
Cuticular Hydrocarbon Profile of Parasitic Beetles, Aethina tumida (Coleoptera: Nitidulidae)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. GC-MS Analyses
2.3. Statistical Analyses
3. Results
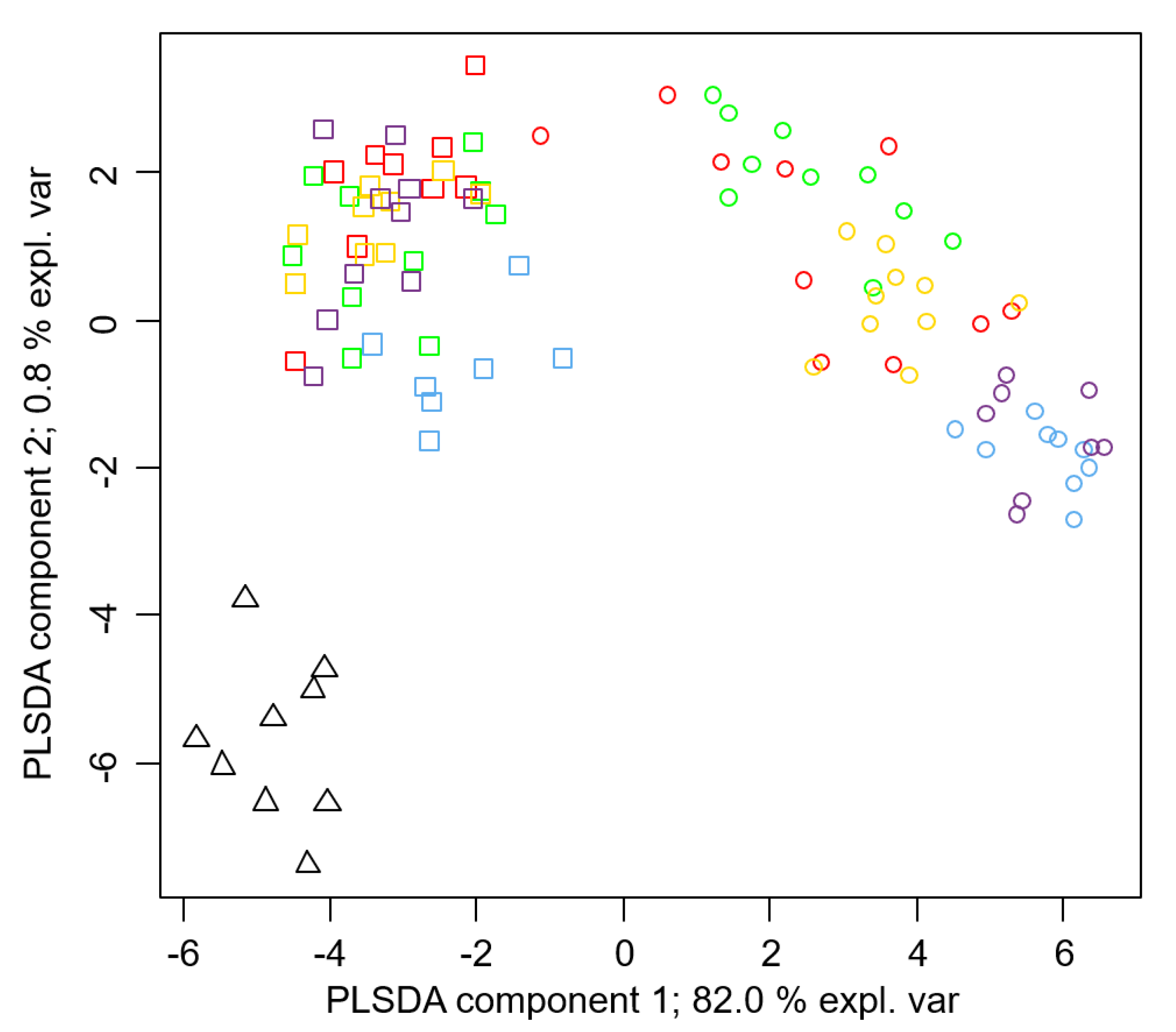
3.1. Colony Membership Allocation
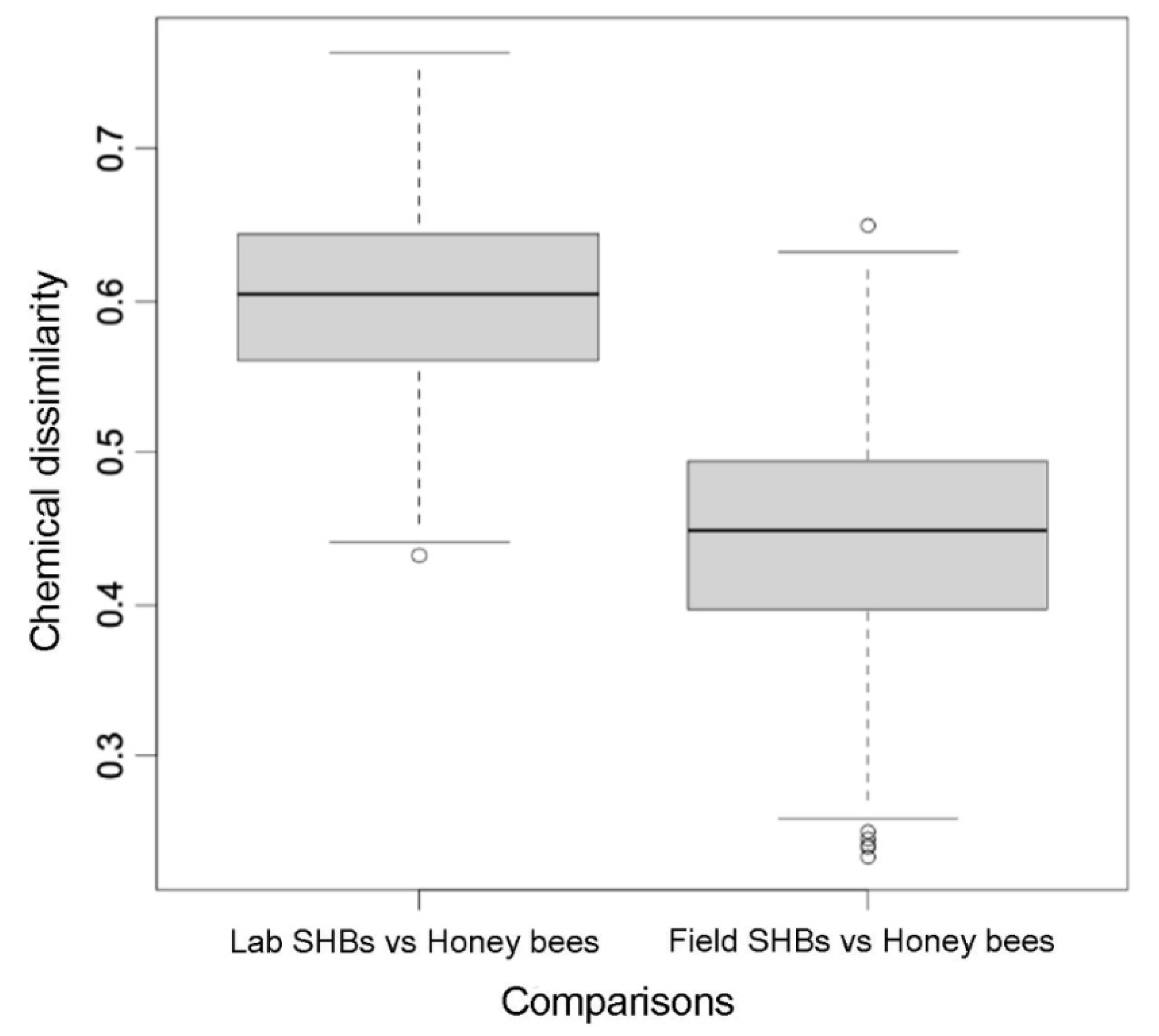
3.2. Qualitative Chemical Analysis
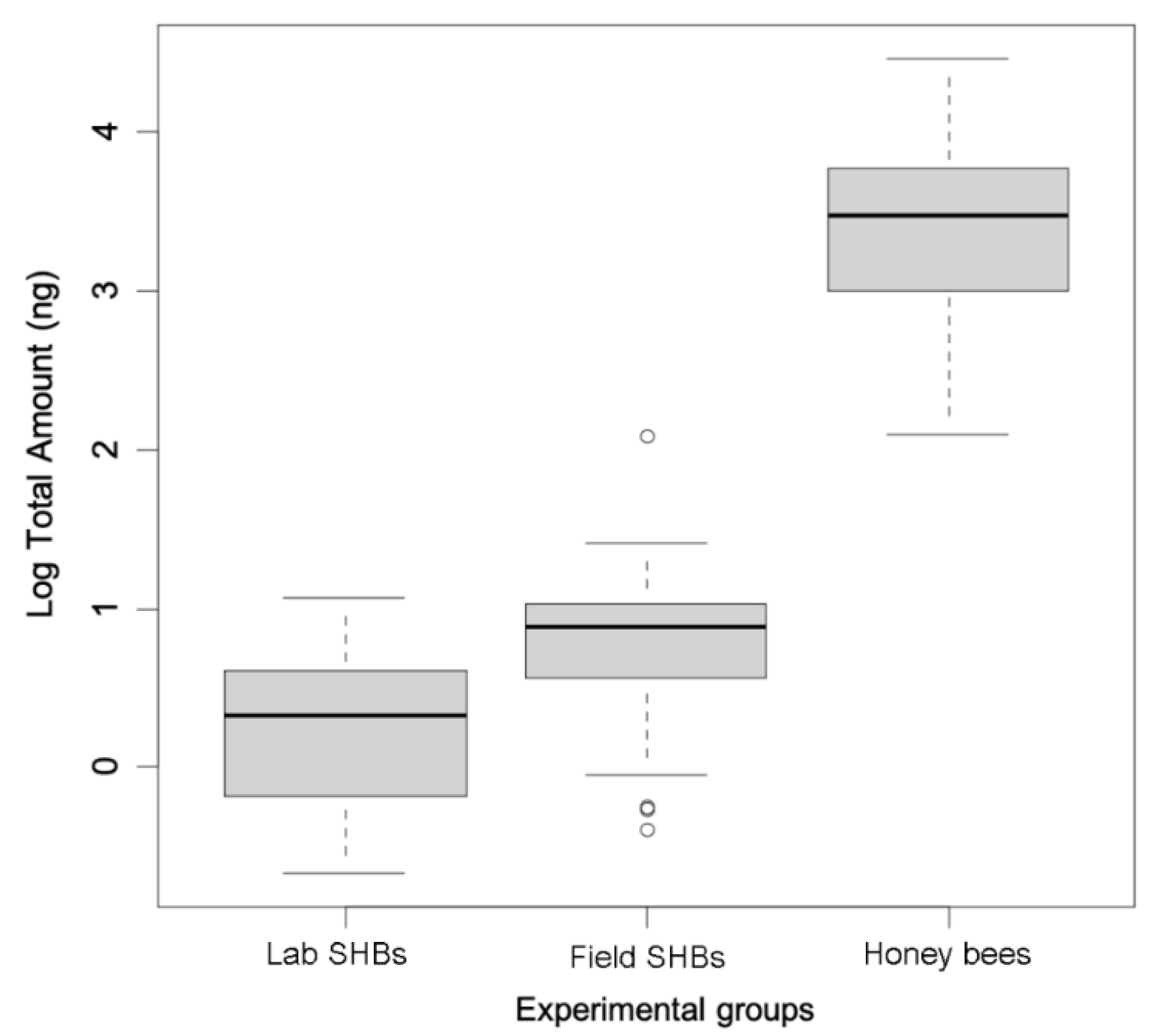
3.3. Quantitative Chemical Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breed, M.D.; Cook, C.N.; McCreery, H.F.; Rodriguez, M. Nestmate recognition in eusocial insects: The honeybee as a model system. In Social Recognition in Invertebrates: The Knowns and the Unknowns; Aquiloni, L., Tricarico, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 147–164. ISBN 9783319175997. [Google Scholar]
- d’Ettorre, P.; Lenoir, A. Nestmate recognition. In Ant Ecology; Lach, L., Parr, C., Abbott, K., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 194–209. ISBN 9780191720192. [Google Scholar]
- Leonhardt, S.D.; Menzel, F.; Nehring, V.; Schmitt, T. Ecology and evolution of communication in social insects. Cell 2016, 164, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Cervo, R.; Cini, A.; Turillazzi, S. Visual recognition in social wasps. In Social Recognition in Invertebrates: The Knowns and the Unknowns; Aquiloni, L., Tricarico, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 125–145. ISBN 9783319175997. [Google Scholar]
- Cini, A.; Sumner, S.; Cervo, R. Inquiline social parasites as tools to unlock the secrets of insect sociality. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180193. [Google Scholar] [CrossRef] [Green Version]
- Schönrogge, K.; Barbero, F.; Casacci, L.P.; Settele, J.; Thomas, J.A. Acoustic communication within ant societies and its mimicry by mutualistic and socially parasitic myrmecophiles. Anim. Behav. 2017, 134, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Blomquist, G.J.; Bagnères, A.G. Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology; Cambridge University Press: Cambridge, UK, 2010; ISBN 9780511711909. [Google Scholar]
- Bruschini, C.; Cervo, R.; Stefano, T. Pheromones in Social Wasps. Vitam Horm. 2010, 83, 447–492. [Google Scholar] [CrossRef]
- Lenoir, A.; D’Ettorre, P.; Errard, C.; Hefetz, A. Chemical ecology and social parasitism in ants. Annu. Rev. Entomol. 2001, 46, 573–599. [Google Scholar] [CrossRef] [PubMed]
- Cini, A.; Bruschini, C.; Signorotti, L.; Pontieri, L.; Turillazzi, S.; Cervo, R. The chemical basis of host nest detection and chemical integration in a cuckoo paper wasp. J. Exp. Biol. 2011, 214, 3698–3703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dettner, K.; Liepert, C. Chemical mimicry and camouflage. Annu. Rev. Entomol. 1994, 39, 129–154. [Google Scholar] [CrossRef]
- Uboni, A.; Bagnères, A.G.; Christidès, J.P.; Lorenzi, M.C. Cleptoparasites, social parasites and a common host: Chemical insignificance for visiting host nests, chemical mimicry for living in. J. Insect Physiol. 2012, 58, 1259–1264. [Google Scholar] [CrossRef]
- Vander Meer, R.K.; Wojcik, D.P. Chemical mimicry in the myrmecophilous beetle Myrmecaphodius excavaticollis. Science 1982, 218, 806–808. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, M.C.; d’Ettorre, P. Nestmate recognition in social insects: What does it mean to be chemically insignificant? Front. Ecol. Evol. 2020, 7, 488. [Google Scholar] [CrossRef] [Green Version]
- Cappa, F.; Bruschini, C.; Cipollini, M.; Pieraccini, G.; Cervo, R. Sensing the intruder: A quantitative threshold for recognition cues perception in honeybees. Naturwissenschaften 2014, 101, 149–152. [Google Scholar] [CrossRef] [Green Version]
- Cini, A.; Gioli, L.; Cervo, R. A quantitative threshold for nest-mate recognition in a paper social wasp. Biol. Lett. 2009, 5, 459–461. [Google Scholar] [CrossRef] [Green Version]
- Ichinose, K.; Lenoir, A. Hydrocarbons detection levels in ants. Insectes Soc. 2010, 57, 453–455. [Google Scholar] [CrossRef]
- Akino, T.; Knapp, J.J.; Thomas, J.A.; Elmes, G.W. Chemical mimicry and host specificity in the butterfly Maculinea rebeli, a social parasite of Myrmica ant colonies. Proc. R. Soc. B Biol. Sci. 1999, 266, 1419–1426. [Google Scholar] [CrossRef] [Green Version]
- Lorenzi, M.C.; Cervo, R.; Zacchi, F.; Turillazzi, S.; Bagnères, A.G. Dynamics of chemical mimicry in the social parasite wasp Polistes semenowi (Hymenoptera: Vespidae). Parasitology 2004, 129, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Young, H.P.; Schal, C. Cuticular hydrocarbon synthesis in relation to feeding and developmental stage in nymphs of Blattella germanica (Dictyoptera: Blattellidae). Ann. Entomol. Soc. Am. 1997, 90, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Bos, N.; Grinsted, L.; Holman, L. Wax On, Wax off: Nest soil facilitates indirect transfer of recognition cues between ant nestmates. PLoS ONE 2011, 6, e19435. [Google Scholar] [CrossRef] [PubMed]
- Couvillon, M.J.; Caple, J.P.; Endsor, S.L.; Kärcher, M.; Russell, T.E.; Storey, D.E.; Ratnieks, F.L.W. Nest-mate recognition template of guard honeybees (Apis mellifera) is modified by wax comb transfer. Biol. Lett. 2007, 3, 228–230. [Google Scholar] [CrossRef]
- D’Ettorre, P.; Wenseleers, T.; Dawson, J.; Hutchinson, S.; Boswell, T.; Ratnieks, F.L.W. Wax combs mediate nestmate recognition by guard honeybees. Anim. Behav. 2006, 71, 773–779. [Google Scholar] [CrossRef] [Green Version]
- Dahbi, A.; Cerdá, X.; Hefetz, A.; Lenoir, A. Adult transport in the ant cataglyphis iberica: A means to maintain a uniform colonial odour in a species with multiple nests. Physiol. Entomol. 1997, 22, 13–19. [Google Scholar] [CrossRef]
- Soroker, V.; Vienne, C.; Hefetz, A. Hydrocarbon dynamics within and between nestmates in Cataglyphis niger (Hymenoptera: Formicidae). J. Chem. Ecol. 1995, 21, 365–378. [Google Scholar] [CrossRef]
- Cappa, F.; Beani, L.; Cervo, R. The importance of being yellow: Visual over chemical cues in gender recognition in a social wasp. Behav. Ecol. 2016, 27, 1182–1189. [Google Scholar] [CrossRef] [Green Version]
- Cappa, F.; Petrocelli, I.; Dani, F.R.; Dapporto, L.; Giovannini, M.; Silva-Castellari, J.; Turillazzi, S.; Cervo, R. Natural biocide disrupts nestmate recognition in honeybees. Sci. Rep. 2019, 9, 3171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonnell, C.M.; Alaux, C.; Parrinello, H.; Desvignes, J.P.; Crauser, D.; Durbesson, E.; Beslay, D.; Le Conte, Y. Ecto- and endoparasite induce similar chemical and brain neurogenomic responses in the honey bee (Apis mellifera). BMC Ecol. 2013, 13, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blomquist, G.J.; Jackson, L.L. Incorporation of labelled dietary n-alkanes into cuticular lipids of the grasshopper Melanoplus sanguinipes. J. Insect Physiol. 1973, 19, 1639–1647. [Google Scholar] [CrossRef]
- Liang, D.; Silverman, J. “You are what you eat”: Diet modifies cuticular hydrocarbons and nestmate recognition in the Argentine ant, Linepithema humile. Naturwissenschaften 2000, 87, 412–416. [Google Scholar] [CrossRef]
- Lundie, A.E. The small hive beetle, Aethina tumida. Sci. Bull. Dep. Agric. For. Union S. Afr. 1940, 220, 30. [Google Scholar]
- Al Toufailia, H.; Alves, D.A.; Bená, D.D.C.; Bento, J.M.S.; Iwanicki, N.S.A.; Cline, A.R.; Ellis, J.D.; Ratnieks, F.L.W. First record of small hive beetle, Aethina tumida Murray, in South America. J. Apic. Res. 2017, 56, 76–80. [Google Scholar] [CrossRef]
- Liu, Y.; Han, W.; Gao, J.; Su, S.; Beaurepaire, A.; Yañez, O.; Neumann, P. Out of Africa: Novel source of small hive beetles infesting Eastern and Western honey bee colonies in China. J. Apic. Res. 2020, 60, 108–110. [Google Scholar] [CrossRef]
- Neumann, P.; Pettis, J.S.; Schäfer, M.O. Quo vadis Aethina tumida? Biology and control of small hive beetles. Apidologie 2016, 47, 427–466. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P. Small hive beetle in Italy: What can we expect in the future? In Small Hive Beetle—A Growing Problem in the 21st Century; Carreck, N.L., Ed.; International Bee Research Association: Bristol, UK, 2017; pp. 33–40. [Google Scholar]
- Gonthier, J.; Papach, A.; Straub, L.; Campbell, J.W.; Williams, G.R.; Neumann, P. Bees and flowers: How to feed an invasive beetle species. Ecol. Evol. 2019, 9, 6422–6432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiewok, S.; Neumann, P. Infestation of commercial bumblebee (Bombus impatiens) field colonies by small hive beetles (Aethina tumida). Ecol. Entomol. 2006, 31, 623–628. [Google Scholar] [CrossRef]
- Neumann, P.; Naef, J.; Crailsheim, K.; Crewe, R.M.; Pirk, C.W.W. Hit-and-run trophallaxis of small hive beetles. Ecol. Evol. 2015, 5, 5478–5486. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.; Hoffmann, D.; Duncan, M.; Spooner-Hart, R.; Pettis, J.S. Long-range dispersal of small hive beetles. J. Apic. Res. 2012, 51, 214–215. [Google Scholar] [CrossRef]
- Neumann, P.; Elzen, P.J. The biology of the small hive beetle (Aethina tumida, Coleoptera: Nitidulidae): Gaps in our knowledge of an invasive species. Apidologie 2004, 35, 229–247. [Google Scholar] [CrossRef] [Green Version]
- Spiewok, S.; Pettis, J.S.; Duncan, M.; Spooner-Hart, R.; Westervelt, D.; Neumann, P. Small hive beetle, Aethina tumida, populations I: Infestation levels of honeybee colonies, apiaries and regions. Apidologie 2007, 38, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Spiewok, S.; Duncan, M.; Spooner-Hart, R.; Pettis, J.S.; Neumann, P. Small hive beetle, Aethina tumida, populations II: Dispersal of small hive beetles. Apidologie 2008, 39, 683–693. [Google Scholar] [CrossRef]
- Schmolke, M.D. A Study of Aethina Tumida: The Small Hive Beetle; University of Rhodesia: Harare, Zimbabwe, 1974; Project Report. [Google Scholar]
- Neumann, P.; Evans, J.D.; Pettis, J.S.; Pirk, C.W.W.; Schäfer, M.O.; Tanner, G.; Ellis, J.D. Standard methods for small hive beetle research. J. Apic. Res. 2013, 52, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Papach, A.; Gonthier, J.; Williams, G.R.; Neumann, P. Project report sex ratio of small hive beetles: The role of pupation and adult longevity. Insects 2019, 10, 133. [Google Scholar] [CrossRef] [Green Version]
- Aitchison, J. The Statistical Analysis of Compositional Data; Chapman and Hall: London, UK, 1986; ISBN 9780412280603. [Google Scholar]
- Neumann, P.; Hoffmann, D.; Duncan, M.; Spooner-Hart, R. High and rapid infestation of isolated commercial honey bee colonies with small hive beetles in Australia. J. Apic. Res. 2010, 49, 343–344. [Google Scholar] [CrossRef]
- Neumann, P.; Pirk, C.W.W.; Hepburn, H.R.; Solbrig, A.J.; Ratnieks, F.L.W.; Elzen, P.J.; Baxter, J.R. Social encapsulation of beetle parasites by Cape honeybee colonies (Apis mellifera capensis Esch.). Naturwissenschaften 2001, 88, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, V.; Pogoda, W.; Verhoff, M.A.; Toennes, S.W.; Amendt, J. Estimating the age of the adult stages of the blow flies Lucilia sericata and Calliphora vicina (Diptera: Calliphoridae) by means of the cuticular hydrocarbon n-pentacosane. Sci. Justice 2017, 57, 361–365. [Google Scholar] [CrossRef]
- Butterworth, N.J.; Drijfhout, F.P.; Byrne, P.G.; Keller, P.A.; Wallman, J.F. Major transitions in cuticular hydrocarbon expression coincide with sexual maturity in a blowfly (Diptera: Calliphoridae). J. Chem. Ecol. 2020, 46, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Vaníčková, L.; Svatoš, A.; Kroiss, J.; Kaltenpoth, M.; Do Nascimento, R.R.; Hoskovec, M.; Břízová, R.; Kalinová, B. Cuticular Hydrocarbons of the South American Fruit Fly Anastrepha fraterculus: Variability with sex and age. J. Chem. Ecol. 2012, 38, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Dapporto, L.; Sledge, F.M.; Turillazzi, S. Dynamics of cuticular chemical profiles of Polistes dominulus workers in orphaned nests (Hymenoptera, Vespidae). J. Insect Physiol. 2005, 51, 969–973. [Google Scholar] [CrossRef]
- Fedina, T.Y.; Kuo, T.-H.; Dreisewerd, K.; Dierick, H.A.; Yew, J.Y.; Pletcher, S.D. Dietary effects on cuticular hydrocarbons and sexual attractiveness in Drosophila. PLoS ONE 2012, 7, e49799. [Google Scholar] [CrossRef] [Green Version]
- Florane, C.B.; Bland, J.M.; Husseneder, C.; Raina, A.K. Diet-mediated inter-colonial aggression in the formosan subterranean termite Coptotermes formosanus. J. Chem. Ecol. 2004, 30, 2559–2574. [Google Scholar] [CrossRef]
- Otte, T.; Hilker, M.; Geiselhardt, S. The effect of dietary fatty acids on the cuticular hydrocarbon phenotype of an herbivorous insect and consequences for mate recognition. J. Chem. Ecol. 2014, 41, 32–43. [Google Scholar] [CrossRef]
- Otte, T.; Hilker, M.; Geiselhardt, S. Phenotypic plasticity of cuticular hydrocarbon profiles in insects. J. Chem. Ecol. 2018, 44, 235–247. [Google Scholar] [CrossRef]
- Breed, M.D.; Leger, E.A.; Pearce, A.N.; Wang, Y.J. Comb wax effects on the ontogeny of honey bee nestmate recognition. Anim. Behav. 1998, 55, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Dani, F.R.; Jones, G.R.; Corsi, S.; Beard, R.; Pradella, D.; Turillazzi, S. Nestmate recognition cues in the honey bee: Differential importance of cuticular alkanes and alkenes. Chem. Senses 2005, 30, 477–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kather, R.; Drijfhout, F.P.; Shemilt, S.; Martin, S.J. Evidence for passive chemical camouflage in the parasitic mite Varroa destructor. J. Chem. Ecol. 2015, 41, 178–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, J.D.; Delaplane, K.S.; Hood, W.M.; Elzen, P.J. Small hive beetle (Aethina tumida Murray) weight, gross biometry, and sex proportion at three locations in the south-eastern United States. Am. Bee J. 2002, 142, 520–522. [Google Scholar]
- Dani, F.R.; Jones, G.R.; Destri, S.; Spencer, S.H.; Turillazzi, S. Deciphering the recognition signature within the cuticular chemical profile of paper wasps. Anim. Behav. 2001, 62, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Kather, R.; Martin, S.J. Evolution of cuticular hydrocarbons in the hymenoptera: A meta-analysis. J. Chem. Ecol. 2015, 41, 871–883. [Google Scholar] [CrossRef] [Green Version]
- Brückner, A.; Heethoff, M.; Blühgen, N. The relathionship between epicuticular long-chained hydrocarbons and surface area volume ratios in insects (Diptera, Hymenoptera, Lepidoptera). PLoS ONE 2017, 12, e0175001. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Lab SHB Mean + SD | Field SHB Mean + SD | Honey Bees Mean + SD | χ2 | p | Lab SHB vs. Field SHB | Lab SHB vs. Honey Bees | Field SHB vs. Honey Bees | Loadings PLSDA1 | Loadings PLSDA2 |
|---|---|---|---|---|---|---|---|---|---|---|
| C19:1 *,** | 0 ± 0 | 0 ± 0 | 0.26 ± 0.48 | 17.158 | <0.001 | n.s. | <0.001 | <0.001 | 0.138 | −0.132 |
| n-C19 * | 0 ± 0 | 0.03 ± 0.24 | 1.99 ± 3.55 | 72.880 | <0.001 | n.s. | <0.001 | <0.001 | 0.214 | −0.004 |
| C21:1 * | 0 ± 0 | 0.04 ± 0.19 | 0.31 ± 0.38 | 26.821 | <0.001 | 0.012 | <0.001 | <0.001 | −0.186 | −0.088 |
| n-C21 * | 0 ± 0 | 0.94 ± 0.61 | 10.01 ± 8.41 | 22.086 | <0.001 | <0.001 | <0.001 | n.s. | 0.135 | −0.052 |
| C22:1 | 0.32 ± 0.73 | 0.33 ± 0.56 | 0.11 ± 0.24 | 1.544 | 0.492 | n.s. | n.s. | n.s. | 0.101 | 0.211 |
| n-C22 * | 0 ± 0 | 0.29 ± 0.56 | 3.15 ± 2.87 | 40.414 | <0.001 | n.s. | <0.001 | <0.001 | −0.047 | −0.011 |
| Unidentified ester *** | 0.33 ± 0.55 | 0.42 ± 0.55 | 0 ± 0 | 22.007 | <0.001 | n.s. | <0.001 | <0.001 | 0.158 | 0.108 |
| C23:1a | 2.16 ± 0.008 | 10.02 ± 6.21 | 14.99 ± 23.26 | 19.498 | <0.001 | n.s | 0.046 | <0.0001 | −0.116 | 0.031 |
| C23:1b * | 0 ± 0 | 0.70 ± 0.61 | 2.04 ± 2.86 | 14.413 | 0.001 | 0.003 | <0.001 | n.s. | −0.187 | −0.052 |
| n-C23 | 2.23 ± 1.36 | 10.74 ± 5.34 | 196.32 ± 295.13 | 0.371 | 0.856 | n.s. | n.s. | n.s. | −0.118 | 0.117 |
| C24:1a * | 0 ± 0 | 0.02 ± 0.11 | 0.32 ± 1.12 | 8.203 | 0.021 | n.s. | n.s. | 0.032 | 0.004 | 0.212 |
| C24:1b | 0.08 ± 0.25 | 0.17 ± 0.35 | 1.05 ± 1.94 | 11.404 | 0.005 | n.s. | 0.046 | 0.012 | −0.013 | 0.086 |
| n-C24 * | 0 ± 0 | 1.10 ± 0.55 | 14.67 ± 12.86 | 23.262 | <0.001 | <0.001 | 0.001 | n.s. | 0.075 | −0.093 |
| C25:2 * | 0 ± 0 | 0.08 ± 0.35 | 1.65 ± 5.47 | 23.887 | <0.001 | n.s. | 0.015 | <0.001 | 0.076 | 0.033 |
| C25:1a | 9.97 ± 8.12 | 17.12 ± 9.38 | 44.79 ± 69.01 | 29.544 | <0.001 | 0.015 | <0.001 | <0.001 | 0.054 | 0.242 |
| C25:1b | 0.49 ± 0.69 | 1.66 ± 1.09 | 7.07 ± 13.19 | 7.711 | 0.026 | n.s. | n.s. | 0.05 | 0.101 | −0.026 |
| n-C25 | 4.80 ± 3.28 | 16.94 ± 7.54 | 373.49 ± 482.33 | 0.259 | 0.892 | n.s. | n.s. | n.s. | −0.145 | −0.006 |
| meC25 | 0.90 ± 1.57 | 0.58 ± 0.72 | 8.21 ± 5.43 | 15.768 | 0.001 | n.s. | 0.044 | <0.001 | −0.031 | 0.224 |
| C26:1a | 0.09 ± 0.30 | 1.67 ± 1.48 | 0.94 ± 1.66 | 20.703 | <0.001 | 0.005 | 0.005 | <0.001 | 0.013 | −0.007 |
| C26:1b * | 0 ± 0 | 0.22 ± 0.53 | 0.33 ± 0.53 | 7.724 | 0.026 | n.s. | 0.047 | n.s. | 0.123 | 0.026 |
| n-C26 * | 0 ± 0 | 0.21 ± 0.45 | 18.38 ± 14.564 | 51.920 | <0.001 | n.s. | <0.001 | <0.001 | −0.062 | 0.206 |
| meC26a | 1.19 ± 0.49 | 1.41 ± 0.79 | 0.87 ± 1.35 | 41.729 | <0.001 | 0.041 | <0.001 | <0.001 | 0.043 | 0.046 |
| meC26b * | 0 ± 0 | 0.57 ± 0.83 | 0.48 ± 1.36 | 5.428 | 0.077 | n.s. | n.s. | n.s. | 0.171 | 0.070 |
| C27:1a | 1.89 ± 1.64 | 4.94 ± 2.45 | 28.30 ± 42.73 | 9.322 | 0.013 | n.s. | n.s. | 0.007 | −0.140 | −0.092 |
| C27:1b | 0.26 ± 0.58 | 0.47 ± 2.49 | 9.18 ± 15.63 | 43.211 | <0.001 | n.s. | 0.011 | <0.001 | −0.001 | 0.067 |
| n-C27 | 4.73 ± 3.81 | 14.88 ± 9.64 | 501.33 ± 341.54 | 0.073 | 0.964 | n.s. | n.s. | n.s. | −0.085 | 0.100 |
| meC27a | 2.25 ± 2.54 | 0.04 ± 0.19 | 0.02 ± 0.16 | 28.048 | <0.001 | <0.001 | <0.001 | n.s. | 0.153 | 0.010 |
| meC27b | 0.87 ± 1.76 | 1.10 ± 0.77 | 23.05 ± 27.22 | 6.683 | 0.042 | n.s. | 0.019 | n.s. | 0.003 | −0.018 |
| C28:1a | 0.04 ± 0.22 | 0.38 ± 0.64 | 1.48 ± 2.70 | 17.695 | <0.001 | 0.010 | n.s. | <0.001 | −0.078 | −0.210 |
| C28:1b * | 0 ± 0 | 0.33 ± 0.46 | 0.84 ± 1.04 | 7.487 | 0.029 | n.s. | 0.008 | n.s. | 0.047 | 0.148 |
| n-C28 | 0.45 ± 0.62 | 1.03 ± 0.87 | 12.65 ± 8.86 | 4.377 | 0.128 | n.s. | n.s. | n.s. | 0.099 | −0.184 |
| meC28 ** | 0.20 ± 0.44 | 0 ± 0 | 1.63 ± 2.61 | 35.014 | <0.001 | 0.002 | n.s. | <0.001 | 0.036 | 0.092 |
| C29:2a * | 0 ± 0 | 0.44 ± 0.61 | 1.22 ± 2.69 | 6.402 | 0.048 | <0.05 | n.s. | n.s. | 0.072 | 0.142 |
| C29:2b | 0.08 ± 0.25 | 0.01 ± 0.10 | 0.06 ± 0.31 | 1.764 | 0.413 | n.s. | n.s. | n.s. | 0.131 | −0.140 |
| C29:1a | 1.55 ± 1.21 | 2.33 ± 1.62 | 9.91 ± 18.72 | 37.454 | <0.001 | n.s. | <0.001 | <0.001 | −0.018 | 0.140 |
| C29:1b * | 0 ± 0 | 0.26 ± 1.71 | 20.66 ± 20.56 | 55.416 | <0.001 | n.s. | <0.001 | <0.001 | 0.004 | −0.085 |
| n-C29 | 12.42 ±6.83 | 22.11 ± 15.45 | 294.39 ± 204.54 | 8.662 | 0.017 | n.s. | <0.05 | n.s. | −0.175 | 0.096 |
| meC29a | 0.07 ± 0.22 | 0.69 ± 0.68 | 18.15 ± 24.33 | 15.965 | 0.001 | 0.029 | <0.001 | 0.036 | 0.201 | −0.039 |
| me-C29b | 0.19 ± 0.42 | 0.01 ± 0.11 | 2.22 ± 3.55 | 25.779 | <0.001 | 0.030 | n.s. | <0.001 | −0.061 | −0.007 |
| C30:1 * | 0 ± 0 | 0.07 ± 0.33 | 2.63 ± 2.62 | 54.909 | <0.001 | n.s. | <0.001 | <0.001 | 0.108 | 0.137 |
| n-C30 | 0.13 ± 0.41 | 0.22 ± 0.51 | 7.05 ± 6.33 | 47.351 | <0.001 | n.s. | <0.001 | <0.001 | 0.122 | −0.141 |
| meC30 *,** | 0 ± 0 | 0 ± 0 | 0.49 ± 0.93 | 17.158 | <0.001 | n.s. | <0.001 | <0.001 | 0.198 | −0.031 |
| C31:2a *,** | 0 ± 0 | 0 ± 0 | 0.31 ± 0.94 | 8.694 | 0.017 | n.s. | 0.022 | 0.014 | 0.176 | 0.035 |
| C31:2b * | 0 ± 0 | 0.04 ± 0.27 | 4.26 ± 4.63 | 55.987 | <0.001 | n.s. | <0.001 | <0.001 | 0.114 | −0.075 |
| C31:1a * | 0 ± 0 | 4.69 ± 7.10 | 89.27 ± 70.97 | 24.395 | <0.001 | <0.001 | <0.001 | n.s. | 0.092 | −0.066 |
| C31:1b * | 0 ± 0 | 4.10 ± 6.16 | 87.23 ± 71.41 | 25.265 | <0.001 | <0.001 | <0.001 | n.s. | 0.197 | −0.043 |
| n-C31 | 1.21 ± 0.92 | 4.74 ± 7.48 | 196.39 ± 158.69 | 3.335 | 0.211 | n.s. | n.s. | n.s. | 0.083 | 0.314 |
| meC31a ** | 0.08 ± 0.25 | 0 ± 0 | 6.30 ± 9.13 | 64.691 | <0.001 | 0.020 | <0.001 | <0.001 | 0.068 | 0.292 |
| meC31b *,** | 0 ± 0 | 0 ± 0 | 1.52 ± 1.74 | 53.756 | <0.001 | n.s. | <0.001 | <0.001 | 0.012 | 0.140 |
| C32:1 * | 0 ± 0 | 0.11 ± 0.59 | 8.07 ± 6.43 | 69.219 | <0.001 | n.s. | <0.001 | <0.001 | 0.202 | −0.045 |
| C33:2 | 3.96 ± 3.34 | 3.49 ± 3.12 | 22.24 ± 23.78 | 3.129 | 0.230 | n.s. | n.s. | n.s. | 0.192 | −0.070 |
| C33:1 | 0.29 ± 0.94 | 9.73 ± 15.77 | 263.50 ± 190.31 | 24.503 | <0.001 | <0.001 | <0.001 | n.s. | 0.207 | −0.002 |
| n-C33 * | 0 ± 0 | 0.01 ± 0.11 | 14.76 ± 26.10 | 27.807 | <0.001 | n.s. | <0.001 | <0.001 | −0.012 | −0.002 |
| meC33 * | 0 ± 0 | 0.53 ± 1.85 | 1.16 ± 2.26 | 9.590 | 0.008 | n.s. | n.s. | 0.027 | 0.077 | 0.309 |
| C35:2 * | 0 ± 0 | 0.03 ± 0.24 | 2.30 ± 2.97 | 35.511 | <0.001 | n.s. | 0.007 | <0.001 | 0.167 | −0.125 |
| C35:1a * | 0 ± 0 | 0.04 ± 0.30 | 4.18 ± 4.80 | 43.226 | <0.001 | n.s. | 0.002 | <0.001 | 0.083 | −0.018 |
| C35:1b * | 0 ± 0 | 0 ± 0 | 3.40 ± 7.35 | 28.375 | <0.001 | n.s. | 0.019 | <0.001 | 0.173 | −0.098 |
| Oleic acid ester 1 * | 0 ± 0 | 0.65 ± 2.27 | 19.59 ± 18.43 | 40.351 | <0.001 | n.s. | 0.001 | <0.001 | 0.179 | −0.057 |
| Oleic acid ester 2 * | 0 ± 0 | 0.85 ± 4.98 | 46.01 ± 47.44 | 53.180 | <0.001 | n.s. | <0.001 | <0.001 | 0.106 | 0.053 |
| Oleic acid ester 3 | 29.01 ± 19.68 | 9.55 ± 8.87 | 7.95 ± 30.33 | 43.506 | <0.001 | n.s. | <0.001 | <0.001 | −0.012 | −0.107 |
| Oleic acid ester 4 | 9.58 ± 7.28 | 3.44 ± 3.99 | 3.91 ± 20.67 | 39.310 | <0.001 | n.s. | <0.001 | <0.001 | 0.158 | 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papach, A.; Cappa, F.; Cervo, R.; Dapporto, L.; Balusu, R.; Williams, G.R.; Neumann, P. Cuticular Hydrocarbon Profile of Parasitic Beetles, Aethina tumida (Coleoptera: Nitidulidae). Insects 2021, 12, 751. https://doi.org/10.3390/insects12080751
Papach A, Cappa F, Cervo R, Dapporto L, Balusu R, Williams GR, Neumann P. Cuticular Hydrocarbon Profile of Parasitic Beetles, Aethina tumida (Coleoptera: Nitidulidae). Insects. 2021; 12(8):751. https://doi.org/10.3390/insects12080751
Chicago/Turabian StylePapach, Anna, Federico Cappa, Rita Cervo, Leonardo Dapporto, Rammohan Balusu, Geoffrey R. Williams, and Peter Neumann. 2021. "Cuticular Hydrocarbon Profile of Parasitic Beetles, Aethina tumida (Coleoptera: Nitidulidae)" Insects 12, no. 8: 751. https://doi.org/10.3390/insects12080751
APA StylePapach, A., Cappa, F., Cervo, R., Dapporto, L., Balusu, R., Williams, G. R., & Neumann, P. (2021). Cuticular Hydrocarbon Profile of Parasitic Beetles, Aethina tumida (Coleoptera: Nitidulidae). Insects, 12(8), 751. https://doi.org/10.3390/insects12080751