
Evolution of Adaptive Variation in the Mosquito Culex pipiens: Multiple Independent Origins of Insecticide Resistance Mutations


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
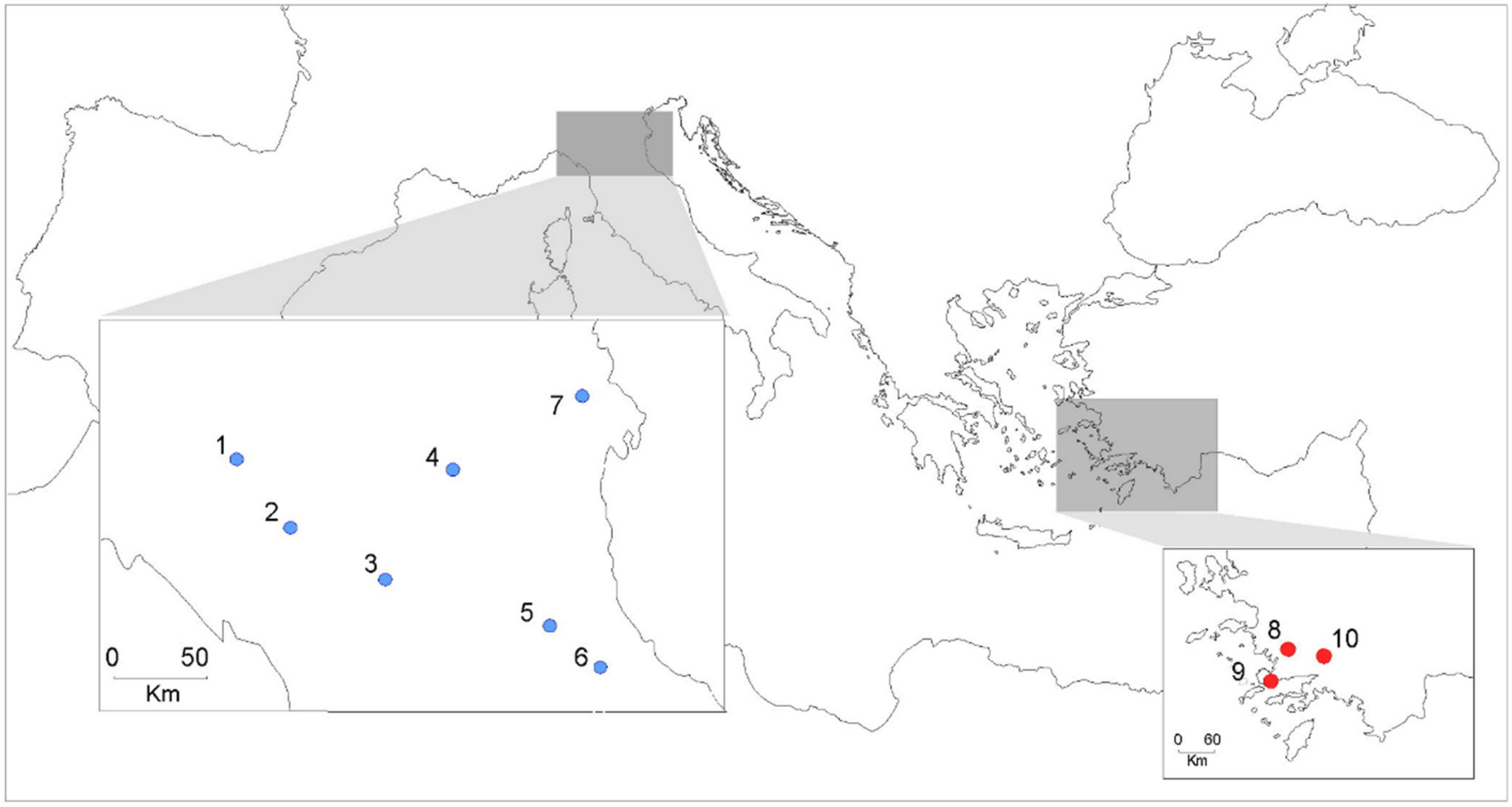
2.1. Study Sites and Samples
2.2. Laboratory Procedures
2.3. Sequence Analyses
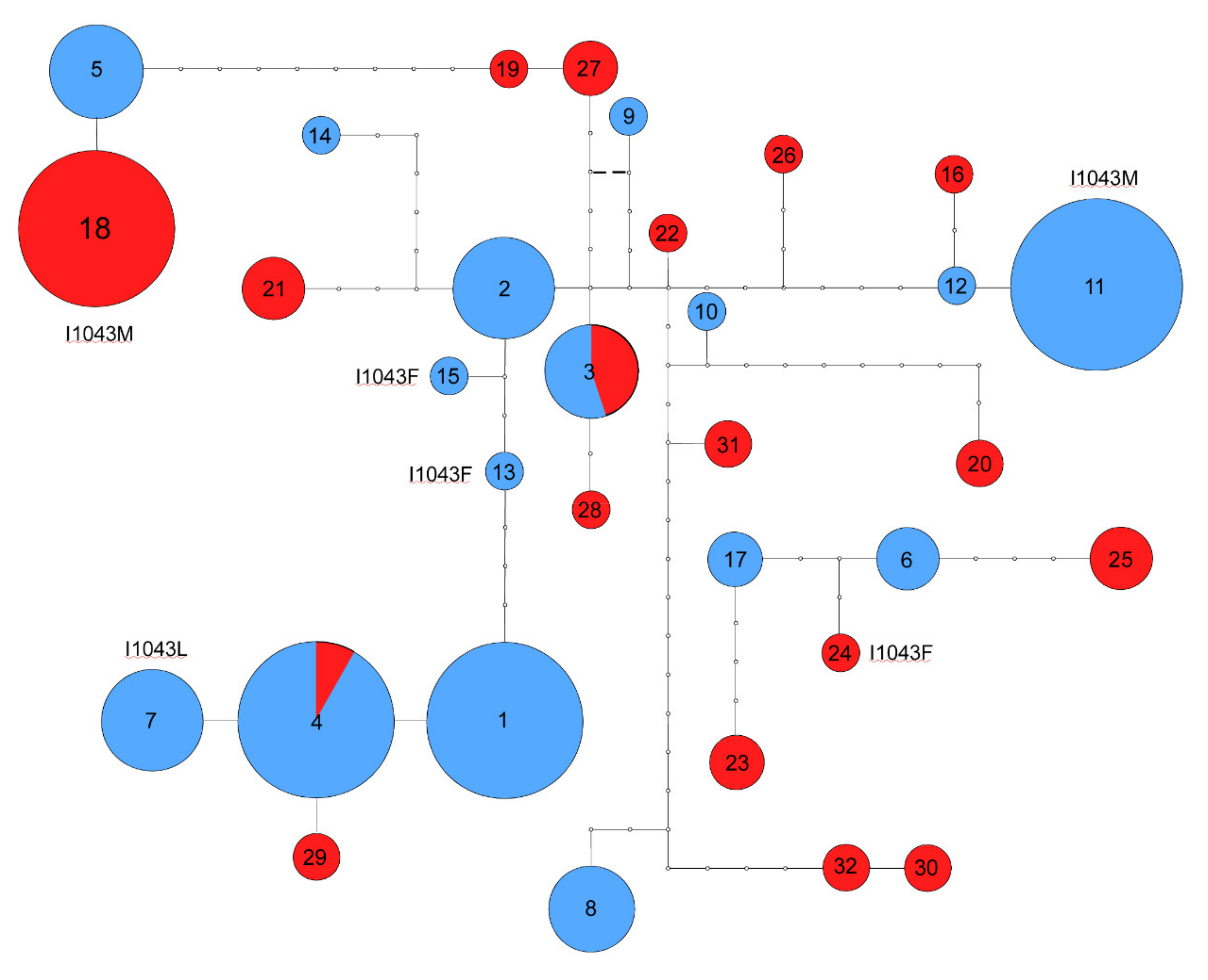
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ffrench-Constant, R.H. The molecular genetics of insecticide resistance. Genetics 2013, 194, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Feyereisen, R.; Dermauw, W.; Van Leeuwen, T. Genotype to phenotype, the molecular and physiological dimensions of resistance in arthropods. Pest. Biochem. Physiol. 2015, 121, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Ann. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, N.J.; Bass, C.; Dixon, A.; Neve, P. The evolutionary origins of pesticide resistance. Biol. Rev. 2019, 94, 135–155. [Google Scholar] [CrossRef]
- IRAC Insecticide MoA Classification. Available online: https://irac-online.org/modes-of-action/ (accessed on 5 July 2021).
- Tunaz, H.; Uygun, N. Insect growth regulators for insect pest control. Turk. J. Agric. For. 2004, 28, 377–387. [Google Scholar]
- Sun, R.; Liu, C.; Zhang, H.; Wang, Q. Benzoylurea chitin synthesis inhibitors. J. Agric. Food. Chem. 2015, 63, 6847–6865. [Google Scholar] [CrossRef] [PubMed]
- Douris, V.; Steinbach, D.; Panteleri, R.; Livadaras, I.; Pickett, J.A.; Leeuwen, T.V.; Nauen, R.; Vontas, J. Resistance mutation conserved between insects and mites unravels the benzoylurea insecticide mode of action on chitin biosynthesis. Proc. Natl. Acad. Sci. USA 2016, 113, 14692–14697. [Google Scholar] [CrossRef] [Green Version]
- Grigoraki, L.; Puggioli, A.; Mavridis, K.; Douris, V.; Montanari, M.; Bellini, R.; Vontas, J. Striking diflubenzuron resistance in Culex pipiens, the prime vector of West Nile Virus. Sci. Rep. 2017, 7, 11699. [Google Scholar] [CrossRef] [Green Version]
- Fotakis, E.A.; Mastrantonio, V.; Grigoraki, L.; Porretta, D.; Puggioli, A.; Chaskopoulou, A.; Osório, H.; Weill, M.; Bellini, R.; Urbanelli, S. Identification and detection of a novel point mutation in the Chitin Synthase gene of Culex pipiens associated with diflubenzuron resistance. PLoS Negl. Trop. Dis. 2020, 14, e0008284. [Google Scholar] [CrossRef]
- Porretta, D.; Fotakis, E.A.; Mastrantonio, V.; Chaskopoulou, A.; Michaelakis, A.; Kioulos, I.; Weill, M.; Urbanelli, S.; Vontas, J.; Bellini, R. Focal distribution of diflubenzuron resistance mutations in Culex pipiens mosquitoes from Northern Italy. Acta. Trop. 2019, 193, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Guz, N.; Çağatay, N.S.; Fotakis, E.A.; Durmuşoğlu, E.; Vontas, J. Detection of diflubenzuron and pyrethroid resistance mutations in Culex pipiens from Muğla, Turkey. Acta. Trop. 2020, 203, 105294. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, F.; Angel, G.; Geoffroy, B.; Hervy, J.P.; Rhaiem, A. The Mosquitoes of Europe/Les Moustiques d’Europe. Logiciel d’identification Et d’enseignement (CD-Rom); IRD Editions & EID MeÂditerrane Âe: Montpellier, France, 2001. [Google Scholar]
- Smith, J.L.; Fonseca, D.M. Rapid assays for identification of members of the Culex (Culex) pipiens complex, their hybrids, and other sibling species (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 2004, 70, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Navajas, M.; Lagnel, J.; Fauvel, G.; De Moraes, G. Sequence variation of ribosomal internal transcribed spacers (ITS) in commercially important phytoseiidae mites. Exp. Appl. Acarol. 1999, 23, 851–859. [Google Scholar] [CrossRef]
- Martin, D.P.; Lemey, P.; Posada, D. Analysing Recombination in Nucleotide Sequences. Mol. Ecol. Resour. 2011, 11, 943–955. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. Intraspecific gene genealogies: trees grafting into networks. Trends. Ecol. Evol. 2001, 16, 37–45. [Google Scholar] [CrossRef]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistics analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- Andreev, D.; Kreitman, M.; Phillips, T.W.; Beeman, R.W.; ffrench-Constant, R.H. Multiple origins of cyclodiene insecticide resistance in Tribolium castaneum (Coleoptera: Tenebrionidae). J. Mol. Evol. 1999, 48, 615e624. [Google Scholar] [CrossRef]
- Karasov, T.; Messer, P.W.; Petrov, D.A. Evidence that adaptation in Drosophila is not limited by mutation at single sites. PLoS Genet. 2010, 6, 1000924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinkevich, F.D.; Su, C.; Lazo, T.A.; Hawthorne, D.J.; Tingey, W.M.; Naimov, S.; Scott, J.G. Multiple evolutionary origins of knockdown resistance (kdr) in pyrethroid-resistant Colorado potato beetle, Leptinotarsa decemlineata. Pestic. Biochem. Physiol. 2012, 104, 192–200. [Google Scholar] [CrossRef]
- Cosme, L.V.; Gloria-Soria, A.; Caccone, A.; Powell, J.R.; Martins, A.J. Evolution of kdr haplotypes in worldwide populations of Aedes aegypti: Independent origins of the F1534C kdr mutation. PLoS Negl. Trop. Dis. 2020, 14, e0008219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellini, R.; Michaelakis, A.; Petric, D.; Schaffner, F.; Alten, B.; Angelini, P.; Aranda, C.; Becker, N.; Carrieri, M.; Di Luca, M.; et al. Practical management plan for invasive mosquito species in Europe: I. Asian tiger mosquito (Aedes albopictus). Travel. Med. Infect. Dis. 2020, 35, 101691. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Country | Code. Locality | No Larvae Analyzed | No. Homozygous Genotypes | Lat. | Long. | Year | chs-alleles |
|---|---|---|---|---|---|---|---|
| Italy | 1. Fiorenzuola | 28 | 12 | 44,9241667 N | 9,9130555 E | 2017 | a1(6), a2(10), a3(2), a4(2), a5(4) |
| 2. Parma | 26 | 11 | 44,768382 N | 10,319429 E | 2017 | a1(4), a2(6), a3(4), a4(4), a5(4) | |
| 3. Bologna | 25 | 11 | 44,533022 N | 11,344763 E | 2020 | a4(6), a5(2), a6(2), a7(6), a8(6) | |
| 4. Mizzana | 27 | 12 | 44,851931 N | 11,588654 E | 2017 | a1(8), a4(4), a6(6), a8(2), a9(2), a10(2) | |
| 5. Forlì | 25 | 13 | 44,2109124 N | 12,0552449 E | 2020 | a5(4), a11(16), a12(2), a13(2), a14(2) | |
| 6 Cesena | 25 | 12 | 44,1425853 N | 12,2596991 E | 2020 | a4(2), a7(8), a11(10), a15(2), a16(2) | |
| 7. Rovigo | 24 | 10 | 45,069454 N | 11,790022 E | 2018 | a1(4), a3(2), a4(2), a7(2), a8(4), a17(6) | |
| Turkey | 8. Milas | 30 | 16 | 36,796029 N | 28,796965 E | 2016 | a3(2), a18(10), a19(2), a20(2), a21(4), a22(2), a23(6), a24(2), a25(2) |
| 9. Bodrum | 28 | 15 | 37,058288 N | 27,233579 E | 2016 | a18(12), a21(4), a26(2), a27(6), a28(2), a29(2), a30(2) | |
| 10. Menteşe | 28 | 12 | 37,253156 N | 28,244305 E | 2016 | a3(4), a4(2), a20(2), a25(6), a29(2), a31(4), a32(4) |
| Allele | Variable sites | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
2 4 |
2 8 |
3 4 |
4 6 |
5 2 |
6 4 |
9 1 |
1 1 2 |
1 2 4 |
1 3 9 |
1 4 8 |
1 5 4 |
1 7 5 |
1 9 0 |
1 9 9 |
2 2 3 |
2 3 8 |
2 4 1 |
2 7 1 |
2 8 9 |
2 9 0 |
2 9 2 |
2 9 5 |
3 1 1 |
3 1 7 |
3 1 9 |
3 3 1 |
3 5 8 |
3 6 1 |
3 6 7 |
3 7 3 |
3 8 8 |
3 9 1 |
4 0 9 |
4 1 8 |
4 2 1 |
4 2 7 |
4 3 9 |
4 5 4 |
4 6 9 |
4 7 2 |
5 1 1 |
5 1 7 |
5 2 0 |
5 2 3 |
5 3 2 |
5 5 6 |
5 5 9 |
5 6 0 |
5 6 5 |
5 7 4 |
5 7 7 |
5 8 0 |
5 8 1 |
6 0 0 |
6 0 1 |
6 0 4 | |||
| a1 | T | T | G | T | G | G | G | G | C | C | C | A | C | G | G | C | C | T | C | T | A | C | G | T | T | G | C | T | C | T | G | G | A | G | A | A | C | G | C | A | A | A | C | C | C | C | G | C | T | C | C | G | G | T | T | G | C | ||
| a2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a3 | - | - | - | - | - | - | - | - | - | - | - | G | - | C | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a4 | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a5 | - | - | - | - | - | - | A | - | - | - | - | - | - | A | - | T | - | C | T | C | - | - | - | - | C | - | - | - | - | - | - | - | - | - | - | - | - | - | - | C | T | - | - | - | T | T | - | - | A | - | - | - | A | C | - | C | - | ||
| a6 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | C | - | - | - | - | C | - | T | C | - | C | - | A | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a7 | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | C | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a8 | C | C | - | - | A | - | - | T | - | T | - | - | - | C | A | T | - | - | - | C | - | - | - | - | C | - | - | - | A | C | - | A | - | A | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a9 | - | - | - | - | - | - | - | - | - | - | - | - | - | C | - | T | - | - | - | C | - | - | - | - | C | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | T | - | A | - | A | - | - | - | - | C | - | - | - | ||
| a10 | - | - | - | - | - | - | - | - | T | - | - | - | - | C | - | T | - | - | - | C | - | - | - | - | C | - | - | - | - | - | - | - | G | A | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a11 | C | C | - | - | - | A | - | - | - | - | - | - | T | A | - | T | - | - | - | C | - | G | A | - | C | - | - | - | A | - | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a12 | C | C | - | - | - | A | - | - | - | - | - | - | T | A | - | T | - | - | - | C | - | - | A | - | C | - | - | - | A | - | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a13 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | T | - | A | - | C | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a14 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | - | G | - | T | - | C | - | - | G | - | T | - | - | - | - | A | - | - | A | - | - | - | - | T | ||
| a15 | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | T | - | - | - | C | T | - | A | - | C | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a16 | C | C | - | - | - | A | - | - | - | - | - | - | T | A | - | T | - | - | - | C | - | - | A | - | C | - | - | - | A | - | - | - | - | A | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | C | - | C | - | ||
| a17 | - | - | - | - | - | - | - | - | - | - | - | - | - | C | - | T | - | - | - | C | - | - | - | - | C | - | - | C | - | C | - | A | - | A | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a18 | - | - | - | - | - | - | A | - | - | - | - | - | - | A | - | T | - | C | T | C | - | G | - | - | C | - | - | C | - | - | - | - | - | - | - | - | - | - | C | C | T | - | - | - | T | T | - | - | A | - | - | - | A | C | - | C | - | ||
| a19 | - | - | - | - | - | - | - | - | - | - | - | - | - | C | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | T | T | - | - | A | - | - | - | A | C | - | C | - | |||
| a20 | - | - | - | C | - | - | - | - | - | - | - | G | - | - | - | T | A | - | - | C | - | - | - | - | C | - | - | - | - | - | - | - | G | A | - | - | - | A | - | - | - | - | - | - | - | - | - | - | A | T | - | - | A | C | - | C | - | ||
| a21 | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | - | - | G | T | - | - | - | - | - | - | - | - | - | - | T | A | - | - | - | - | - | - | - | - | ||
| a22 | - | - | - | - | - | - | - | - | - | - | - | - | - | C | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | - | ||
| a23 | - | - | - | - | - | - | - | - | - | - | - | G | - | C | - | T | - | - | - | C | - | T | - | - | C | - | - | C | - | C | T | A | - | A | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | - | ||
| a24 | - | - | A | - | - | - | - | - | - | - | - | - | - | C | - | T | - | - | - | C | T | - | - | - | C | - | T | C | - | C | - | A | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a25 | C | C | - | - | - | - | - | - | T | - | - | - | - | - | - | T | - | - | T | C | - | - | - | - | C | - | T | C | - | C | - | A | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a26 | - | - | - | - | - | - | - | A | - | - | - | - | - | A | - | T | - | - | - | C | - | - | A | C | C | A | - | - | A | - | - | - | - | A | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a27 | - | - | - | - | - | - | - | - | - | - | - | - | - | C | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | T | T | - | - | A | - | - | - | A | C | - | - | - | ||
| a28 | - | - | - | - | - | - | - | - | - | - | - | G | - | C | - | T | - | - | - | C | - | - | A | - | C | - | - | - | - | - | - | - | - | - | - | - | T | - | - | - | - | - | T | - | - | C | - | - | - | - | A | - | - | - | - | - | - | ||
| a29 | - | - | - | - | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | C | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a30 | C | C | - | - | A | - | - | - | - | T | - | - | - | A | - | T | - | - | - | C | - | - | - | - | C | - | - | - | A | - | - | - | - | A | - | - | - | A | - | - | - | - | - | - | - | - | - | - | A | - | - | A | - | - | - | - | - | ||
| a31 | - | - | A | - | - | - | - | - | - | - | - | - | - | C | - | T | - | - | - | C | - | - | - | - | C | - | - | - | - | - | - | - | G | A | - | - | - | - | T | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | ||
| a32 | C | C | - | - | A | - | - | - | - | T | - | - | - | A | - | T | - | T | - | C | - | - | - | - | C | - | - | - | A | - | - | - | - | A | - | - | - | A | - | - | - | - | - | - | - | - | - | - | - | - | - | A | - | - | - | - | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastrantonio, V.; Porretta, D.; Lucchesi, V.; Güz, N.; Çağatay, N.S.; Bellini, R.; Vontas, J.; Urbanelli, S. Evolution of Adaptive Variation in the Mosquito Culex pipiens: Multiple Independent Origins of Insecticide Resistance Mutations. Insects 2021, 12, 676. https://doi.org/10.3390/insects12080676
Mastrantonio V, Porretta D, Lucchesi V, Güz N, Çağatay NS, Bellini R, Vontas J, Urbanelli S. Evolution of Adaptive Variation in the Mosquito Culex pipiens: Multiple Independent Origins of Insecticide Resistance Mutations. Insects. 2021; 12(8):676. https://doi.org/10.3390/insects12080676
Chicago/Turabian StyleMastrantonio, Valentina, Daniele Porretta, Valentina Lucchesi, Nurper Güz, Naciye Sena Çağatay, Romeo Bellini, John Vontas, and Sandra Urbanelli. 2021. "Evolution of Adaptive Variation in the Mosquito Culex pipiens: Multiple Independent Origins of Insecticide Resistance Mutations" Insects 12, no. 8: 676. https://doi.org/10.3390/insects12080676
APA StyleMastrantonio, V., Porretta, D., Lucchesi, V., Güz, N., Çağatay, N. S., Bellini, R., Vontas, J., & Urbanelli, S. (2021). Evolution of Adaptive Variation in the Mosquito Culex pipiens: Multiple Independent Origins of Insecticide Resistance Mutations. Insects, 12(8), 676. https://doi.org/10.3390/insects12080676