
Bombyx mori β-1,3-Glucan Recognition Protein 4 (BmβGRP4) Could Inhibit the Proliferation of B. mori Nucleopolyhedrovirus through Promoting Apoptosis

 and
and 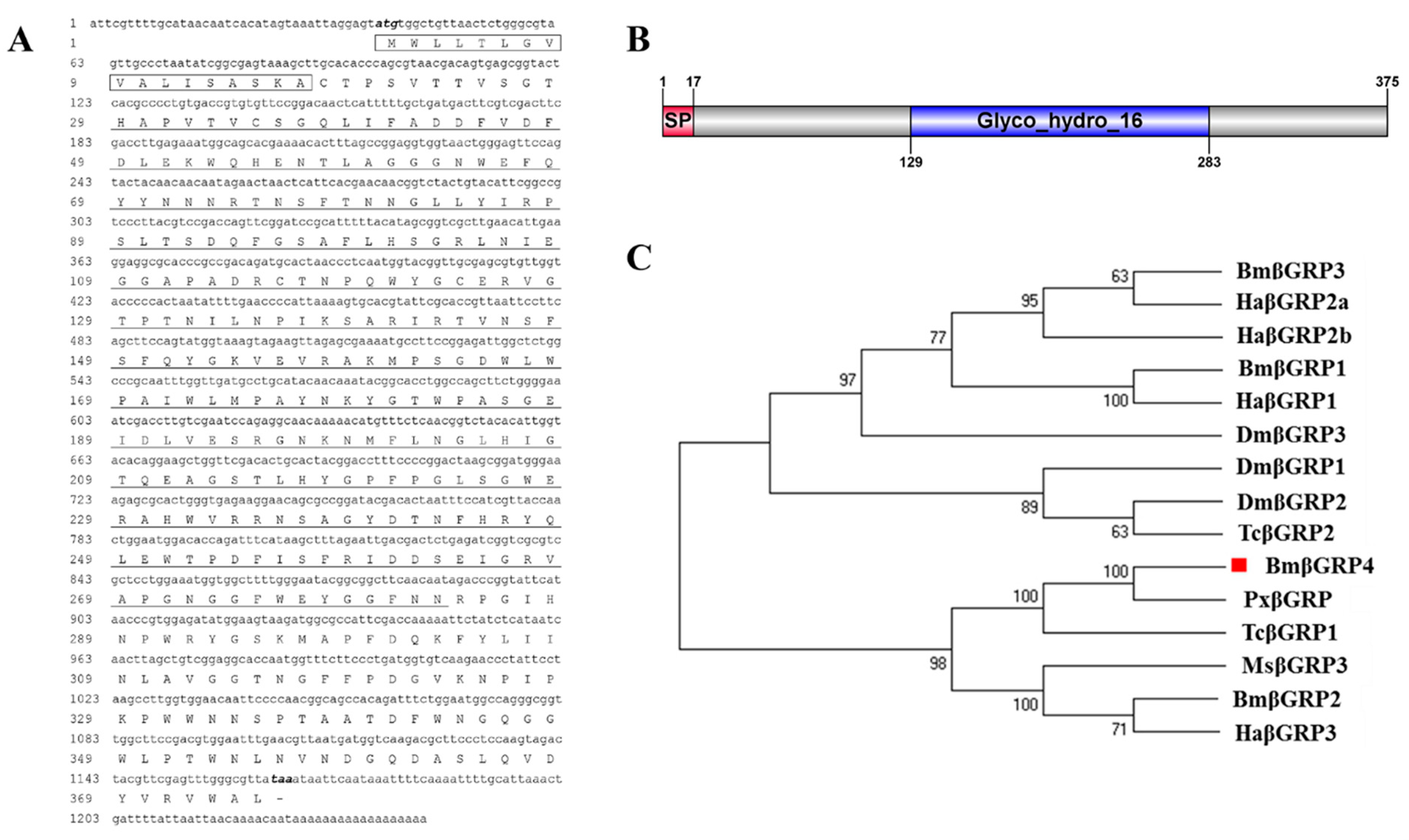{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silkworm Larvae, Cell Line and Virus Preparation
2.2. Identification and Phylogenetic Analysis of BmGRP4
2.3. RNA Isolation, cDNA Synthesis and RT-qPCR Analysis
2.4. Prokaryotic Expression and Protein Purification
2.5. Antibody Preparation and Western Blot Analysis
2.6. RNAi by Injecting Small Interfering RNA in B. mori Larvae and BmNPV Infection
2.7. Overexpression of BmβGRP4 in BmN Cells and BmNPV Infection
2.8. Detection of Apoptosis-Related Gene Expression and Observation of Apoptosis Morphology
2.9. Expression Level Analysis of BmPTEN and BmIAP
3. Results
3.1. Identification of BmβGRP4 and Bioinformatics Analysis
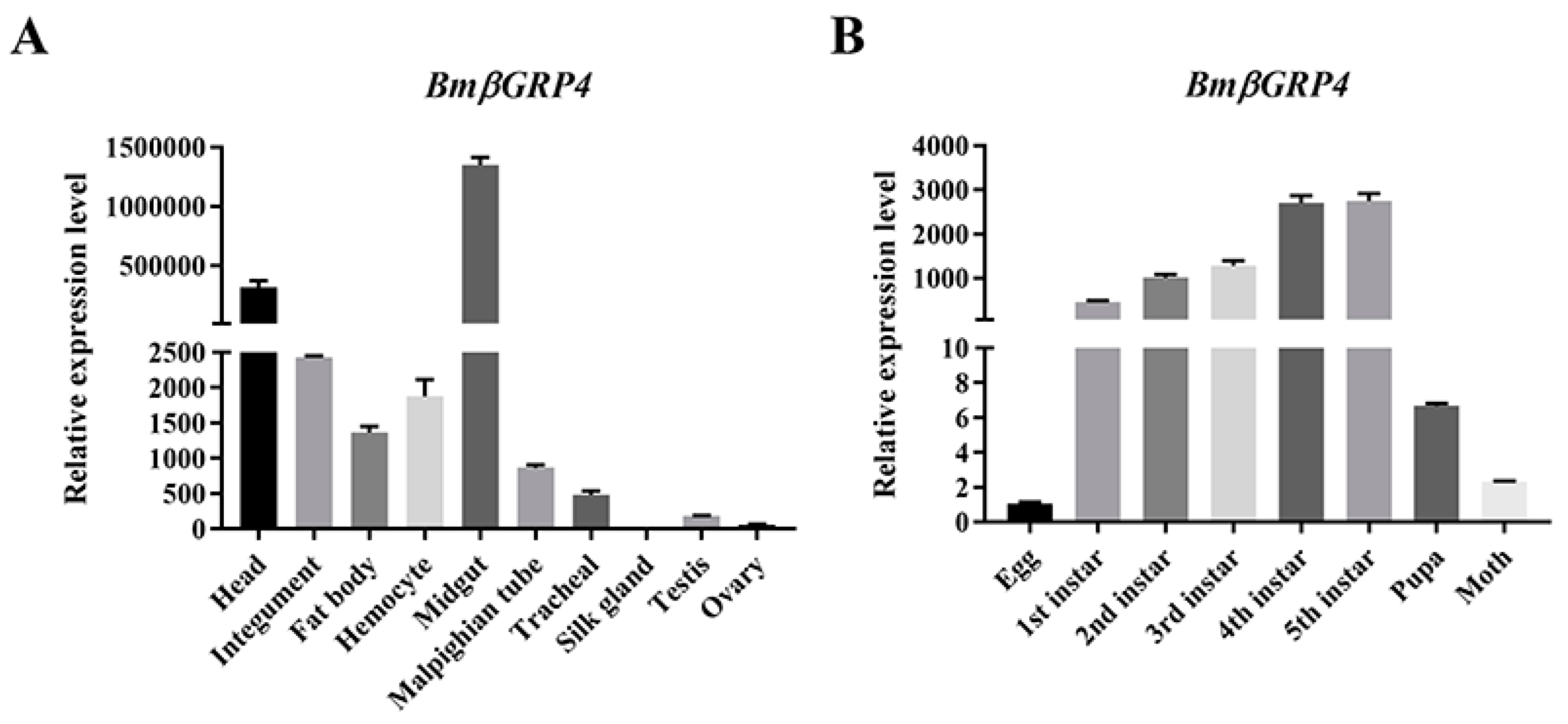
3.2. Tissue Distribution and Developmental Stage Expression of BmβGRP4
3.3. Recombinant BmβGRP4 Expression, Purification and Antiserum Preparation
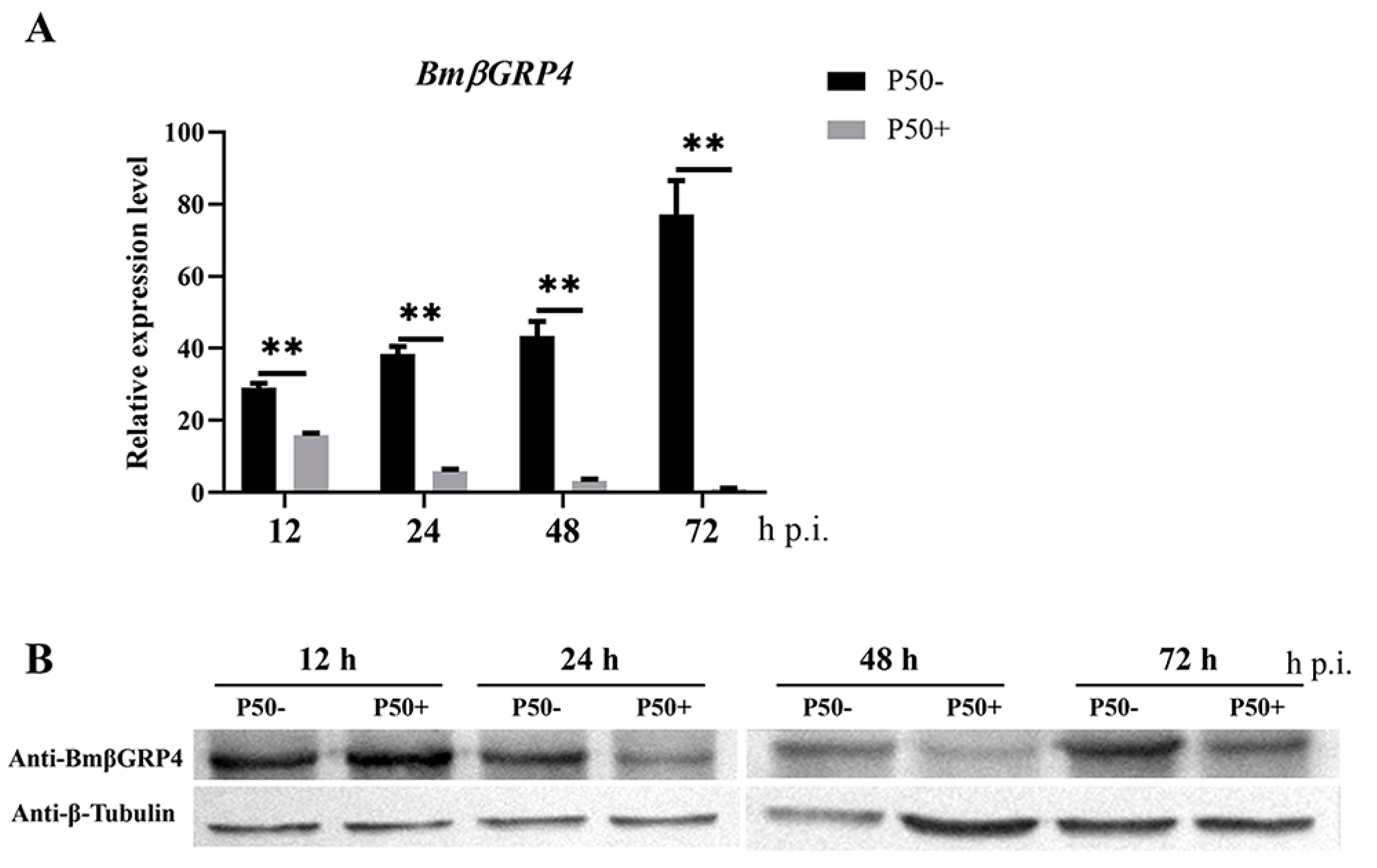
3.4. Expression Pattern Analysis of BmβGRP4 following BmNPV Infection
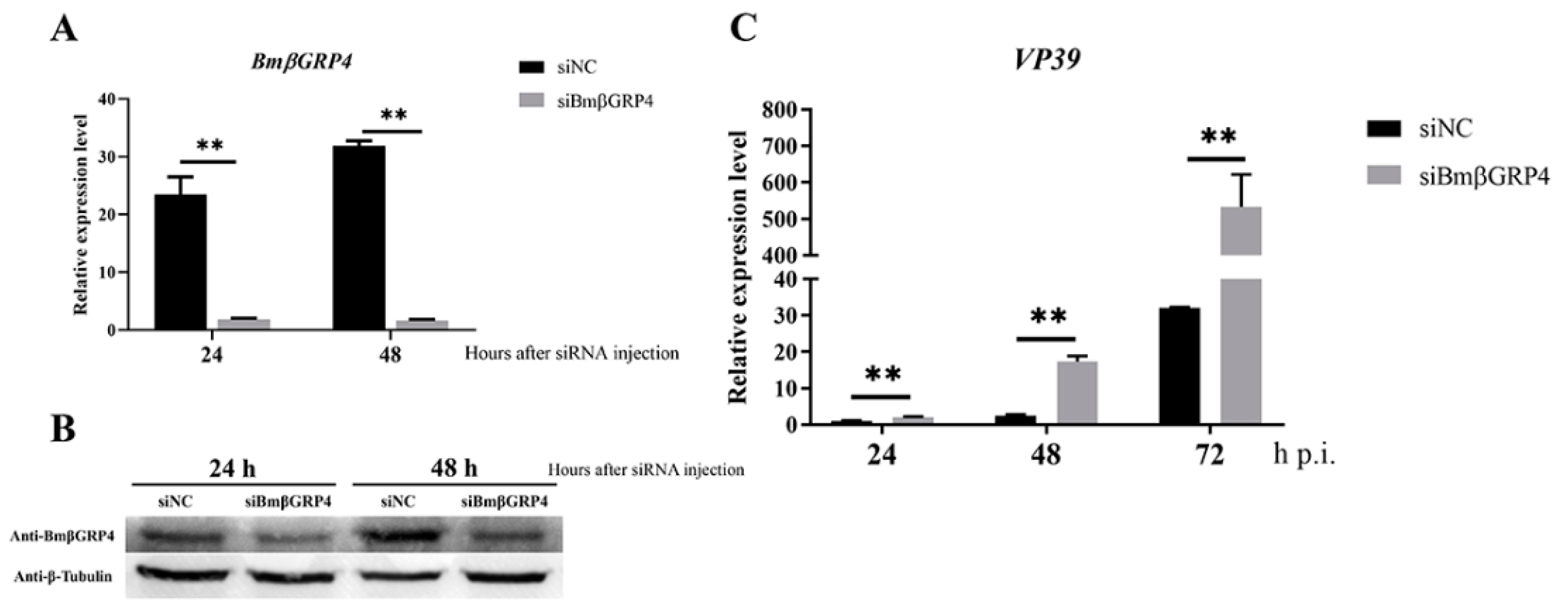
3.5. Knockdown of BmβGRP4 by RNAi Facilitated Virus Proliferation in B. mori Larvae
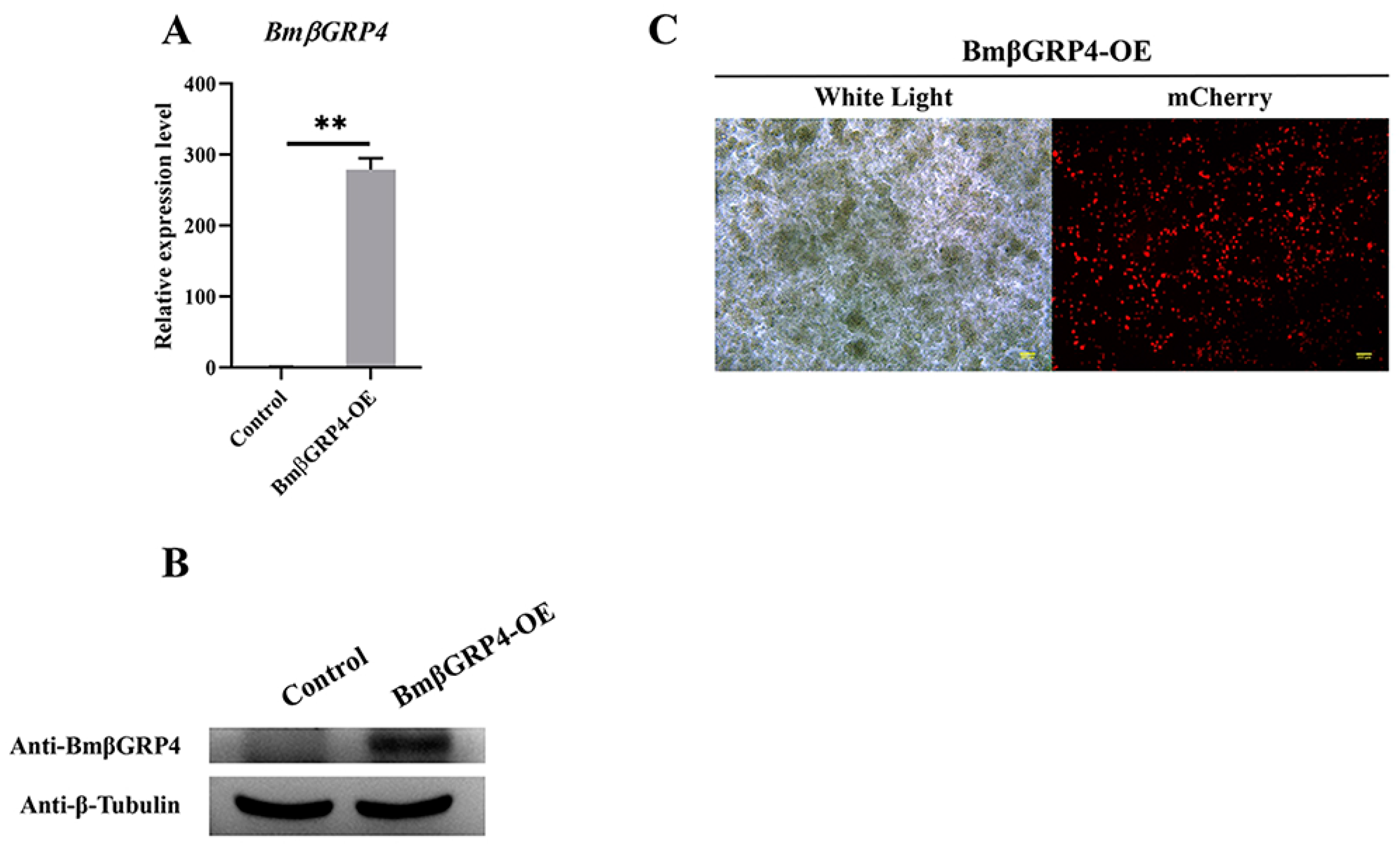
3.6. Overexpression of BmβGRP4 Suppressed Virus Proliferation in BmN Cells
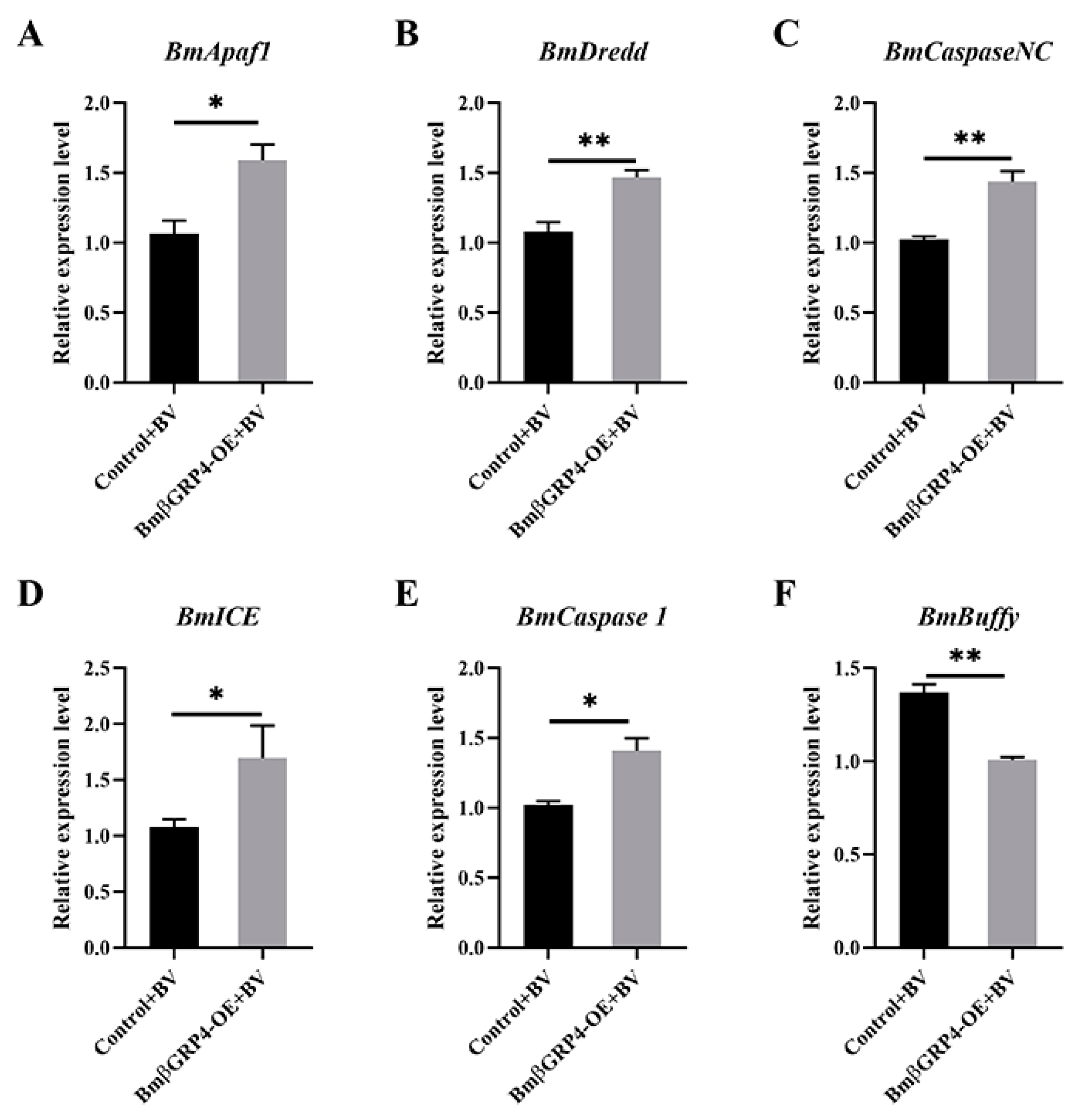
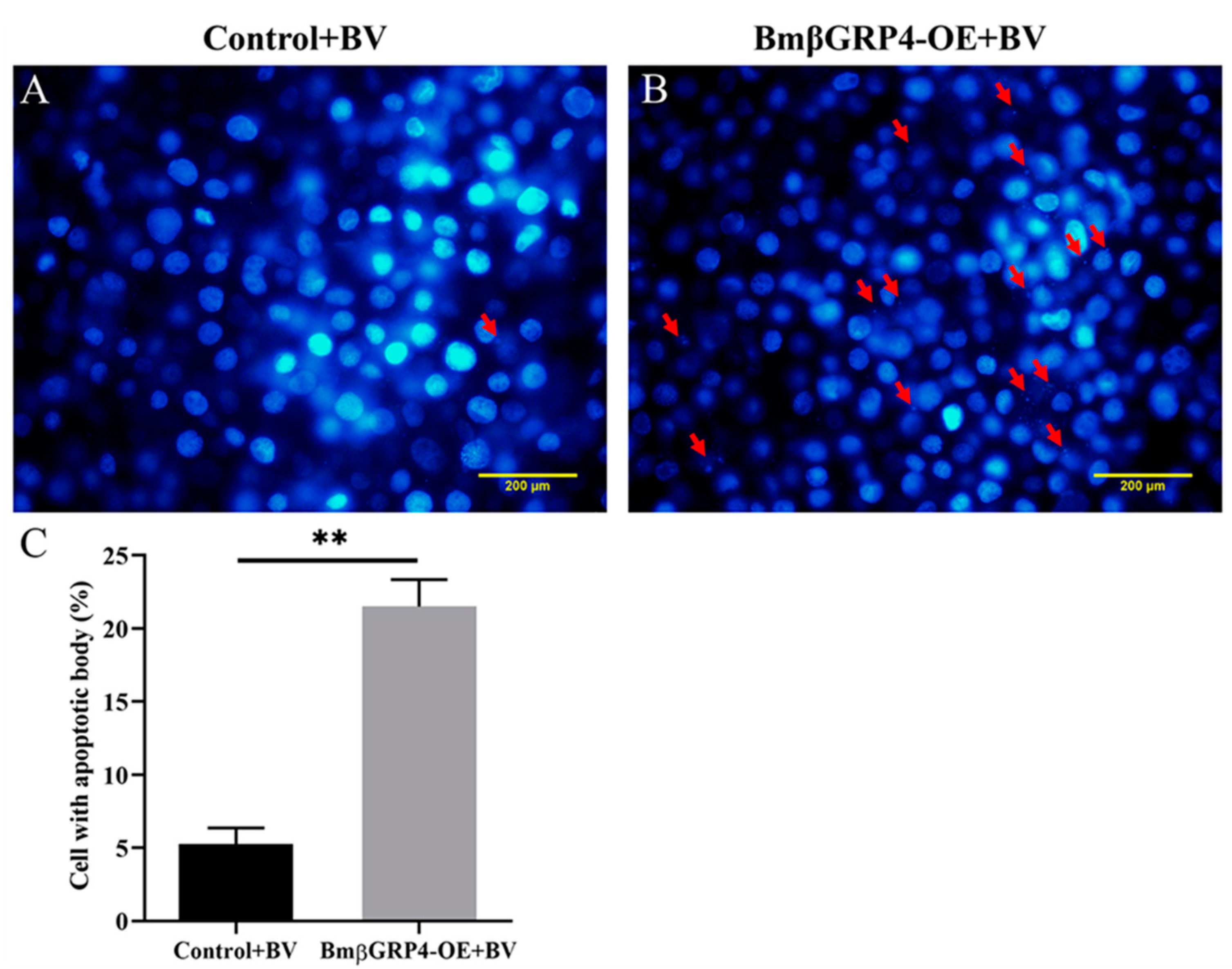
3.7. BmβGRP4 Promoted BmNPV Induced Apoptosis in BmN Cells
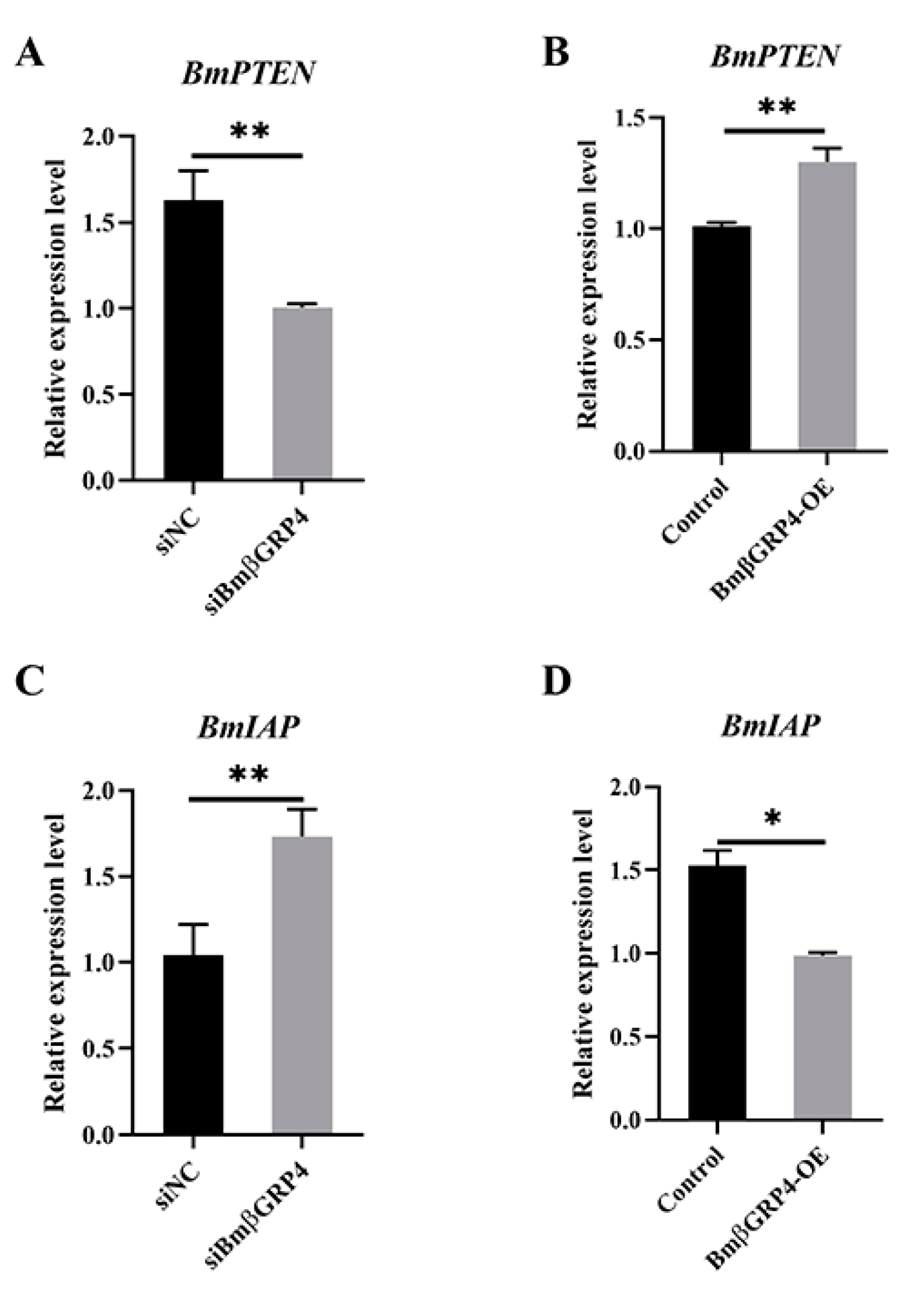
3.8. BmβGRP4 Positively Regulated BmPTEN and Negatively Regulated BmIAP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xia, Q.; Zhou, Z.; Lu, C.; Cheng, D.; Dai, F.; Li, B.; Zhao, P.; Zha, X.; Cheng, T.; Chai, C. A Draft Sequence for the Genome of the Domesticated Silkworm (Bombyx Mori). Science 2005, 306, 1937–1940. [Google Scholar]
- Kato, T.; Kajikawa, M.; Maenaka, K.; Park, E.Y. Silkworm Expression System as a Platform Technology in Life Science. Appl. Microbiol. Biotechnol. 2010, 85, 459–470. [Google Scholar] [CrossRef] [Green Version]
- The International Silkworm Genome Consortium. The Genome of a Lepidopteran Model Insect, the Silkworm Bombyx Mori. Insect Biochem. Mol. Biol. 2008, 38, 1036–1045. [Google Scholar] [CrossRef]
- Hu, X.; Shen, Y.; Zheng, Q.; Wang, G.; Wu, X.; Gong, C. Bm59 Is an Early Gene, but Is Unessential for the Propagation and Assembly of Bombyx Mori Nucleopolyhedrovirus. Mol. Genet. Genom. 2016, 291, 145–154. [Google Scholar] [CrossRef]
- Ponnuvel, K.M.; Nakazawa, H.; Furukawa, S.; Asaoka, A.; Ishibashi, J.; Tanaka, H.; Yamakawa, M. A Lipase Isolated from the Silkworm Bombyx Mori Shows Antiviral Activity against Nucleopolyhedrovirus. J. Virol. 2003, 77, 10725–10729. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Wang, X.-Y.; Du, C.; Gao, J.; Xu, J.-P. Expression Analysis of Several Antiviral Related Genes to Bmnpv in Different Resistant Strains of Silkworm, Bombyx Mori. J. Insect Sci. 2014, 14, 76. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Yu, H.-Z.; Geng, L.; Xu, J.-P.; Yu, D.; Zhang, S.-Z.; Ma, Y.; Fei, D.-Q. Comparative Transcriptome Analysis of Bombyx Mori (Lepidoptera) Larval Midgut Response to Bmnpv in Susceptible and near-Isogenic Resistant Strains. PLoS ONE 2016, 11, e0155341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.-Z.; Wang, X.-Y.; Xu, J.-P.; Ma, Y.; Zhang, S.-Z.; Yu, D.; Fei, D.-Q.; Muhammad, A. Itraq-Based Quantitative Proteomics Analysis of Molecular Mechanisms Associated with Bombyx Mori (Lepidoptera) Larval Midgut Response to Bmnpv in Susceptible and near-Isogenic Strains. J. Proteom. 2017, 165, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Qiao, N.; Zhang, W.; Cheng, R.L.; Zhang, X.Q.; Bao, Y.Y.; Xu, Y.P.; Gu, L.Z.; Han, J.D.; Zhang, C.X. Dynamic Interactions between Bombyx Mori Nucleopolyhedrovirus and Its Host Cells Revealed by Transcriptome Analysis. J. Virol. 2012, 86, 7345–7359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.L.; Liu, T.H.; Wang, W.; Pan, C.X.; Wu, Y.F.; Du, G.Y.; Chen, P.; Lu, C.; Pan, M.H. Bmreepa Is a Novel Gene That Facilitates Bmnpv Entry into Silkworm Cells. PLoS ONE 2015, 10, e0144575. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Antiviral Signaling through Pattern Recognition Receptors. J. Biochem. 2007, 141, 137. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Chen, J.C.; Lin, Y.C.; Kitikiew, S.; Li, H.F.; Bai, J.C.; Tseng, K.C.; Lin, B.W.; Liu, P.C.; Shi, Y.Z. Endogenous Molecules Induced by a Pathogen-Associated Molecular Pattern (Pamp) Elicit Innate Immunity in Shrimp. PLoS ONE 2014, 9, e115232. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Su, F.; Li, Q.; Zhang, J.; Li, Y.; Tang, T.; Hu, Q.; Yu, X.-Q. Pattern Recognition Receptors in Drosophila Immune Responses. Dev. Comp. Immunol. 2020, 102, 103468. [Google Scholar] [CrossRef]
- Gobert, V.; Gottar, M.; Matskevich, A.A.; Rutschmann, S.; Royet, J.; Belvin, M.; Hoffmann, J.A.; Ferrandon, D. Dual Activation of the Drosophila Toll Pathway by Two Pattern Recognition Receptors. Science 2003, 302, 2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; He, Y.; Cao, X.; Gunaratna, R.T.; Chen, Y.R.; Blissard, G.; Kanost, M.R.; Jiang, H. Phylogenetic Analysis and Expression Profiling of the Pattern Recognition Receptors: Insights into Molecular Recognition of Invading Pathogens in Manduca Sexta. Insect Biochem. Mol. Biol. 2015, 62, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjugam, M.; Vaseeharan, B.; Iswarya, A.; Amala, M.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Benelli, G. A Study on Β-Glucan Binding Protein (Β-Gbp) and Its Involvement in Phenoloxidase Cascade in Indian White Shrimp Fenneropenaeus Indicus. Mol. Immunol. 2017, 92, 1–11. [Google Scholar] [CrossRef]
- Jiang, H.; Ma, C.; Lu, Z.-Q.; Kanost, M.R. Beta-1,3-Glucan Recognition Protein-2 (Betagrp-2) from Manduca Sexta; an Acute-Phase Protein That Binds Beta-1,3-Glucan and Lipoteichoic Acid to Aggregate Fungi and Bacteria and Stimulate Prophenoloxidase Activation. Insect Biochem. Mol. Biol. 2004, 34, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H. Peptidoglycan Recognition Proteins: On and Off Switches for Innate Immunity. Immunol. Rev. 2004, 198, 83–96. [Google Scholar] [CrossRef]
- Krem, M.M.; Cera, E.D. Evolution of Enzyme Cascades from Embryonic Development to Blood Coagulation. Trends Biochem. Sci. 2002, 27, 67–74. [Google Scholar] [CrossRef]
- Nappi, A.J.; Vass, E. Cytotoxic Reactions Associated with Insect Immunity. Adv. Exp. Med. Biol. 2001, 484, 329. [Google Scholar]
- Ochiai, M.; Ashida, M. Purification of a Beta-1,3-Glucan Recognition Protein in the Prophenoloxidase Activating System from Hemolymph of the Silkworm, Bombyx Mori. J. Biol. Chem. 1988, 263, 12056. [Google Scholar] [CrossRef]
- Huang, W.; Xu, X.; Freed, S.; Zheng, Z.; Shuang, W.; Ren, S.; Jin, F. Molecular Cloning and Characterization of a Β-1,3-Glucan Recognition Protein from Plutella Xylostella (L.). New Biotechnol. 2015, 32, 290. [Google Scholar] [CrossRef]
- Thompson, M.R.; Kaminski, J.J.; Kurt-Jones, E.A.; Fitzgerald, K.A. Pattern Recognition Receptors and the Innate Immune Response to Viral Infection. Viruses 2011, 3, 920–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. Innate Immune Recognition of Viral Infection. Nat. Immunol. 2006, 7, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Heiberg, I.L.; Winther, T.N.; Paludan, S.R.; Hogh, B. Pattern Recognition Receptor Responses in Children with Chronic Hepatitis B Virus Infection. J. Clin. Virol. 2012, 54, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.W.; McGraw, E.A.; Ammar, E.D.; Dietzgen, R.G.; Hogenhout, S.A. Drosophila Melanogaster Mounts a Unique Immune Response to the Rhabdovirus Sigma Virus. Appl. Environ. Microbiol. 2008, 74, 3251–3256. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Deng, X.-Y.; Qian, H.-Y.; Qin, G.-X.; Hou, C.-X.; Guo, X.-J. Cloning and Expression Analysis of a Peptidoglycan Recognition Protein in Silkworm Related to Virus Infection. Gene 2014, 552, 24–31. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.-Z.; Zhu, L.-B.; You, L.-L.; Wang, J.; Cao, H.-H.; Liu, Y.-X.; Toufeeq, S.; Wang, Y.-L.; Kong, X.; Xu, J.-P. A Novel Digestive Proteinase Lipase Member H-a in Bombyx Mori Contributes to Digestive Juice Antiviral Activity against B. Mori Nucleopolyhedrovirus. Insects 2020, 11, 154. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Liu, W.; Guo, H.; Dang, Y.; Cheng, T.; Yang, W.; Sun, Q.; Wang, B.; Wang, Y.; Xie, E.; et al. Distinct Functions of Bombyx Mori Peptidoglycan Recognition Protein 2 in Immune Responses to Bacteria and Viruses. Front. Immunol. 2019, 10, 776. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Kang, T.-T.; Bao, X.-Y.; Dong, Z.-Q.; Zhu, Y.; Xiao, W.-F.; Pan, M.-H.; Lu, C. Evolutionary and Functional Analyses of the Interaction between the Bombyx Mori Inhibitor of Apoptosis (Iap) and Nucleopolyhedrovirus Iaps. Insect Sci. 2020, 27, 463–474. [Google Scholar] [CrossRef]
- Everett, H.; McFadden, G. Apoptosis: An Innate Immune Response to Virus Infection. Trends Microbiol. 1999, 7, 160–165. [Google Scholar] [CrossRef]
- Clarke, T.E.; Clem, R.J. Insect Defenses against Virus Infection: The Role of Apoptosis. Int. Rev. Immunol. 2003, 22, 401–424. [Google Scholar] [CrossRef]
- Engelhard, E.K.; Volkman, L.E. Developmental Resistance in Fourth Instar Trichoplusia Ni Orally Inoculated with Autographa Californica M Nuclear Polyhedrosis Virus. Virology 1995, 209, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Kanost, M.R. A Β1,3-Glucan Recognition Protein from an Insect, Manduca Sexta, Agglutinates Microorganisms and Activates the Phenoloxidase Cascade. J. Biol. Chem. 2000, 275, 7505–7514. [Google Scholar] [CrossRef] [Green Version]
- Fabrick, J.A.; Baker, J.E.; Kanost, M.R. Cdna Cloning, Purification, Properties, and Function of a Beta-1,3-Glucan Recognition Protein from a Pyralid Moth, Plodia Interpunctella. Insect Biochem. Mol. Biol. 2003, 33, 579–594. [Google Scholar] [CrossRef]
- Wu, T.; Zhao, Y.; Wang, Z.; Song, Q.; Wang, Z.; Xu, Q.; Wang, Y.; Wang, L.; Zhang, Y.; Feng, C. Β-1,3-Glucan Recognition Protein 3 Activates the Prophenoloxidase System in Response to Bacterial Infection in Ostrinia Furnacalis Guenée. Dev. Comp. Immunol. 2017, 79, 31. [Google Scholar] [CrossRef]
- Li, G.; Zhou, Q.; Qiu, L.; Yao, Q.; Chen, K.; Tang, Q.; Hu, Z. Serine Protease Bm-Sp142 Was Differentially Expressed in Resistant and Susceptible Bombyx Mori Strains, Involving in the Defence Response to Viral Infection. PLoS ONE 2017, 12, e0175518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.-H.; Zhang, S.-Z.; Zhu, L.-B.; Wang, J.; Liu, Y.-X.; Wang, Y.-L.; Kong, X.; You, L.-L.; Toufeeq, S.; Liu, S.-H.; et al. The Digestive Proteinase Trypsin, Alkaline a Contributes to Anti-Bmnpv Activity in Silkworm (Bombyx Mori). Dev. Comp. Immunol. 2021, 119, 104035. [Google Scholar] [CrossRef] [PubMed]
- Marques, J.T.; Imler, J.-L. The Diversity of Insect Antiviral Immunity: Insights from Viruses. Curr. Opin. Microbiol. 2016, 32, 71–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhu, L.-B.; Ma, Y.; Liu, Y.-X.; Cao, H.-H.; Wang, Y.-L.; Kong, X.; Huang, Z.-H.; Zhu, H.-D.; Wang, Y.-X.; et al. Bombyx mori β-1,3-Glucan Recognition Protein 4 (BmβGRP4) Could Inhibit the Proliferation of B. mori Nucleopolyhedrovirus through Promoting Apoptosis. Insects 2021, 12, 743. https://doi.org/10.3390/insects12080743
Wang J, Zhu L-B, Ma Y, Liu Y-X, Cao H-H, Wang Y-L, Kong X, Huang Z-H, Zhu H-D, Wang Y-X, et al. Bombyx mori β-1,3-Glucan Recognition Protein 4 (BmβGRP4) Could Inhibit the Proliferation of B. mori Nucleopolyhedrovirus through Promoting Apoptosis. Insects. 2021; 12(8):743. https://doi.org/10.3390/insects12080743
Chicago/Turabian StyleWang, Jie, Lin-Bao Zhu, Yan Ma, Ying-Xue Liu, Hui-Hua Cao, Yu-Ling Wang, Xue Kong, Zhi-Hao Huang, Han-Dan Zhu, Yan-Xiang Wang, and et al. 2021. "Bombyx mori β-1,3-Glucan Recognition Protein 4 (BmβGRP4) Could Inhibit the Proliferation of B. mori Nucleopolyhedrovirus through Promoting Apoptosis" Insects 12, no. 8: 743. https://doi.org/10.3390/insects12080743
APA StyleWang, J., Zhu, L.-B., Ma, Y., Liu, Y.-X., Cao, H.-H., Wang, Y.-L., Kong, X., Huang, Z.-H., Zhu, H.-D., Wang, Y.-X., Liu, S.-H., & Xu, J.-P. (2021). Bombyx mori β-1,3-Glucan Recognition Protein 4 (BmβGRP4) Could Inhibit the Proliferation of B. mori Nucleopolyhedrovirus through Promoting Apoptosis. Insects, 12(8), 743. https://doi.org/10.3390/insects12080743