
Japanese Honeybees (Apis cerana japonica Radoszkowski, 1877) May Be Resilient to Land Use Change



Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Japanese Beekeeping
1.2. Aims and Scope
- How does land use in Japan affect the honey produced by A. cerana japonica?
- How does the time of year and location affect the pollen collected and honey produced by A. cerana japonica?
2. Materials and Methods
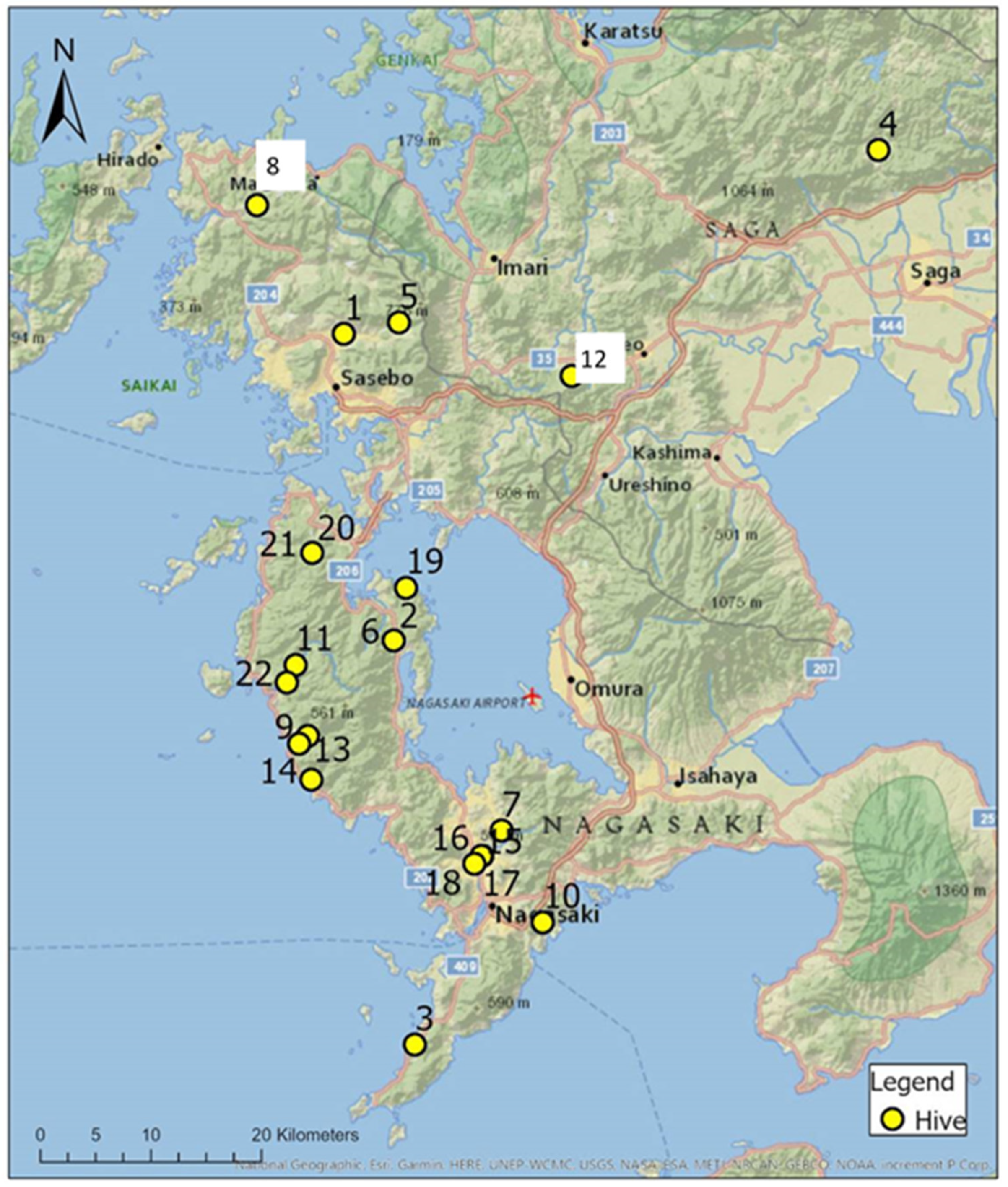
2.1. Honey Sampling
2.2. Pollen Sampling
2.3. Land Use Composition
2.4. Statistical Analysis
3. Results
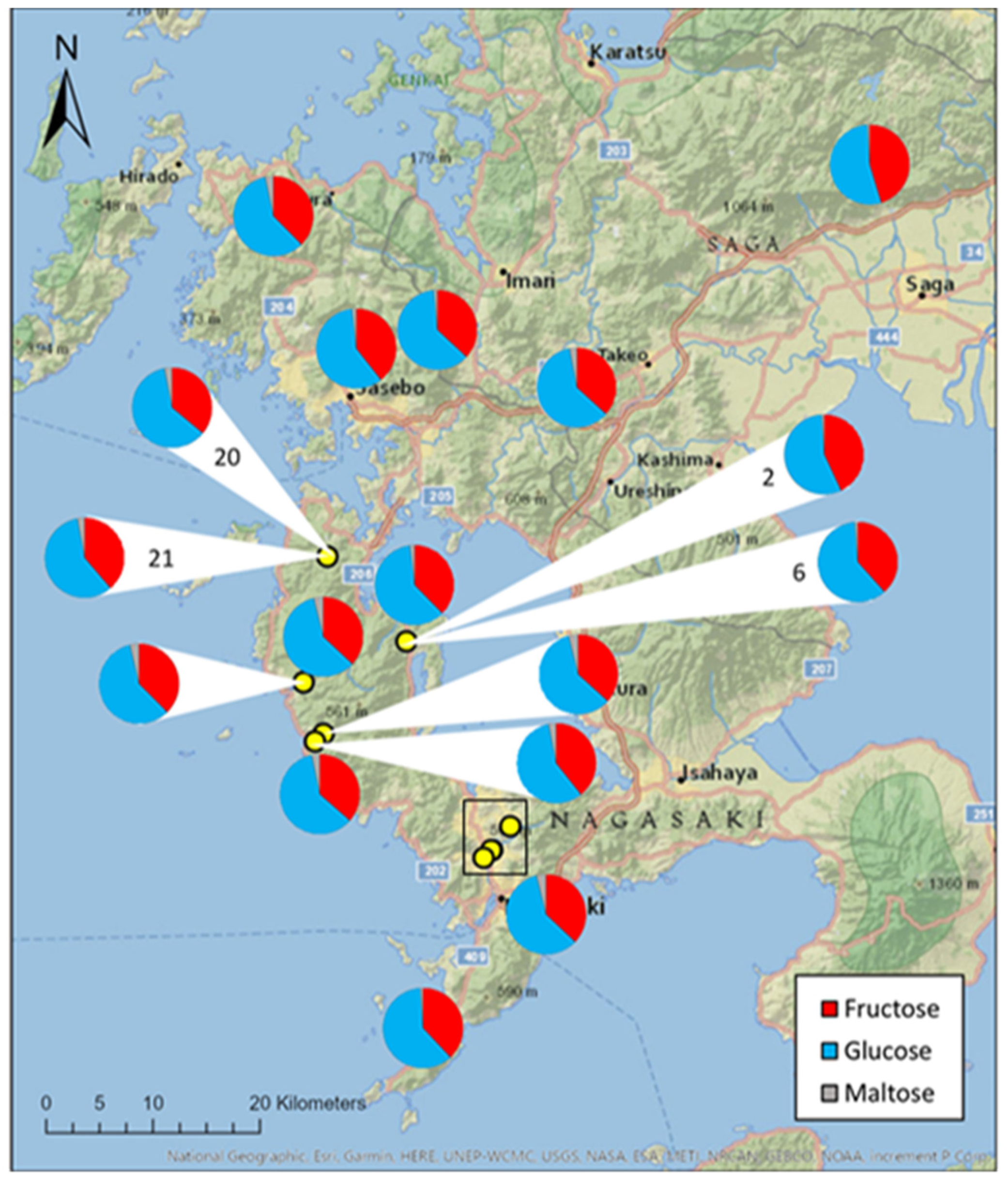
3.1. Honey Sugar Composition
3.2. Pollen Biotype Composition
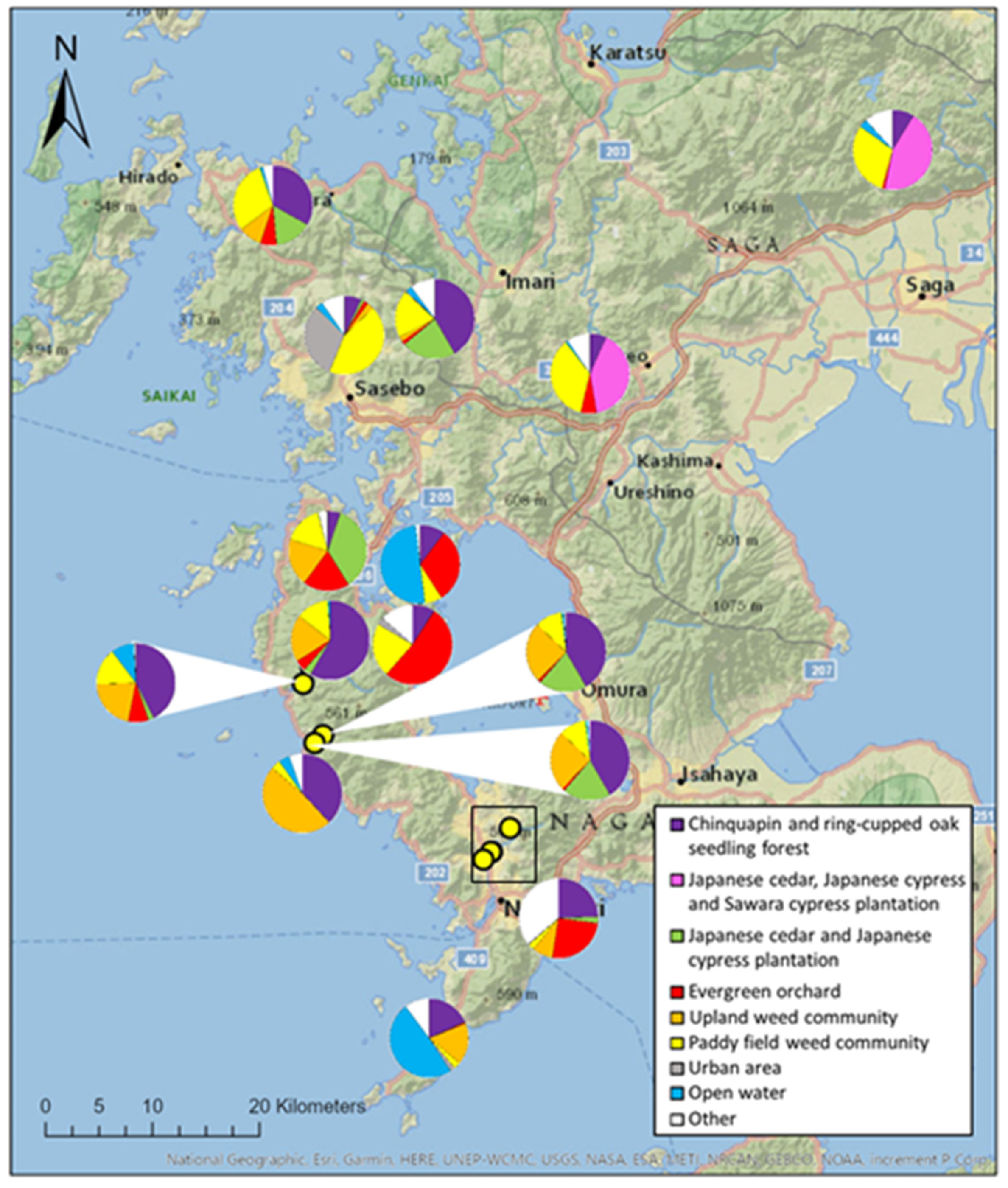
3.3. Land Use Composition
4. Discussion
4.1. The Importance of Honey Sugar Composition
4.2. Could Urban Land Use Be Less Detrimental?
4.3. Could Japanese Honeybees Be More Resilient?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sponsler, D.B.; Grozinger, C.M.; Hitaj, C.; Rundlöf, M.; Botías, C.; Code, A.; Lonsdorf, E.V.; Melathopoulos, A.P.; Smith, D.; Suryanarayanan, S.; et al. Pesticides and pollinators: A socioecological synthesis. Sci. Total. Environ. 2019, 662, 1012–1027. [Google Scholar] [CrossRef]
- Aznar-Sánchez, J.A.; Piquer-Rodríguez, M.; Velasco-Muñoz, J.F.; Manzano-Agugliaro, F. Worldwide research trends on sustainable land use in agriculture. Land Use Policy 2019, 87, 104069. [Google Scholar] [CrossRef]
- Vanbergen, A.J.; Garratt, M.P.; Vanbergen, A.J.; Baude, M.; Biesmeijer, J.C.; Britton, N.F.; Brown, M.J.F.; Brown, M.; Bryden, J.; Budge, G.E.; et al. Threats to an ecosystem service: Pressures on pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Hadley, A.S.; Betts, M.G. The effects of landscape fragmentation on pollination dynamics: Absence of evidence not evidence of absence. Biol. Rev. 2011, 87, 526–544. [Google Scholar] [CrossRef]
- Betts, M.G.; Hadley, A.S.; Kormann, U. The landscape ecology of pollination. Landsc. Ecol. 2019, 34, 961–966. [Google Scholar] [CrossRef] [Green Version]
- Menz, M.H.; Phillips, R.D.; Winfree, R.; Kremen, C.; Aizen, M.; Johnson, S.; Dixon, K. Reconnecting plants and pollinators: Challenges in the restoration of pollination mutualisms. Trends Plant Sci. 2011, 16, 4–12. [Google Scholar] [CrossRef]
- Hopwood, J.; Black, S.H.; Lee-Mӓder, E.; Charlap, A.; Preston, R.; Mozumder, K.; Fleury, S. Literature Review: Pollinator Habitat Enhancement and Best Management Practices in Highway Rights-of-way. Fed. Highw. Adm. 2015. Prepared by: The Xerces Society for Invertebrate Conservation in collaboration with ICF International. Available online: https://xerces.org/sites/default/files/2018-05/15-055_01_pollinators_BMPs_in_highway_ROW.pdf (accessed on 29 July 2021).
- Lowenstein, D.M.; Matteson, K.C.; Minor, E.S. Diversity of wild bees supports pollination services in an urbanized landscape. Oecologia 2015, 179, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Donkersley, P.; Rhodes, G.; Pickup, R.W.; Jones, K.C.; Power, E.F.; Wright, G.A.; Wilson, K. Nutritional composition of honeybee food stores vary with floral composition. Oecologia 2017, 185, 749–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sponsler, D.B.; Matcham, E.G.; Lin, C.-H.; Lanterman, J.L.; Johnson, R.M. Spatial and taxonomic patterns of honey bee foraging: A choice test between urban and agricultural landscapes. J. Urban Ecol. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Ruedenauer, F.A.; Raubenheimer, D.; Kessner-Beierlein, D.; Grund-Mueller, N.; Noack, L.; Spaethe, J.; Leonhardt, S.D. Best be(e) on low fat: Linking nutrient perception, regulation and fitness. Ecol. Lett. 2020, 23, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Danner, N.; Molitor, A.M.; Schiele, S.; Härtel, S.; Steffan-Dewenter, I. Season and landscape composition affect pollen foraging distances and habitat use of honey bees. Ecol. Appl. 2016, 26, 1920–1929. [Google Scholar] [CrossRef] [PubMed]
- Osborne, J.L.; Smith, A.; Clark, S.J.; Reynolds, D.R.; Barron, M.C.; Lim, K.S.; Reynolds, A.M. The Ontogeny of Bumblebee Flight Trajectories: From Naïve Explorers to Experienced Foragers. PLoS ONE 2013, 8, e78681. [Google Scholar] [CrossRef] [Green Version]
- Donkersley, P.; Elsner-Adams, E.; Maderson, S. A One-Health Model for Reversing Honeybee (Apis mellifera L.) Decline. Veter. Sci. 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Rader, R.; Bartomeus, I.; Garibaldi, L.A.; Garratt, M.P.D.; Howlett, B.; Winfree, R.; Cunningham, S.; Mayfield, M.; Arthur, A.D.; Andersson, G.; et al. Non-bee insects are important contributors to global crop pollination. Proc. Natl. Acad. Sci. USA 2015, 113, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Becher, M.A.; Osborne, J.L.; Thorbek, P.; Kennedy, P.J.; Grimm, V. REVIEW: Towards a systems approach for understanding honeybee decline: A stocktaking and synthesis of existing models. J. Appl. Ecol. 2013, 50, 868–880. [Google Scholar] [CrossRef] [Green Version]
- Donkersley, P. Trees for bees. Agric. Ecosyst. Environ. 2019, 270–271, 79–83. [Google Scholar] [CrossRef]
- Senapathi, D.; Goddard, M.; Kunin, W.E.; Baldock, K.C.R. Landscape impacts on pollinator communities in temperate systems: Evidence and knowledge gaps. Funct. Ecol. 2016, 31, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Powney, G.D.; Carvell, C.; Edwards, M.; Morris, R.K.A.; Roy, H.E.; Woodcock, B.A.; Isaac, N.J.B. Widespread losses of pollinating insects in Britain. Nat. Commun. 2019, 10, 1–6. [Google Scholar] [CrossRef]
- Ollerton, J. Pollinator Diversity: Distribution, Ecological Function, and Conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 353–376. [Google Scholar] [CrossRef] [Green Version]
- Eggleton, P. The State of the World’s Insects. Annu. Rev. Environ. Resour. 2020, 45, 61–82. [Google Scholar] [CrossRef]
- Hanley, N.; Breeze, T.; Ellis, C.; Goulson, D. Measuring the economic value of pollination services: Principles, evidence and knowledge gaps. Ecosyst. Serv. 2015, 14, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Wood, T.J.; Michez, D.; Paxton, R.J.; Drossart, M.; Neumann, P.; Gérard, M.; Vanderplanck, M.; Barraud, A.; Martinet, B.; Leclercq, N.; et al. Managed honey bees as a radar for wild bee decline? Apidologie 2020, 51, 1100–1116. [Google Scholar] [CrossRef]
- López-Uribe, M.M.; Ricigliano, V.A.; Simone-Finstrom, M. Defining Pollinator Health: Assessing Bee Ecological, Genetic, and Physiological Factors at the Individual, Colony, and Population Levels. Annu. Rev. Anim. Biosci. 2020, 8, 269–294. [Google Scholar] [CrossRef] [PubMed]
- Carreck, N.L.; Dietemann, V.; Neumann, P.; Ellis, J.D. The COLOSS BEEBOOK: Global standards in honey bee research. J. Apic. Res. 2020, 59, 1–4. [Google Scholar] [CrossRef]
- Menzel, R. The honeybee as a model for understanding the basis of cognition. Nat. Rev. Neurosci. 2012, 13, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Rivière, J.; Alaux, C.; Le Conte, Y.; Layec, Y.; Lozac’h, A.; Rodin, V.; Singhoff, F. Toward a complete agent-based model of a Honeybee colony. In Highlights of Practical Applications of Agents, Multi-Agent Systems, and Complexity: The PAAMS Collection; Springer: Cham, Switzerland, 2018; pp. 493–505. [Google Scholar]
- Bagheri, S.; Mirzaie, M. A mathematical model of honey bee colony dynamics to predict the effect of pollen on colony failure. PLoS ONE 2019, 14, e0225632. [Google Scholar] [CrossRef]
- Tatsuno, M.; Osawa, N. Flower visitation patterns of the coexisting honey bees Apis cerana japonica and Apis mellifera (Hymenoptera: Apidae). Èntomol. Sci. 2016, 19, 255–267. [Google Scholar] [CrossRef]
- Kohsaka, R.; Park, M.S.; Uchiyama, Y. Beekeeping and honey production in Japan and South Korea: Past and present. J. Ethn. Foods 2017, 4, 72–79. [Google Scholar] [CrossRef]
- Mitsumori, Y. An Analysis of Impact of Urban Beekeeping Projects on Community: Ginza Bee Projects brought not only bees, but also a more sophisticated image to Ginza. In Proceedings of the 2020 IEEE 8th R10 Humanitarian Technology Conference (R10-HTC), Kuching, Malaysia, 1–3 December 2020; pp. 1–7. [Google Scholar] [CrossRef]
- Ministry of Agriculture Forestry Fisheries. Situation of Beekeeping; 令和2年11月農林水産省 生産局 畜産部 (Reiwa Livestock Department, Production Bureau, Ministry of Agriculture, Forestry and Fisheries), Japan. 2020. Available online: https://www.maff.go.jp/j/chikusan/kikaku/lin/sonota/attach/pdf/bee-8.pdf (accessed on 29 July 2021).
- National Bee Unit. The Hive Count; FERA, York. 2021. Available online: nationalbeeunit.com/index.cfm?pageid=362 (accessed on 29 July 2021).
- Wells, H.; Wells, P.H. Optimal Diet, Minimal Uncertainty and Individual Constancy in the Foraging of Honey Bees, Apis mellifera. J. Anim. Ecol. 1986, 55, 881. [Google Scholar] [CrossRef]
- Yoshida, T. Nihonmitsubachi no Shiikuho to Seitai (Keeping Method and Ecology of Japanese Honey Bee); Tamagawa University Press: Tokyo, Japan, 2000. [Google Scholar]
- Nagasaki University Community Beekeepers. Personal Communication, 2021.
- Adams, C.J.; Boult, C.H.; Deadman, B.; Farr, J.M.; Grainger, M.; Manley-Harris, M.; Snow, M.J. Isolation by HPLC and characterisation of the bioactive fraction of New Zealand manuka (Leptospermum scoparium) honey. Carbohydr. Res. 2008, 343, 651–659. [Google Scholar] [CrossRef]
- Combarros-Fuertes, P.; Estevinho, L.M.; Teixeira-Santos, R.; Rodrigues, A.G.; Pina-Vaz, C.; Fresno, J.M.; Tornadijo, M.E. Antibacterial Action Mechanisms of Honey: Physiological Effects of Avocado, Chestnut, and Polyfloral Honey upon Staphylococcus aureus and Escherichia coli. Molecules 2020, 25, 1252. [Google Scholar] [CrossRef] [Green Version]
- Bucekova, M.; Jardekova, L.; Juricova, V.; Bugarova, V.; Di Marco, G.; Gismondi, A.; Leonardi, D.; Farkasovska, J.; Godocikova, J.; Laho, M.; et al. Antibacterial Activity of Different Blossom Honeys: New Findings. Molecules 2019, 24, 1573. [Google Scholar] [CrossRef] [Green Version]
- Escuredo, O.; Carmen Seijo, M. Honey: Chemical composition, stability and authenticity. Foods 2019, 8, 577. [Google Scholar] [CrossRef] [Green Version]
- Anupama, D.; Bhat, K.; Sapna, V. Sensory and physico-chemical properties of commercial samples of honey. Food Res. Int. 2003, 36, 183–191. [Google Scholar] [CrossRef]
- White, J.W., Jr.; Subers, M.H.; Schepartz, A.I. The identification of inhibine, the antibacterial factor in honey, as hydrogen peroxide and its origin in a honey glucose-oxidase system. Biochim. Biophys. Acta Bioenerg. 1963, 73, 57–70. [Google Scholar] [CrossRef]
- Brudzynski, K. A current perspective on hydrogen peroxide production in honey. A review. Food Chem. 2020, 332. [Google Scholar] [CrossRef]
- Ouchemoukh, S.; Schweitzer, P.; Bey, M.B.; Djoudad-Kadji, H.; Louaileche, H. HPLC sugar profiles of Algerian honeys. Food Chem. 2010, 121, 561–568. [Google Scholar] [CrossRef]
- Wang, J.; Kliks, M.M.; Jun, S.; Jackson, M.; Li, Q.X. Rapid Analysis of Glucose, Fructose, Sucrose, and Maltose in Honeys from Different Geographic Regions using Fourier Transform Infrared Spectroscopy and Multivariate Analysis. J. Food Sci. 2010, 75, C208–C214. [Google Scholar] [CrossRef]
- Giesecke, T.; Fontana, S.L.; Van Der Knaap, W.O.; Pardoe, H.S.; Pidek, I. From early pollen trapping experiments to the Pollen Monitoring Programme. Veg. Hist. Archaeobotany 2010, 19, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Donkersley, P.; Rhodes, G.; Pickup, R.W.; Jones, K.C.; Wilson, K. Honeybee nutrition is linked to landscape composition. Ecol. Evol. 2014, 4, 4195–4206. [Google Scholar] [CrossRef] [Green Version]
- Lau, P.; Bryant, V.; Ellis, J.D.; Huang, Z.Y.; Sullivan, J.; Schmehl, D.R.; Cabrera, A.R.; Rangel, J. Seasonal variation of pollen collected by honey bees (Apis mellifera) in developed areas across four regions in the United States. PLoS ONE 2019, 14, e0217294. [Google Scholar] [CrossRef]
- Jones, G.D. Ollen analyses for pollination research, acetolysis. J. Pollinat. Ecol. 2014, 13, 203–217. [Google Scholar] [CrossRef]
- Clermont, A.; Eickermann, M.; Kraus, F.; Hoffmann, L.; Beyer, M. Correlations between land covers and honey bee colony losses in a country with industrialized and rural regions. Sci. Total. Environ. 2015, 532, 1–13. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing v4.0.5; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Shimakura, M. Palynomorphs of Japanese plants. Spec. Publ. Osaka Mus. Nat. Hist. 1973, 5, 1–60. [Google Scholar]
- Steffan-Dewenter, I.; Kuhn, A. Honeybee foraging in differentially structured landscapes. Proc. R. Soc. B Boil. Sci. 2003, 270, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Nottebrock, H.; Schmid, B.; Mayer, K.; Devaux, C.; Esler, K.J.; Böhning-Gaese, K.; Schleuning, M.; Pagel, J.; Schurr, F.M. Sugar landscapes and pollinator-mediated interactions in plant communities. Ecography 2016, 40, 1129–1138. [Google Scholar] [CrossRef]
- Nzeako, B.C.; Hamdi, J. Antimicrobial potential of honey on some microbial isolates. Sultan Qaboos Univ. Med. J. 2000, 2, 75–79. [Google Scholar]
- Erler, S. Pharmacy bee hive—Use of natural drugs to increase pollinator health. In Proceedings of the 2016 International Congress of Entomology, Orlando, FL, USA, 26 September 2016. [Google Scholar] [CrossRef]
- Tauber, J.P.; Collins, W.R.; Schwarz, R.S.; Chen, Y.; Grubbs, K.; Huang, Q.; Lopez, D.; Peterson, R.; Evans, J.D. Natural Product Medicines for Honey Bees: Perspective and Protocols. Insects 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Erler, S.; Moritz, R.F.A. Pharmacophagy and pharmacophory: Mechanisms of self-medication and disease prevention in the honeybee colony (Apis mellifera). Apidologie 2015, 47, 389–411. [Google Scholar] [CrossRef] [Green Version]
- Fujlta, I. Determination of Maltose in Honey. Int. J. Food Sci. Nutr. Diet. 2012, 1, 1–2. [Google Scholar] [CrossRef]
- Ball, D.W. The Chemical Composition of Honey. J. Chem. Educ. 2007, 84. [Google Scholar] [CrossRef]
- Tangtorwongsakul, P.; Warrit, N.; Gale, G.A. Effects of landscape cover and local habitat characteristics on visiting bees in tropical orchards. Agric. For. Èntomol. 2017, 20, 28–40. [Google Scholar] [CrossRef]
- Harrison, T.; Gibbs, J.; Winfree, R. Anthropogenic landscapes support fewer rare bee species. Landsc. Ecol. 2017, 34, 967–978. [Google Scholar] [CrossRef]
- Nürnberger, F.; Steffan-Dewenter, I.; Härtel, S. Combined effects of waggle dance communication and landscape heterogeneity on nectar and pollen uptake in honey bee colonies. PeerJ 2017, 5. [Google Scholar] [CrossRef]
- Rupprecht, C.D.D. Informal Urban Green Space: Residents’ Perception, Use, and Management Preferences across Four Major Japanese Shrinking Cities. Land 2017, 6, 59. [Google Scholar] [CrossRef]
- O’Neil, J.A.; Gallagher, C.E. Determining What is Important in Terms of the Quality of an Urban Green Network: A Study of Urban Planning in England and Scotland. Plan. Pr. Res. 2014, 29, 202–216. [Google Scholar] [CrossRef]
- Sutter, L.; Jeanneret, P.; Bartual, A.M.; Bocci, G.; Albrecht, M. Enhancing plant diversity in agricultural landscapes promotes both rare bees and dominant crop-pollinating bees through complementary increase in key floral resources. J. Appl. Ecol. 2017, 54, 1856–1864. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.A.; Nimmo, D.G.; Cunningham, S.; Walker, K.; Bennett, A.F. The response of wild bees to tree cover and rural land use is mediated by species’ traits. Biol. Conserv. 2019, 231, 1–12. [Google Scholar] [CrossRef]
- Garbuzov, M.; Schürch, R.; Ratnieks, F.L. Eating locally: Dance decoding demonstrates that urban honey bees in Brighton, UK, forage mainly in the surrounding urban area. Urban Ecosyst. 2014, 18, 411–418. [Google Scholar] [CrossRef]
- Rupprecht, C.; Byrne, J.; Ueda, H.; Lo, A. ‘It’s real, not fake like a park’: Residents’ perception and use of informal urban green-space in Brisbane, Australia and Sapporo, Japan. Landsc. Urban Plan. 2015, 143, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Todorova, A.; Asakawa, S.; Aikoh, T. Preferences for and attitudes towards street flowers and trees in Sapporo, Japan. Landsc. Urban Plan. 2004, 69, 403–416. [Google Scholar] [CrossRef]
- Petrova, E.G.; Mironov, Y.V.; Aoki, Y.; Matsushima, H.; Ebine, S.; Furuya, K.; Petrova, A.; Takayama, N.; Ueda, H. Comparing the visual perception and aesthetic evaluation of natural landscapes in Russia and Japan: Cultural and environmental factors. Prog. Earth Planet. Sci. 2015, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Penick, C.A.; Crofton, C.A.; Appler, R.H.; Frank, S.D.; Dunn, R.R.; Tarpy, D.R. The contribution of human foods to honey bee diets in a mid-sized metropolis. J. Urban Ecol. 2016, 2. [Google Scholar] [CrossRef] [Green Version]
- Piroux, M.; Lambert, O.; Puyo, S.; Farrera, I.; Thorin, C.; L’Hostis, M.; Vigues, B.; Bastian, S. Correlating the pollens gathered by apis mellifera with the landscape features in western france. Appl. Ecol. Environ. Res. 2014, 12, 423–439. [Google Scholar] [CrossRef]
- Jiao, Y.; Ding, Y.; Zha, Z.; Okuro, T. Crises of Biodiversity and Ecosystem Services in Satoyama Landscape of Japan: A Review on the Role of Management. Sustainability 2019, 11, 454. [Google Scholar] [CrossRef] [Green Version]
- Haaland, C.; Bosch, C.K.V.D. Challenges and strategies for urban green-space planning in cities undergoing densification: A review. Urban For. Urban Green. 2015, 14, 760–771. [Google Scholar] [CrossRef]
- Hendriksma, H.P.; Shafir, S. Honey bee foragers balance colony nutritional deficiencies. Behav. Ecol. Sociobiol. 2016, 70, 509–517. [Google Scholar] [CrossRef]
- Anderson, K.E.; Carroll, M.J.; Sheehan, T.; Mott, B.M.; Maes, P.; Corby-Harris, V. Hive-stored pollen of honey bees: Many lines of evidence are consistent with pollen preservation, not nutrient conversion. Mol. Ecol. 2014, 23, 5904–5917. [Google Scholar] [CrossRef]
- Yoshida, T.; Saito, J.; Kajigaya, N. The mating flight times of native Apis cerana japonica Radoszkowski and introduced Apis mellifera L. in sympatric conditions. Apidologie 1994, 25, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Cai, F.; Si, A.; Zhang, S.; Tautz, J.; Chen, S. East Learns from West: Asiatic Honeybees Can Understand Dance Language of European Honeybees. PLoS ONE 2008, 3, e2365. [Google Scholar] [CrossRef] [Green Version]
- Wallberg, A.; Han, F.; Wellhagen, G.; Dahle, B.; Kawata, M.; Haddad, N.; Simões, Z.L.P.; Allsopp, M.H.; Kandemir, I.; De La Rúa, P.; et al. A worldwide survey of genome sequence variation provides insight into the evolutionary history of the honeybee Apis mellifera. Nat. Genet. 2014, 46, 1081–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sniderman, J.M.K.; Matley, K.A.; Haberle, S.; Cantrill, D.J. Pollen analysis of Australian honey. PLoS ONE 2018, 13, e0197545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnuchamy, R.; Bonhomme, V.; Prasad, S.; Das, L.; Patel, P.; Gaucherel, C.; Pragasam, A.; Anupama, K. Honey Pollen: Using Melissopalynology to Understand Foraging Preferences of Bees in Tropical South India. PLoS ONE 2014, 9, e101618. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Biotype | Possible Species and Family Name (s) | Description | Reference Image |
|---|---|---|---|
| 1 | Aralia elata (Araliaceae) | Small, circular/semi-circular |  |
| 3 | Firmiana simplex (Malvaceae) | Medium, dark, 3-way symmetry, rounded |  |
| 10 | Very small, light, circular/semi-circular. |  | |
| 14 | Medium, oblong, slightly pointed at ends, arcing lines through |  | |
| 17 | Lithocarpus edulis (Fagaceae) | Small, oval, lines arcing through. |  |
| 20 | Mallotus japonicus (Euphorbiaceae) Citrus/Fortunella crassifolia (Rutaceae) Dendropanax trifidus (Arialaceae) Euonymus japonicus (Celastraceae) | Similar to 16, but more indents, elongated. From end on: small-medium, three rounded sides, triangle inside, with points between indents. |  |
| Sugar | 1 km | 3 km | 5 km | |||
|---|---|---|---|---|---|---|
| rs/r Value | p-Value | rs/r Value | p-Value | rs/r Value | p-Value | |
| Fructose | -0.230 | 0.303 | 0.079 | 0.726 | 0.174 | 0.439 |
| Glucose | −0.095 | 0.675 | −0.051 | 0.822 | −0.297 | 0.179 |
| Maltose | 0.465 | 0.029 | 0.189 | 0.399 | 0.043 | 0.848 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donkersley, P.; Covell, L.; Ota, T. Japanese Honeybees (Apis cerana japonica Radoszkowski, 1877) May Be Resilient to Land Use Change. Insects 2021, 12, 685. https://doi.org/10.3390/insects12080685
Donkersley P, Covell L, Ota T. Japanese Honeybees (Apis cerana japonica Radoszkowski, 1877) May Be Resilient to Land Use Change. Insects. 2021; 12(8):685. https://doi.org/10.3390/insects12080685
Chicago/Turabian StyleDonkersley, Philip, Lucy Covell, and Takahiro Ota. 2021. "Japanese Honeybees (Apis cerana japonica Radoszkowski, 1877) May Be Resilient to Land Use Change" Insects 12, no. 8: 685. https://doi.org/10.3390/insects12080685
APA StyleDonkersley, P., Covell, L., & Ota, T. (2021). Japanese Honeybees (Apis cerana japonica Radoszkowski, 1877) May Be Resilient to Land Use Change. Insects, 12(8), 685. https://doi.org/10.3390/insects12080685