac (
A3,
B5 and
A6, the latter also as mes). Ungues short and thick, internally with only the basal paired teeth, unguiculi excavated (
Figure 1B). Proximal dens with a typically falcate (sinuous) ciliate mac (
Figure 1C), dental spines absent, mucro bidentate, lacking the mucronal spine (
Figure 1D) (adapted from [
15,
17]).
Type species: Falcomurus chilikaensis Mandal, 2018.
Remarks on the genus:
Falcomurus was first described by [
15] as a monotypic genus from India. However, two other species described earlier,
Dicranocentrus litoreus, from the Philippines, and
D. halophilus from Papua New Guinea, mostly comply with Mandal’s description, with the exception of the absence of
Ps2 mac on the dorsal head and presence of an unguiculus outer tooth, and are herein transferred to
Falcomurus.
As originally noted by [
17],
D. litoreus and
D. halophilus show a different morphology compared to all other
Dicranocentrus species, with enlarged
a1–
2 chaetae on the distal labrum, a sinuous ciliate mac on the proximal dens, mucronal spine absent and a reduced dorsal head and trunk macrochaetotaxy. Mari-Mutt compared the dorsal chaetotaxy of his two species to those of
Heteromurus, in lacking the dorsal head
S0 chaeta, Th. III with five main mac (four in
Heteromurus) and Abd. I devoid of mac [
17,
37,
38]. This dorsal chaetotaxy, combined with antennae with five segments (only Ant. I subdivided) and mucro without spines, is more similar to the morphology of
Heteromurus s. str. than to
Dicranocentrus, although the annulated Ant. III is more similar to
Dicranocentrus [
8,
33].
After our morphological study of the three previously described and the two new species of
Falcomurus presented here, several shared features were noted between them and used to revise and complement the original diagnosis of the genus, especially regarding the main dorsal macrochaetotaxy. This character is mostly stable among the species, with few variations on the anterior and posterior head, Th. II
m4 complex and
p series, plus central Abd. IV. Further characteristics may be shared among
Falcomurus species, such as the presence of only two enlarged distal mac on the anterior face of the ventral tube [
15] (p. 81, Figure 11, and the remarks we present on
F. chilikaensis) and elements of the lateral chaetotaxy of the posterior head and trunk segments, which were omitted in [
15,
17]. On the other hand, the Ant. II undivided described for
F. chilikaensis,
D. litoreus and
D. halophilus is not stable in the genus, and the two new species described here have the Ant. II subdivided, as seen in
Dicranocentrus,
Pseudodicranocentrus and
Sinodicranocentrus [
8].
A detailed comparison among the species is presented in F. hilli sp. nov. remarks. Further notes on the morphology of Falcomurus are presented in the first discussion topic.
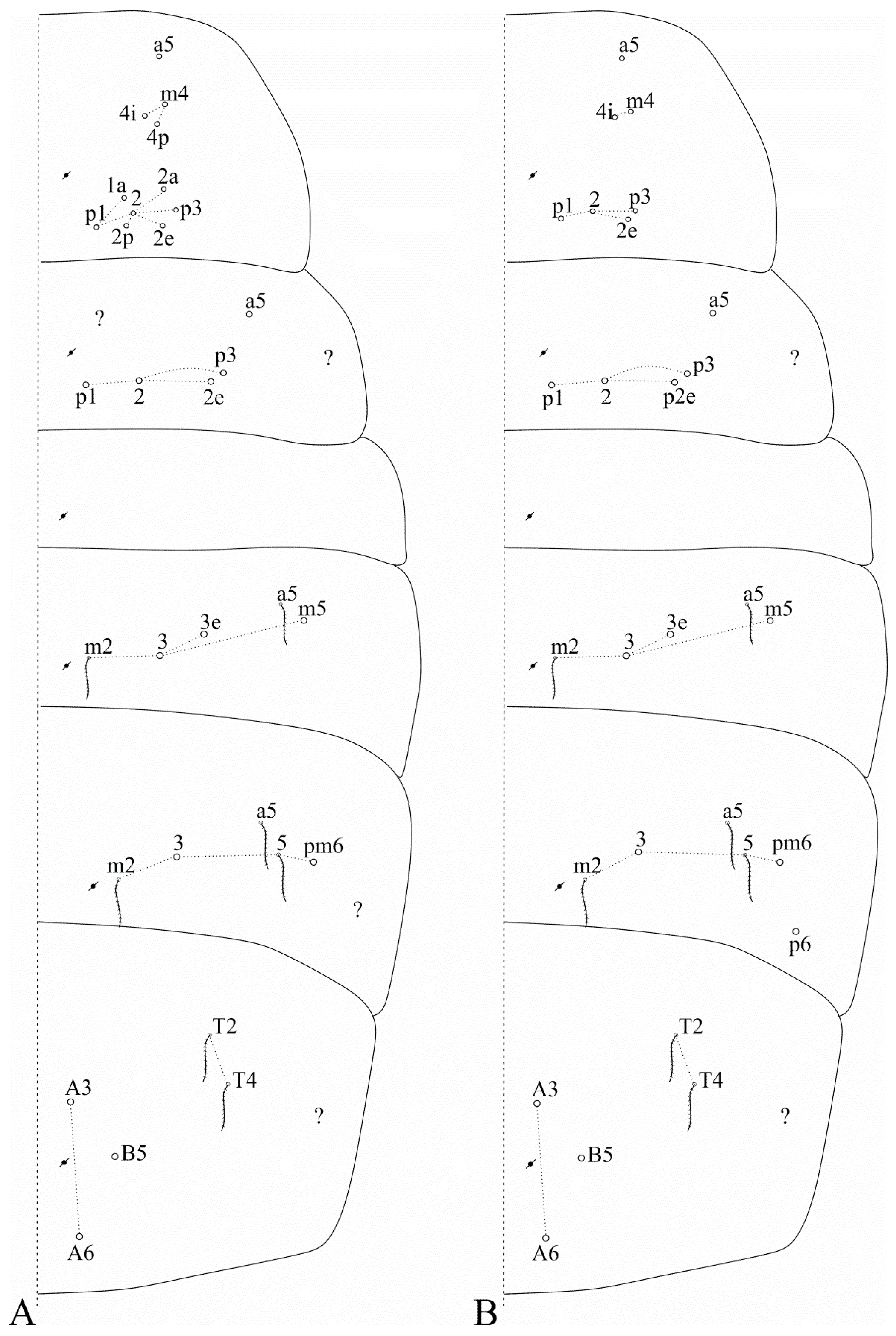
3.2. Falcomurus chilikaensis Mandal, 2018
Falcomurus chilikaensis Mandal, 2018 [
15] (pp. 77–83, Figures 1–17), India, Odisha state, Ganjam district, Rambha town, nearby Sabulia village (original description).
Diagnosis: Dark blue/violet pigments over antennae, dorso-anterior head and tibiotarsi, light blue/violet pigments on femurs and Abd. VI. Ant. IV with a pin projection. Ant. II undivided. Labral
a2 chaeta normal, not enlarged as
a1. Dorsal head with
Ps2 mac (
Figure 2A); post-labial quadrangle with smooth chaetae. Th. II with three
m4 and seven
p mac, respectively (
Figure 3A). Abd. IV with 3 central mac (possibly
A3,
A5 and
B5) (
Figure 3A). Trochanteral organ with 16–18 chaetae. Tibiotarsi proximally scaled. Unguiculi lacking the outer tooth. Dorsal manubrium with short smooth chaetae (adapted from [
15]).
Remarks: It is quite probable that Mandal’s description of F. chilikaensis includes misinterpretations and omissions regarding the dorsal chaetotaxy and other morphological aspects, which we were able to track and correct after reviewing the other species. In this sense, it is worth noting we did not examine the F. chilikaensis type series.
For the dorsal head macrochaetotaxy [
15] (p. 79, Figure 6), the common group of
A0,
A2–
3,
A5 mac seen in several Heteromurinae and all other
Falcomurus species were not clearly represented, and the marked
A0 actually belongs to the antennal series. So we considered in the anterior region the antennal series with three mac (originally represented only by one side), but quite possibly with more chaetae near the eyes, as we observed in the other taxa, and the original three anterior chaetae as misinterpretations of
A0,
A2–
3, while
M3 is actually
A6. Regarding the
M and
S series, the original drawing is not entirely symmetrical, so it is not clear if there are polymorphic chaetae, absent as mac in one of the sides. Since the author omitted mac on both sides in this drawing, we considered the absent mac of these series only as omissions and not as polymorphic chaetae, and such series were entirely represented in the left side of the original drawing. Concerning the posterior head, we believe the most posterior region was disregarded by the author, possibly due to the absence or reduction of mac, and the original drawing represents
Pa5, almost universal to Entomobryoidea,
Pm3 and
Pp3, seen in all other
Falcomurus species, plus
Ps2, exclusive of
F. chilikaensis. It is also quite possible that
F. chilikaensis has interocular chaetae, such as the other species and most, if not all, Entomobryoidea; but, they were overseen due to the presence of dark pigments over the eyepatches [
15] (p. 79, Figure 3). On the labrum, Mandal’s representation of
a1 chaeta clearly marks it as enlarged on the left side, as described by [
17] for
D. litoreus and
D. halophilus, while
a2 is apparently similar to the others [
15] (p. 80, Figure 8).
For the dorsal trunk chaetotaxy [
15] (p. 80, Figure 7), we considered the Th. II main chaetotaxy as it was presented by the author, since it fits the
a and
m series of the other species (but with
m4p), while the
p series shows some secondary multiples of
p1–
2 mac, differently from other
Falcomurus taxa but as seen in part of
Dicranocentrus and some other Orchesellidae [
6,
7,
8,
29,
32,
33]. However, to Th. III until Abd. IV, Mandal’s representation of the dorsal mac encompasses chaetae from the left and right sides at the same time, being more complete on the right side. On the Th. III there are six posterior mac represented on the original drawing, but they are likely the left and right sets of
p1–
2e, seen in all other species of the genus. So, the two superior mac on the right side represent
p3 and
a5, also seen in all other species of
Falcomurus. The upper
a5 mac marked by the author could represent the
a1 mac seen in scaleless Orchesellidae, like
Orchesella [
32] (p. 152). However, since the
a1 mic in the new species occurs near to the pseudopore, next to
m1, a feature seen in several other Heteromurini [
3,
8,
27,
33,
34], we disregarded Mandal’s representation of this mac, considering it as a doubtful feature. On the Abd. II, Mandal’s description shows 3 bothriotricha; however, the Entomobryoidea present 2 + 2 bothriotricha on this segment [
1,
3,
32]. In this segment, the author actually represents, from the left to the right: the left
m3 mac, left
m2 bothriotrichum, right
m2 bothriotrichum, right
m3–
3e mac, right
a5 bothriotrichum and right
m5 mac, all also seen in the other species of the genus. A similar mistake was seen in the Abd. III, which originally represents, from the left to the right: the left
m3 mac (left
m2 bothriotrichum was overseen, but its alveolus is smaller than the mac ones), right
m2 bothriotrichum, right
m3 mac, right
a5 and
m5 bothriotricha and right
pm6 mac, similar to its congeners. In this segment, it is quite possible that the
p6 mac is also present, and it was not represented since it did not fit the drawing. The Abd. IV interpretation was quite puzzling, since the original drawing was different from the expected disposition of
Dicranocentrus and
Heteromurus central mac [
29,
33,
37].
Falcomurus halophilus comb. nov. also has three central mac on Abd. IV,
A3,
A6 and
B5, while the other species of the genus have
A6 as a mes, so we believe the original drawing of
F. chilikaensis shows the chaetae inverted, and they are actually the same three. Once again, the left side is incomplete, as for the most trunk segments, so we disregarded a possible polymorphic state of
A6 and considered the Abd. IV with three internal mac.
Regarding other characteristics, in Ant. IV we disregarded the presence of the antennal bulb, as noted by [
8], but considered the left side structure as a pin projection (see [
15] (p. 79, Figure 5) and [
27] (p. 4, Figure 2B)). Mandal’s description noted his species had all legs scaled; however, the author did not remark on the presence or absence of scales on any particular segment of the legs. In his depiction of the tibiotarsi, the superior (proximal) region has “spine-like” structures, which most likely represent the scales alveoli, as they are somewhat pointed [
15] (p. 81, Figure 12). The habitus photograph of
F. chilikaensis also suggest such scales are present, as seen in the right fore and the left hind legs [
15] (p. 78, Figure 1). In addition, all specimens of
Falcomurus we revised do not have short spines on the tibiotarsi. Because of this, we considered the tibiotarsi of
F. chilikaensis proximally scaled, as we observed in
F. litoreus comb. nov. and
F. hilli sp. nov. Finally, the drawing of the ventral tube corpus in Mandal’s description [
15] (p. 81, Figure 11) possibly represents the lateral region, which is devoid of chaetae in many Entomobryoidea and also in other
Falcomurus species. So, the right side of the drawing possibly represents the two distal macrochaetae (as in [
17] (p. 314, Figure 78)) and the left side, one of the outer posterior ciliate chaetae.
After our reinterpretation of the original F. chilikaensis description, we believe the type material should be re-examined and the taxon redescribed, to confirm its morphology is similar to other Heteromurinae taxa as herein discussed. In this sense, our diagnosis, notes and drawings of this species should be taken as provisional, until further data based on the type specimens are published.
3.3. Falcomurus halophilus (Mari-Mutt, 1985) comb. nov.
Dicranocentrus halophilus Mari-Mutt, 1985 [
17] (pp. 315, 320, Figures 79–82), Papua New Guinea, Morobe, Lae (original description).
Diagnosis: Dark blue/violet pigments over all antennal segments, tibiotarsi, dorso-anterior head and around the mouth, light blue/violet uniformly distributed over trunk, other leg segments, ventral tube corpus and manubrium. Ant. IV without pin projection. Ant. II undivided, without scales. Labral
a1–
2 chaetae enlarged, distal labrum with two distinct pointed papillae. Dorsal head with
Pm1,
Pp1,
Pp5 and
Pe3? mac, without
An1a and
Ps2 mac, interocular area with five ciliate chaetae or four ciliate chaetae and one scale (
Figure 2B). Mandible apex curved to the midline. Post-labial quadrangle without smooth chaetae, anterior post-labial region with a pair of smooth chaetae near the labium. Th. II with two
m4 and four
p mac, respectively (
Figure 3B). Abd. IV with three central mac (
A3,
A6 and
B5) (
Figure 3B). Unguiculi with the outer tooth. Dorsal manubrium without smooth chaetae (adapted from [
17]).
Remarks: Dicranocentrus halophilus is herein transferred to Falcomurus since it shares characteristics with other species of the genus, especially the absence of A1, M3–4, S0 and S2 mac on dorsal head, the overall reduction of main dorsal macrochaetotaxy on trunk, including the Abd. I lacking mac, ungues short and thick, internally with only the basal paired teeth, unguiculi excavated, dens lacking spines, proximally with a falcate ciliate mac, and mucro lacking the mucronal spine.
Our diagnosis of
F. halophilus comb. nov. was based on the original description, since we did not revise the type specimens. As Mari-Mutt’s description of this species noted mostly its dissimilarities with
F. litoreus comb. nov., we used some data of the latter species to complement the diagnosis of
F. halophilus comb. nov. [
17] (pp. 319–320). On the other hand, the exact number of trochanteral spines and the presence of scales on the tibiotarsi and ventral tube anterior face, seen in our revision of
F. litoreus comb. nov. (see the next species), were not considered for
F. halophilus comb. nov., as they were not mentioned by the author. Concerning the interocular chaetotaxy of this species, the author described it with one outer and three inner chaetae plus a scale, similarly to
F. litoreus comb. nov. However, in his drawing [
17] (p. 315, Figure 80), there are only three interocular chaetae and the scale. We believe both Mari-Mutt species have a similar interocular chaetotaxy. Even so, the author regarded the interocular scale as a chaeta in one specimen [
17] (p. 320).
3.4. Falcomurus litoreus (Mari-Mutt, 1985) comb. nov.
Dicranocentrus litoreus Mari-Mutt, 1985 [
17] (pp. 313–314, 318–320, Figures 64–78, The Philippines, Mindoro, Puerto Galera (original description).
Examined material: Holotype (sex unknown) and 4 paratypes (one female, others unknown). The Philippines, Mindoro, Puerto Galera, on the beach, 27–29.xii.1979. All deposited now at MVMA.
Diagnosis: Dark blue/violet pigments over distal Ant. Ib, II and entire III (IV unknown), dorso-anterior head and tibiotarsi, light blue/violet pigments on Th. I, coxae, trochanters and distal ventral tube corpus; trunk devoid of pigments or brownish, with a median unpigmented line from Th. II to Abd. III. Ant. II undivided, with scales. Labral a1–2 chaetae enlarged, m2 normal, two distal labrum papillae pointed but weakly developed. Dorsal head with Pp1 (also as mic) and Pe3? mac, without An1a, Ps2, Pm1 and Pp5 mac, interocular area with four ciliate chaetae and one scale or three ciliate chaetae and two scales. Mandible apex curved to the midline. Post-labial quadrangle without smooth chaetae, anterior post-labial region with a pair of smooth chaetae near the labium. Th. II with two m4 and four central p mac, respectively. Abd. IV with two central mac (A3 and B5). Ventral tube densely scaled, lateral flap with about six smooth and 26 ciliate chaetae. Trochanteral organ with 27–32 chaetae. Tibiotarsi proximally scaled. Unguiculi with the outer tooth. Dorsal manubrium without short smooth chaetae, manubrial plate with 11–13 chaetae.
Additions to the original description: Ventral head and anterior face of the ventral tube densely covered by scales, tibiotarsi scaled proximally.
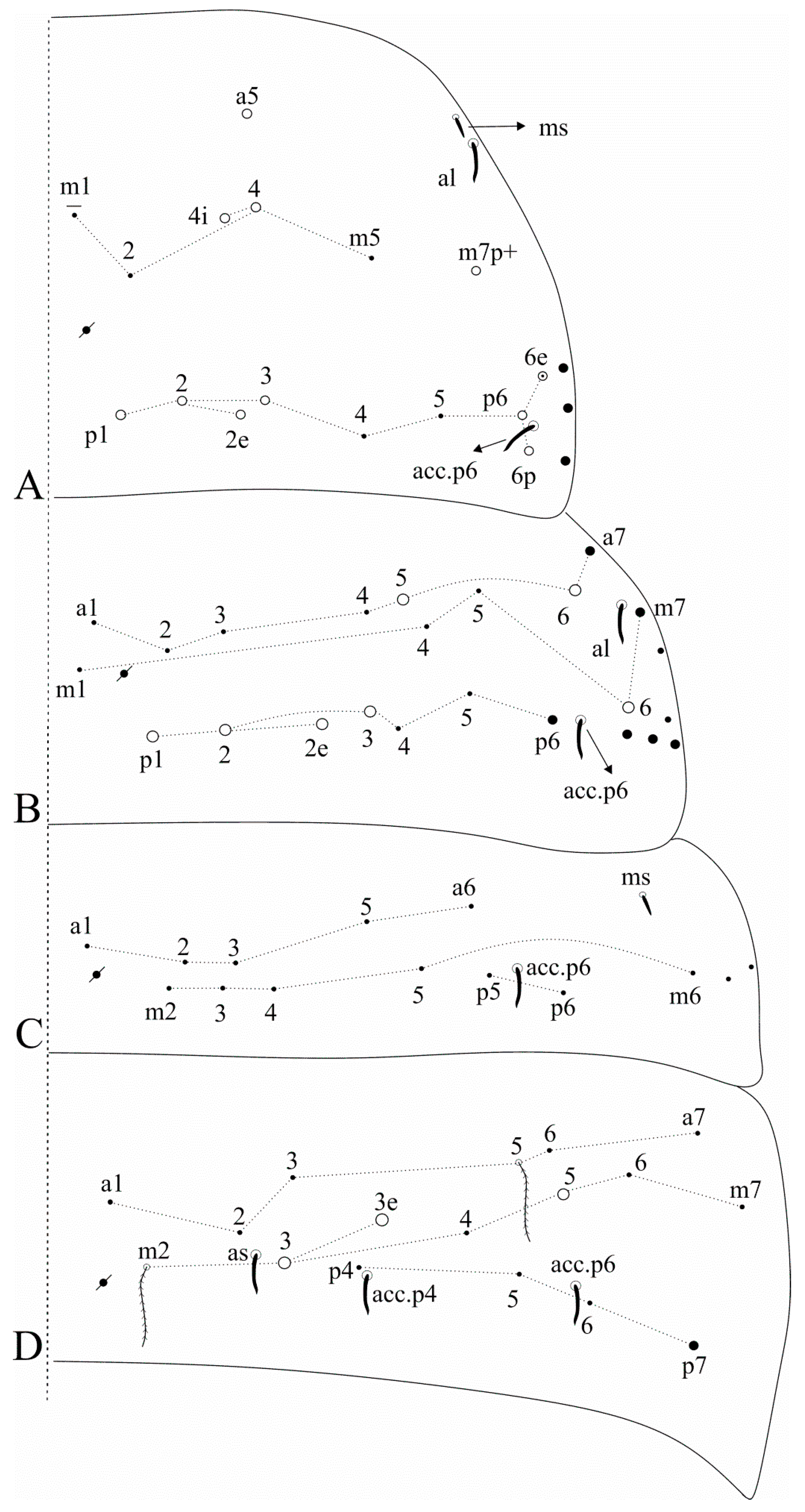
Head (
Figure 4). Two inner pointed labral papillae present, but underdeveloped. Interocular field with four mes (
s,
t,
r and
p) plus
v as a scale,
s also as a scale in one side of one specimen. Head dorsal macrochaetotaxy antennal (
An) row with 6–7 mac, including
An3,
An1a absent, anterior (
A) row with five (
A0,
A2–
3,
A5–
6), medio-ocellar (
M) row with two (
M1–
2),
M0 mic present; sutural (
S) row with six (
S1,
S3–
6), post-occipital anterior (
Pa) row with one (
Pa5), post-occipital medial with one (
Pm3), post-occipital posterior (
Pp) with 1–2 (
Pp1 and
Pp3,
Pp1 also as mic) and post-occipital external with one (
Pe3?) mac; further details are represented in
Figure 4A. Mandible apex somewhat curved to the midline (
Figure 4B). Maxillary outer lobe basal chaeta acuminate, slender (not erect or spiniform) and rough, shorter than the apical one, sublobal plate with four chaeta-like appendages, subequal in size (
Figure 4C). Labium with six main papillae (
H plus
A–
E), with 2, 0, 5, 0, 4, 4 guard chaetae, respectively, with five proximal chaetae subequal in length (
an1–
3,
p2–
3); labium unscaled, labial basomedial (submentum or labial triangle) and basolateral (mentum) fields with chaetae
a1–
5,
m2,
e and
l2 smooth,
l2 also ciliate ([
17], p. 319), basomedian field with 5–7 ciliate chaetae (analysed specimen with seven), including
M1 and
R, basolateral
L1 ciliate, subequal in length to
l2. Post-labial region with several scales and ciliate chaetae, except for the two smooth anterior chaetae nearby the labium, post-labial quadrangle with two subequal ciliate chaetae (
Figure 4D).
Trunk dorsal chaetotaxy (
Figure 5 and
Figure 6). Tergal S-chaetotaxy from Th. II to Abd. V as 2,2|1,3,3,+,3 for sens, and 1,0|1,0,1,0,0 for ms. Th. II, excluding the anterior collar, with one anterior (
a5), three medial (
m4–
4i and a multiple of
m7p row far from the collar) and seven posterior (
p1–
3,
p6–
6e) mac (
Figure 5A). Th. III with two anterior (
a5–
6), two medial (
m6–
7) and four posterior (
p1–
3) mac,
m1 mic not seen but possibly present (
Figure 5B). Abd. I lacking mac,
p5 mic present (
Figure 5C). Abd. II with three medial (
m3–
3e,
m5) mac (
Figure 5D). Abd. III with two medial (
m3 and
pm6) and one posterior (
p6) mac, one mic without clear homology near
pm6,
a1–
3 and
m3e mic not seen, but at least
a1–
3 possible present, lateral tergum with at least eight extra chaetae (
Figure 6A). Abd. IV with two central (
A3 and
B5) and 10 lateral (
D2–
3,
E2–
4,
F1–
2,
Fe2–
3, plus an external one) mac,
as sens internal to
A3,
ps nearby
D3p,
Ae6–
7 only seen by alveoli, lateral tergum with at least 17 mic and six mes without clear homologies (
Figure 6B). Abd. V main chaetotaxy with one anterior (
a6), four medial (
m2–
3,
m5,
m5e), five posterior (
p1,
p3–
ap6) plus
p5a? mac, with several secondary chaetae (only few represented in the drawing), sensillar pattern similar to
Heteromurus (as in [
3,
8]) (
Figure 6C). More details on the trunk’s idio and S-chaetotaxy are represented in
Figure 5 and
Figure 6.
Trunk appendages (
Figure 7). Ungues lateral paired teeth present and underdeveloped. Ventral tube anteriorly without smooth chaetae, with scales from the basis to the apex, with 12–17 ciliate chaetae of different sizes plus the distal two mac (
Figure 7A). Posterior face densely covered by ciliate chaetae and some central smooth chaetae, not entirely clear in the analysed specimens. Lateral flaps with about six smooth and 26 ciliate chaetae each, four distal smooth chaetae larger than the others (
Figure 7B). Manubrial plate with 11–13 ciliate chaetae and five small pseudopores (
Figure 7C).
Remarks: Dicranocentrus litoreus is herein transferred to Falcomurus as it shares with other species mainly the following combination of characters: absence of A1, M3–4, S0 and S2 mac on the dorsal head; the reduction of the main dorsal macrochaetotaxy on the trunk, as described in the genus diagnosis; a foot complex with a short unguis holding internally only the proximal paired teeth; unguiculi excavated; dens devoid of spines, with the typical proximal falcate mac seen in the genus; and the mucronal spine absent.
Our revision of
F. litoreus comb. nov. type material confirmed most of the data provided by [
17], with few corrections: the antennal series could present one more mac and the posterior head could also present
Pp1 as mac (seen only in one side of one specimen), while the ventral tube anterior region does not hold smooth chaetae [
17] (p. 314, Figure 78). We also could confirm the Ant. II undivided and the interocular element
v as a scale, as noted by the author [
17] (p. 318). We disagree with the original description of the mandible apex, described as elongated. In this case, it is quite similar to
F. pulukokos sp. nov. in length, but it is somewhat curved internally.
Unfortunately, we could only revise the dorsal chaetotaxy of the trunk in one specimen, which was damaged near the midline, so we could not confirm the presence of some internal primary mic nor identify further polymorphic chaetae. Due to the poor preservation of all five analysed slides, we could not properly revise the antennae, ventral tube posterior face and ventro-distal manubrium.
3.5. Falcomurus pulukokos sp. nov. Bellini, Souza and Greenslade
Type material: Holotype: female in slide (MVMA), along with two specimens of Dicranocentrus sp.: Australia, Cocos and Keeling Islands, West Island (12°09′ S, 96°49′ E), v–vi.2005, leaf litter light trap near the beach, A. Yen coll. Paratype in slide (MVMA): 1 female, along with two specimens of Dicranocentrus sp., same data of holotype.
Diagnosis: Ant. IV without pin projection, Ant. II subdivided in IIa and IIb, with few scales. Labral a1–2, m2 chaetae enlarged, two inner labral papillae present and well developed. Dorsal head with An1a, Pm1, Pp1 and Pe3? mac, without Ps2 and Pp5 mac, interocular area with three ciliate chaetae and one scale. Mandible apex normal, not elongated, narrow or curved. Post-labial quadrangle with two smooth chaetae, anterior post-labial region with a pair of smooth chaetae near the labium. Th. II with two m4 and four central p mac, respectively. Abd. IV with two central mac (A3 and B5). Ventral tube without scales, lateral flap with about 22 smooth chaetae. Trochanteral organ with at least 20 chaetae. Tibiotarsi unscaled. Unguiculi with the outer tooth. Dorsal manubrium without short smooth chaetae, manubrial plate with 7–8 chaetae.
Description: Body length (head + trunk) of holotype = 1.36 mm; range of type series length = 1.28–1.36 mm; average body length = 1.32 mm. Colour pattern unknown. Coarsely ciliate apically rounded or truncate scales present on Ant. I–II, head (dorsally and ventrally), dorsal trunk, legs (with exception of tibiotarsi), ventral and lateral manubrium and dens. Ventral tube unscaled.
Head (
Figure 8). Antennae shorter than body, ratio antennae:body of holotype = 1:1.84; Ant. III slightly smaller than Ant. IV, antennal ratio Ant. Ia–IV of holotype = 1:7:1.6:9.6:20:21. Ant. IV and III typically annulated. Ant. IV without a subapical organite, pin projection or any clear differentiated apical sens, only with normal sens and ciliate chaetae. Ant. III sense organ with two swollen sensory rods, three guard sensilla (all nearby the sensory rods) plus at least four surrounding regular sens (
Figure 8A). Ant. II subdivided in IIa and IIb, IIb with few scales, ventral face not entirely clear, with a single long smooth chaeta on the upper central region, dorsally with two apical sensory rods. Two inner pointed labral papillae present and well developed (
Figure 8B). Prelabral chaetae smooth and longer than the labral ones, labral
a1–
2 and
m2 enlarged,
m2 slightly longer than other labral chaetae (
Figure 8C). Eyepatches lenses A–F subequal in size, G–H smaller than the others, with four interocular elements:
s,
t and
p chaetae, the latter as a mac, and
v as a scale. Head dorsal macrochaetotaxy antennal (
An) row with seven, including
An1a, anterior (
A) row with five (
A0,
A2–
3,
A5–
6), medio-ocellar (
M) row with two (
M1–
2),
M0 mic present; sutural (
S) row with six (
S1,
S3–
6), post-occipital anterior (
Pa) row with one (
Pa5), post-occipital medial with two (
Pm1 and
Pm3), post-occipital posterior (
Pp) with two (
Pp1 and
Pp3) and post-occipital external with one (
Pe3?) mac; further details are represented in
Figure 8D. Mandibles normal, not remarkably elongated, narrow or curved, with 4–5 apical incisive teeth (
Figure 8E). Maxillae capitulum similar to
F. litoreus comb. nov. [
17] (p. 314, Figure 75). Labium with six main papillae (
H plus
A–
E), with 2, 0, 5, 0, 4, 4 guard chaetae, respectively, papilla
E lateral process finger-shaped and short, not reaching the papilla apex; with five proximal chaetae subequal in length (
an1–
3,
p2–
3); labium unscaled, labial basomedial (submentum or labial triangle) and basolateral (mentum) fields with chaetae
a1–
5,
m2,
e and
l2 smooth, basomedian field with 3–5 ciliate chaetae including
M1 and
R, basolateral
L1 ciliate, subequal in length to
l2 (
Figure 8F,G). Maxillary outer lobe basal chaeta acuminate, slender (not erect or spiniform) and rough, shorter than the apical one, ratio basal chaeta:apical chaeta of holotype = 1:1.15; sublobal plate with four chaeta-like appendages, subequal in size (
Figure 8H). Post-labial chaetotaxy with four smooth chaetae, two on the post-labial quadrangle and two on the first post-labial anterior row, and about 97 ciliate chaetae with different sizes (
Figure 8I).
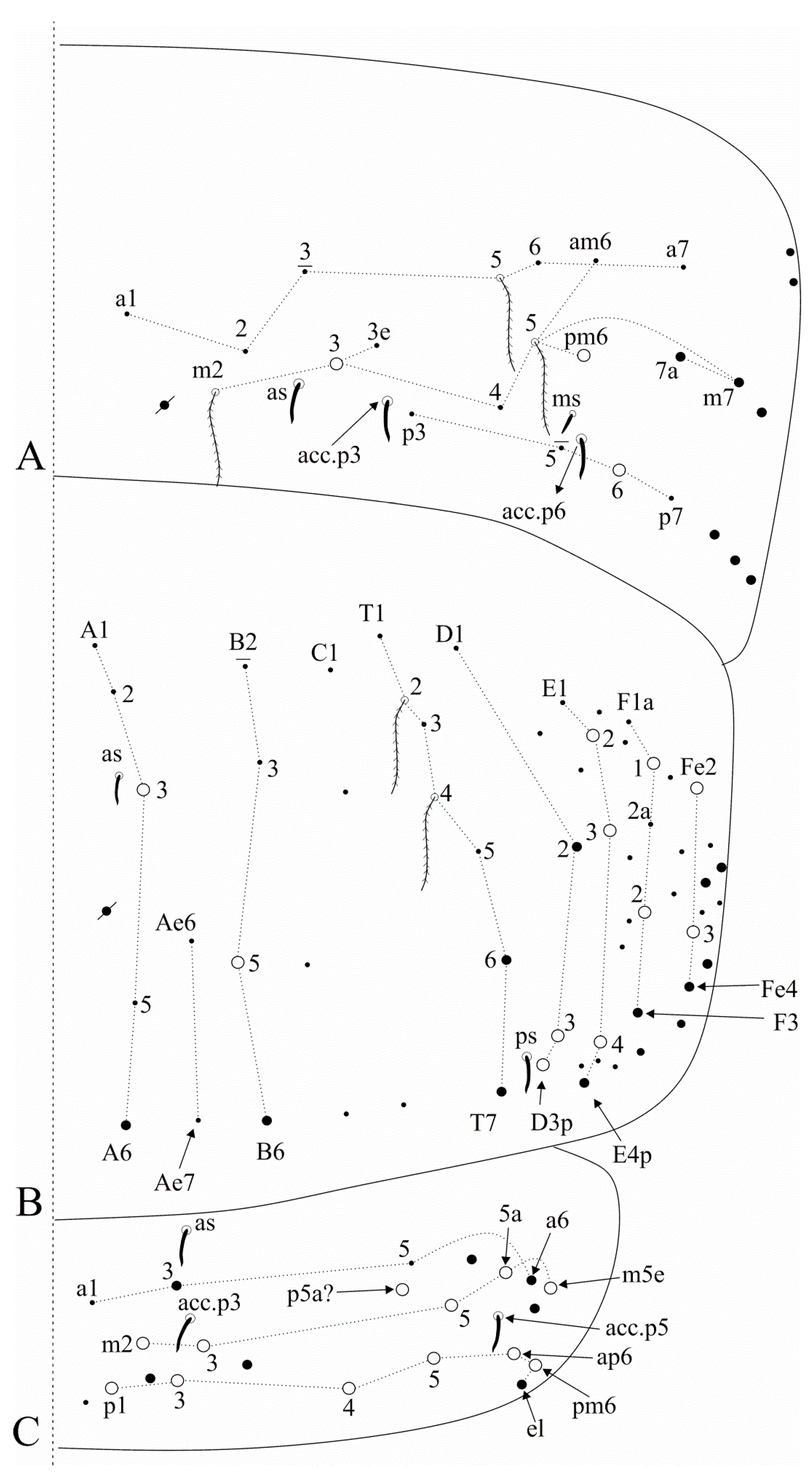
Trunk dorsal chaetotaxy (
Figure 9 and
Figure 10). Tergal S-chaetotaxy from Th. II to Abd. V as 2,2|1,3,3,+,3 for sens, and 1,0|1,0,1,0,0 for ms. Th. II, excluding anterior collar, with one anterior (
a5), three medial (
m4–
4i and a multiple of
m7p row far from the collar) and 6–7 posterior (
p1–
3,
p6–
6e,
p6e also as mic) mac (
Figure 9A). Th. III with two anterior (
a5–
6), one medial (
m6) and four posterior (
p1–
3) mac (
Figure 9B). Abd. I lacking mac,
p5 mic present (
Figure 9C). Abd. II with three medial (
m3–
3e,
m5) mac (
Figure 9D). Abd. III with two medial (
m3 and
pm6) and one posterior (
p6) mac, three mic without clear homologies, lateral tergum with at least 11 extra chaetae (
Figure 10A). Abd. IV with two central (
A3 and
B5) and 7–8 lateral (
D3,
E2–
4,
F1–
2 and
Fe2–
3,
Fe2 also as mic) mac,
as sens internal to
A3,
ps nearby
D3p, central region with at least 13 long sens (possibly many more) plus five mes (
A6,
Ae6,
B6 and two without clear homologies), lateral tergum with at least 12 mic and 10 mes without clear homologies (
Figure 10B). Ratio Abd. III:IV of holotype 1:1.63. Abd. V with five medial (
m2–
3,
m5–
5e), five posterior (
p1,
p3–
ap6) plus
p5a? mac plus several secondary chaetae, sensillar pattern similar to
Heteromurus (as in [
3,
8]) (
Figure 10C). More details on the trunk’s idio and S-chaetotaxy are represented in
Figure 9 and
Figure 10.
Trunk appendages (
Figure 11). Trochanteral organ unclear, with at least 20 spine-like smooth chaetae. Femurs and tibiotarsi lacking extra smooth chaetae. Tenent-hairs capitate, slightly shorter than ungues; tibiotarsus III smooth inner distal chaeta present; anterior and posterior pretarsal chaetae present and well developed; ungues with a pair of lateral underdeveloped teeth on proximal 1/3, inner side with only the two basal paired teeth on proximal 1/3, internal lamellae only merging at the apex, dorsal tooth present, almost in the middle of the unguis; unguiculi excavated, all lamellae smooth, except for the postero-external with an external tooth in its distal half (
Figure 11A). Ventral tube anteriorly without smooth chaetae, with two distal mac (the inner larger than the outer one), plus 15–16 ciliate chaetae of different sizes (
Figure 11B). Posterior face unclear, but apparently with several lateral ciliate chaetae and few centro-distal smooth chaetae. Lateral flaps with about 22 smooth chaetae each, three of them larger than the others (
Figure 11C). Tenaculum corpus with a single clearly ciliate chaeta, each ramus with four teeth. Manubrium and dens lacking smooth chaetae of any size. Manubrial plate with 7–8 chaetae, the two internal slightly larger than the others, plus at least three small pseudopores, possibly more (
Figure 11D). Ventro-distal manubrium with two inner ciliate chaetae plus about eight scales (
Figure 11E). Dorso-proximal dens with a typical sinuous ciliate mac (
Figure 11F), mucro apical tooth larger than the basal one, mucronal spine typically absent (
Figure 11G).
Etymology: The epithet refers to the Malay’s name for the Keeling territory.
Remarks: A detailed comparison of Falcomurus species is presented in the remarks of the next species.
3.6. Falcomurus hilli sp. nov. Bellini, Souza and Greenslade
Type material: Holotype: female in slide (MVMA): Australia, the Coral Sea Islands Territory, Marion Reef, Carola Cay, intertidal zone (19°06′ S, 152°23′ E), 29.xi.1982, pitfall trap, L. Hill coll. Paratypes: five females in slides (MVMA), same data of holotype.
Other analysed material: One subadult male in slide (MVMA): Australia, the Coral Sea Islands Territory, Lihou Reef, National Native Reserve, Georgina Cay (7°25′ S, 151°40′ E), 2.viii.1984, L. Hill coll. Two juveniles in one slide (MVMA): Australia, the Coral Sea Islands Territory, Herald Cays, Southwest Cay (17°00′ S, 149°08′ E), v.2007, beach, pitfall trap, P. Greenslade coll.
Diagnosis: Ant. IV without pin projection, Ant. II subdivided in IIa and IIb, with scales. Labral a1–2, m0–2 chaetae enlarged, four labral papillae present and well developed. Dorsal head with An1a, Pm1, Pp1, Pp5 and Pe3? and mac, without Ps2 mac, interocular area with three ciliate chaetae and one scale. Mandible apex normal, not elongated, narrow or curved. Post-labial region without smooth chaetae, including the post-labial quadrangle. Th. II with two m4 and four central p mac, respectively. Abd. IV with two central mac (A3 and B5). Ventral tube without scales, lateral flap with about nine ciliate and six smooth chaetae. Trochanteral organ with 14–17 chaetae. Tibiotarsi scaled proximally. Unguiculi with the outer tooth. Dorsal manubrium without short smooth chaetae, manubrial plate with six chaetae.
Description: Body length (head + trunk) of holotype = 1.54 mm; range of type series adults (n = 4) length = 1.54–1.66 mm; average body length of adults (n = 4) = 1.6 mm. Colour pattern unknown. Coarsely ciliate apically rounded or truncate scales present on Ant. I–II, head (dorsally and ventrally), dorsal trunk, legs (including proximal tibiotarsi), ventral and lateral manubrium and dens. Ventral tube unscaled.
Head (
Figure 12). Antennae shorter than body, ratio antennae:body of holotype = 1:2.3; Ant. III slightly smaller than Ant. IV, antennal ratio Ant. Ia–IV of holotype = 1:5.6:1.2:9:17:20. Ant. IV and III typically annulated. Ant. IV without a subapical organite, pin projection or any clear differentiated apical sens, only with normal sens and ciliate chaetae. Ant. III sense organ with normal (not swollen) sensory rods, three guard sensilla plus at least five surrounding sens of different sizes (
Figure 12A). Ant. II subdivided in IIa and IIb, IIb with few scales, dorsally with two apical sensory rods, ventrally with a single long smooth chaeta on the central region, plus ordinary ciliate chaetae and sens with different sizes and shapes (
Figure 12B,C). Four labral papillae present and well developed, lateral papillae large and well marked (
Figure 12D). Prelabral chaetae smooth and longer than the labral ones, labral
a1–
2 and
m0–
m2 enlarged,
m series slightly longer than other labral chaetae (
Figure 12E). Eyepatches lenses A–F subequal in size, G–H smaller than the others, with four interocular elements:
s,
t and
p chaetae, the latter as a mes, and
v as a scale. Head dorsal macrochaetotaxy antennal (
An) row with seven, including
An1a, anterior (
A) row with five (
A0,
A2–
3,
A5–
6), medio-ocellar (
M) row with two (
M1–
2),
M0 mic present; sutural (
S) row with six (
S1,
S3–
6), post-occipital anterior (
Pa) row with one (
Pa5), post-occipital medial with two (
Pm1 and
Pm3), post-occipital posterior (
Pp) with three (
Pp1,
Pp3 and
Pp5) and post-occipital external with one (
Pe3?) mac; further details are represented in
Figure 12F. Mandibles normal, not remarkably elongated, narrow or curved, similar to
F. pulukokos sp. nov. (as in
Figure 8E). Maxillae normal. Labium with six main papillae (
H plus
A–
E), with 2, 0, 5, 0, 4, 4 guard chaetae respectively, papilla
E lateral process finger-shaped and short, not reaching the papilla apex; with five proximal chaetae subequal in length (
an1–
3,
p2–
3,
an1 and
p2 slightly enlarged); labium unscaled, labial basomedial (submentum or labial triangle) and basolateral (mentum) fields with chaetae
a1–
5,
m2 and
e smooth,
l2 smooth or ciliate (smooth on holotype), basomedian field with 4–6 ciliate chaetae including
M1 and
R, basolateral
L1 ciliate, subequal in length to
l2. Maxillary outer lobe basal chaeta acuminate, slender (not erect or spiniform) and rough, shorter than the apical one, ratio basal chaeta:apical chaeta of holotype = 1:1.2; sublobal plate with four chaeta-like appendages, subequal in size. Post-labial chaetotaxy without smooth chaetae, with 77–78 ciliate chaetae of different sizes in the holotype, post-labial quadrangle with two subequal ciliate chaetae (
Figure 12G).
Trunk dorsal chaetotaxy (
Figure 13 and
Figure 14). Tergal S-chaetotaxy from Th. II to Abd. V as 2,2|1,3,3,+,3 for sens, and 1,0|1,0,1,0,0 for ms. Th. II, excluding anterior collar, with one anterior (
a5), three medial (
m4–
4i and a multiple of
m7p row far from the collar) and 7 posterior (
p1–
3,
p6–
6e) mac (
Figure 13A). Th. III with two anterior (
a5–
6), one medial (
m6) and four posterior (
p1–
3) mac (
Figure 13B). Abd. I lacking mac,
p5 mic present (
Figure 13C). Abd. II with three medial (
m3–
3e,
m5) mac (
Figure 13D). Abd. III with two medial (
m3 and
pm6) and one posterior (
p6) mac, lateral tergum with at least six extra chaetae (
Figure 14A). Abd. IV with two central (
A3 and
B5) and nine lateral (
D3–
3p,
E2–
4,
F1–
2 and
Fe2–
3) mac,
as sens internal to
A3,
ps nearby
D3p, central region with several long sens (not represented in the drawing), lateral tergum with at least 16 mic and five mes without clear homologies (
Figure 14B). Ratio Abd. III:IV of holotype 1:1.54. Abd. V main chaetotaxy with five medial (
m2–
3,
m5–
5e), five posterior (
p1,
p3–
ap6) plus
p5a? and
pm6 mac and several secondary chaetae (only few represented in the drawing), sensillar pattern similar to
Heteromurus (as in [
3,
8]) (
Figure 14C). More details on the trunk’s idio and S-chaetotaxy are represented in
Figure 13 and
Figure 14.
Trunk appendages (
Figure 15). Trochanteral organ with 14–17 spine-like smooth chaetae (
Figure 15A). Foot complex as in
F. pulukokos sp. nov. (
Figure 11A). Ventral tube anteriorly without smooth chaetae, with two distal mac (the inner larger than the outer one), plus 9–12 ciliate chaetae of different sizes (
Figure 15B). Posterior face not entirely clear, with at least 16 lateral ciliate chaetae and a pair of centro-distal long smooth chaetae. Lateral flaps with about nine ciliate and six smooth chaetae each, three smooth chaetae larger than the others (
Figure 15C). Tenaculum corpus with a single clearly ciliate chaeta, each ramus with four teeth. Manubrium and dens lacking smooth chaetae of any size. Manubrial plate with six chaetae, the two internal slightly larger than the others, plus five small pseudopores (
Figure 15D). Ventro-distal manubrium with two inner ciliate chaetae plus about 10 scales (
Figure 15E). Dorso-proximal dens with a typical sinuous ciliate mac (
Figure 11F), mucro apical tooth larger than the basal one, mucronal spine typically absent (
Figure 11G).
Etymology: The species honours the entomologist Lionel Hill, who collected the type material.
Remarks: The main useful features to separate
Falcomurus species are the subdivision of Ant. II, mandibles apex, labral papillae morphology, posterior dorsal head chaetotaxy, presence of smooth chaetae in the post-labial region, Th. II central chaetotaxy, presence of
A6 mac on Abd. IV, presence of the outer tooth on the unguiculi and manubrial dorsal chaetotaxy. In that regard,
F. chilikaensis is unique due the presence of
Ps2 mac on the dorsal head and Th. II with a complex central macrochaetotaxy, with three
m4 mac and seven mac on the
p1–
3 group, unguiculi without the outer tooth and dorsal manubrium with short smooth chaetae [
15]. This species also combines an undivided Ant. II, post-labial quadrangle with smooth chaetae and
A6 mac present on Abd. IV. Unfortunately, Mandal’s description did not provide data on the mandibles’ apex and labral papillae.
As noted by [
17], the main characteristics to distinguish
F. halophilus comb. nov. from
F. litoreus comb. nov. are the degree of labral papillae development (fully developed in
F. halophilus comb. nov. vs. weakly developed in
F. litoreus comb. nov.), posterior head macrochaetotaxy (with
Pm1 and
Pp5 mac in
F. halophilus comb. nov. vs. without in
F. litoreus comb. nov.) and Abd. IV central macrochaetotaxy (with three mac in
F. halophilus comb. nov. vs. two in
F. litoreus comb. nov.). The author also remarked on the presence of scales on Ant. II in the latter species (vs. absence in
F. halophilus comb. nov.); however, our analysis of the new species showed the number of scales in this segment is reduced, which could be easily overseen due to their fall. Some unknown or unclear features of
F. halophilus comb. nov., such as the presence of a ventral tube and tibiotarsi scales, ventral tube lateral flap’s chaetotaxy, the exact number of trochanteral organ smooth chaetae, among other characteristics, could be useful to better compare this species with its other congeners.
Falcomurus pulukokos sp. nov. and
F. hilli sp. nov. are the only species in the genus with the Ant. II subdivided [
15,
17]. They also differ from Mari-Mutt’s species by their normal mandible apex (curved in
F. halophilus comb. nov. and
F. litoreus comb. nov.) and interocular region with four elements, three chaetae and one scale (five in
F. halophilus comb. nov. and
F. litoreus comb. nov.). The new species differ from each other by their Ant. III sensory rods (swollen in
F. pulukokos sp. nov., normal in
F. hilli sp. nov.); labral papillae (four in
F. hilli sp. nov. and two in
F. pulukokos sp. nov.);
m1–
0 labral chaetae enlarged and dorsal head
Pp5 as mac only in
F. hilli sp. nov.; post-labial chaetotaxy without smooth chaetae on
F. hilli sp. nov. (with four in
F. pulukokos sp. nov.); and ventral tube’s lateral flaps without ciliate chaetae only in
F. pulukokos sp. nov.
More comparisons among the species are presented in
Table 1. An identification key to the
Falcomurus taxa based on our comparisons is presented in the next section.






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}