Solitary Living Brings a Decreased Weight and an Increased Agility to the Domestic Silkworm, Bombyx mori

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Solitary Breeding
2.2. Behavior Experiments
2.3. mRNA and miRNA Sequencing
2.4. Identification of Differently Expressed Genes and miRNAs
2.5. Functional Analysis
2.6. Statistics
3. Results
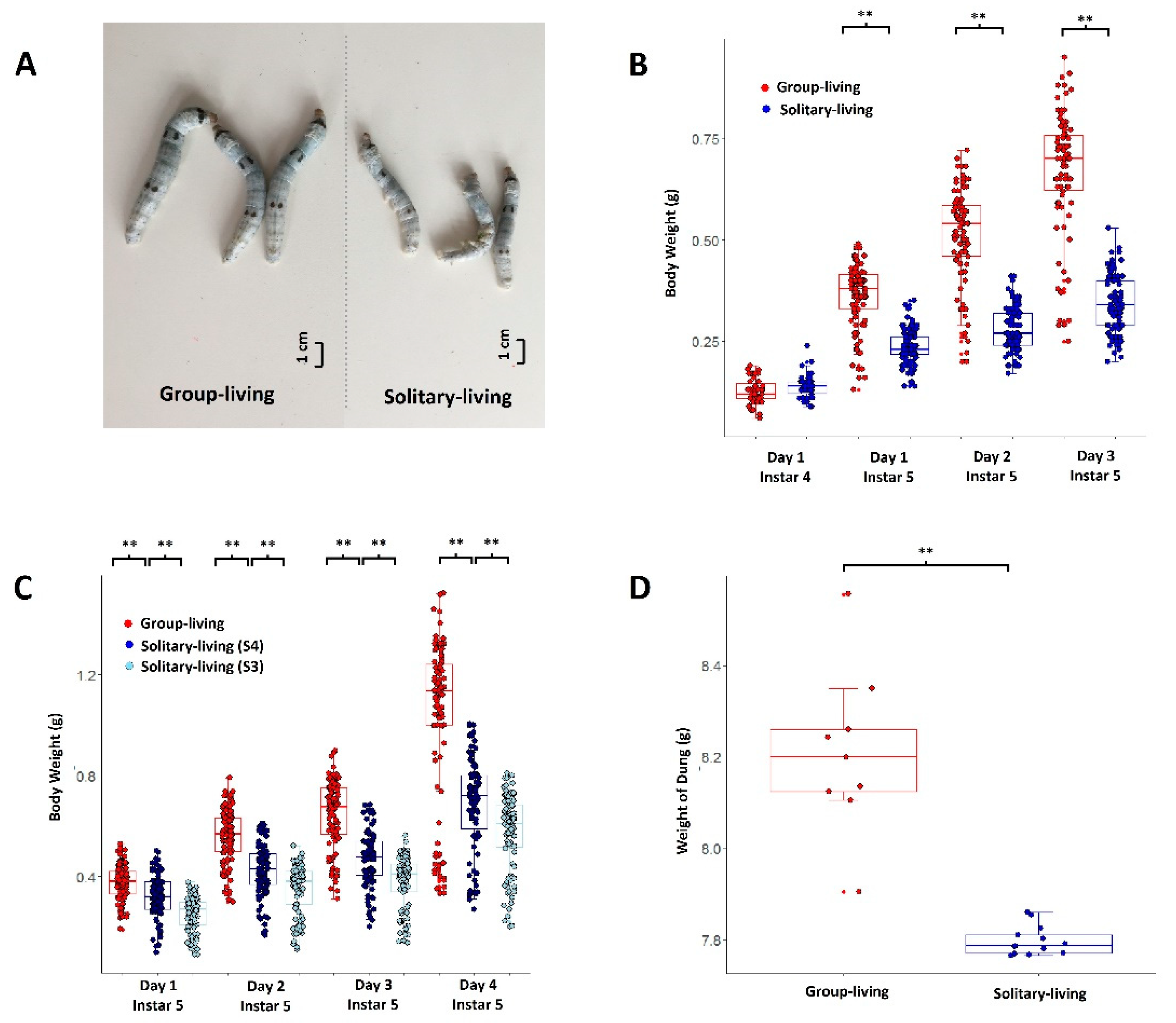
3.1. Solitary Living Brought a Loss of Body Weight
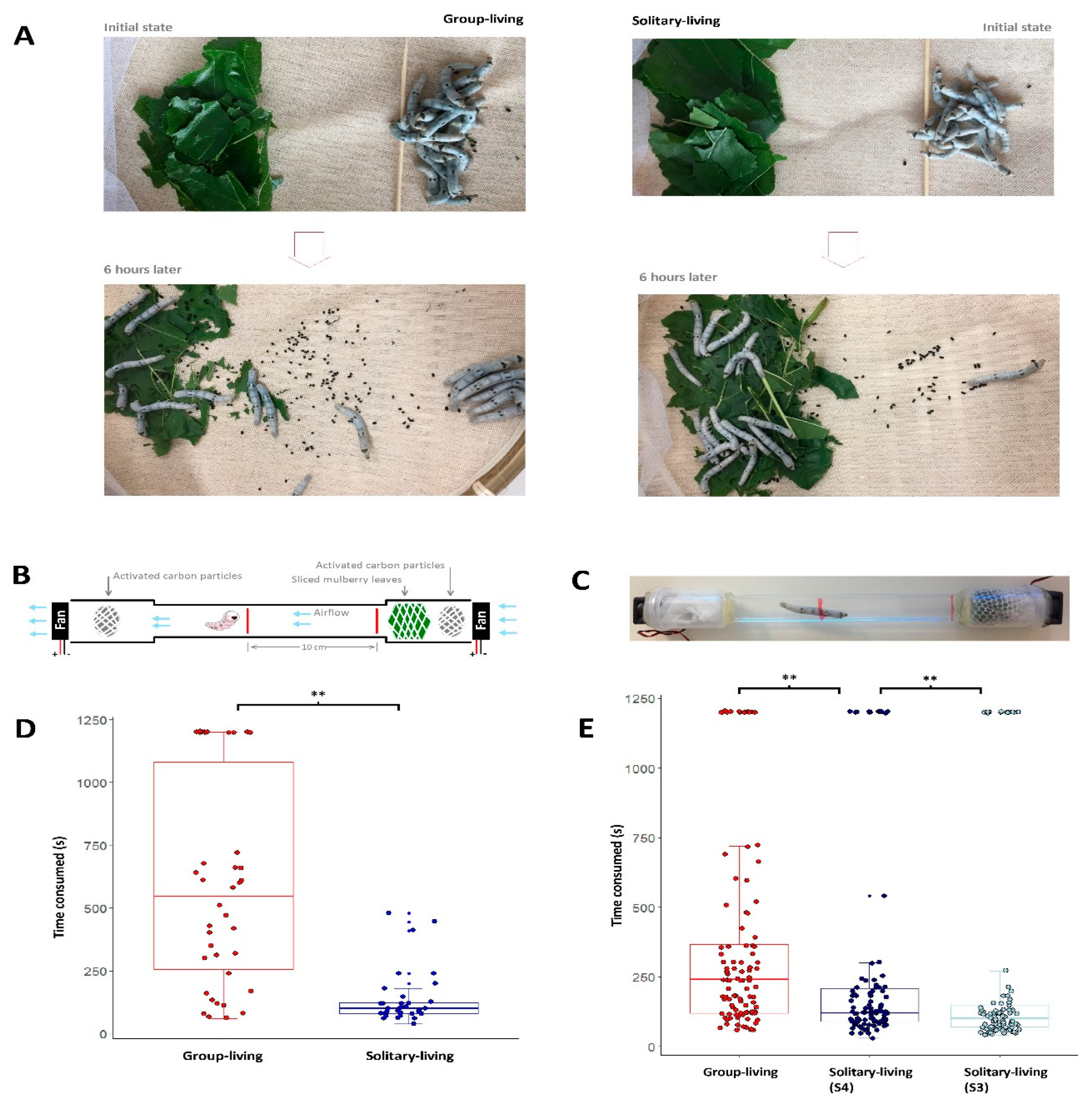
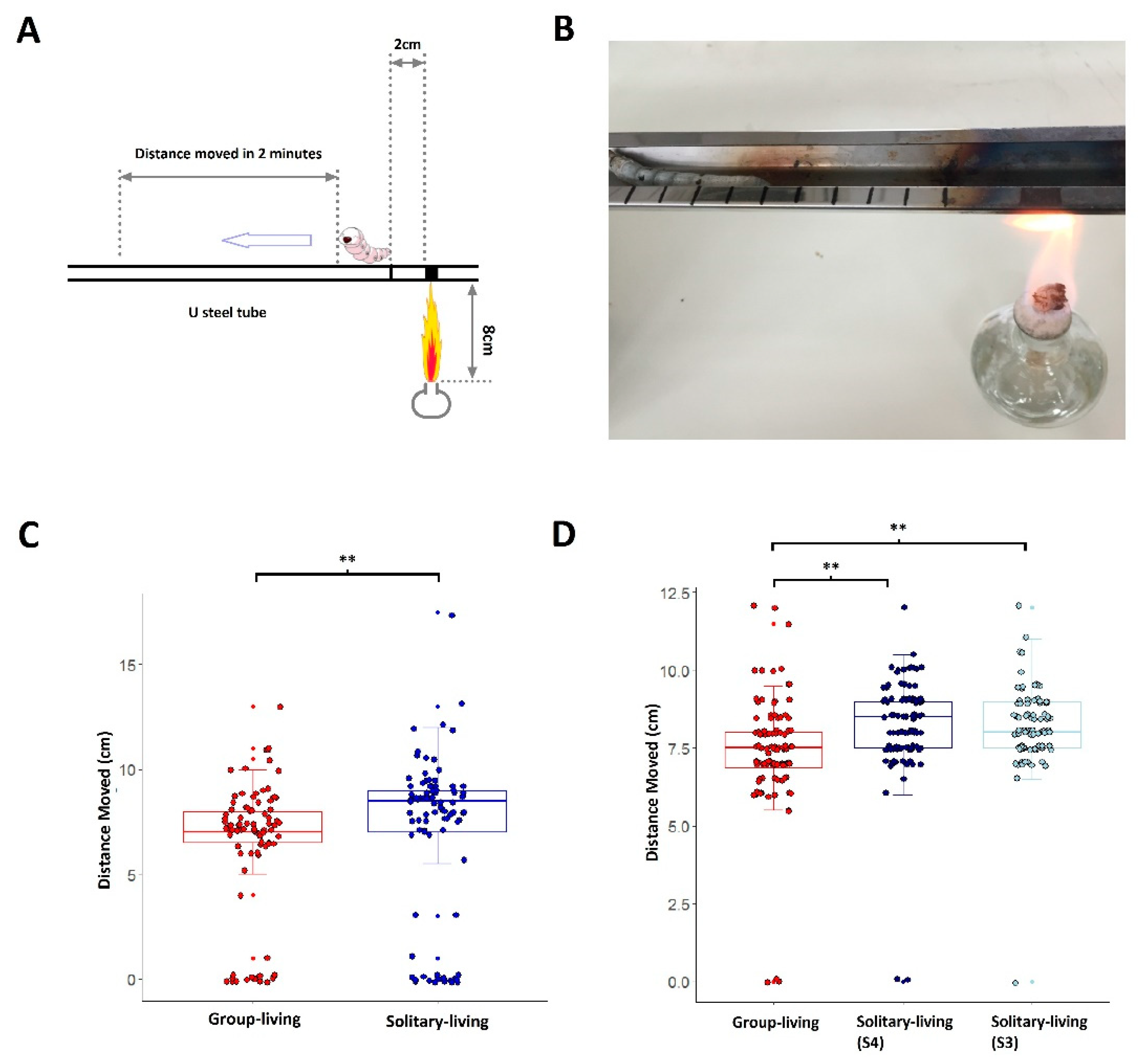
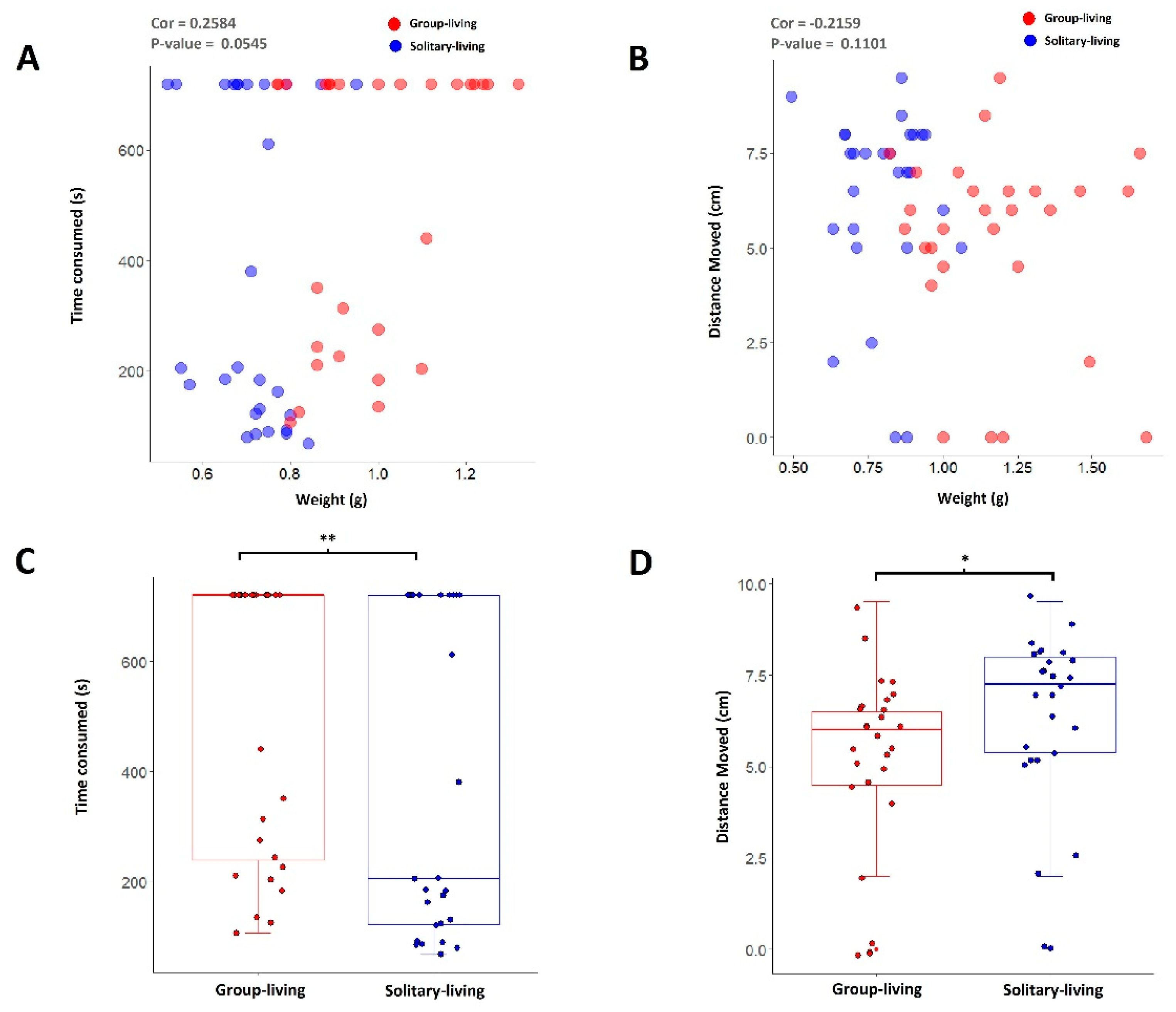
3.2. Solitary-Living Silkworms Have an Increased Agility
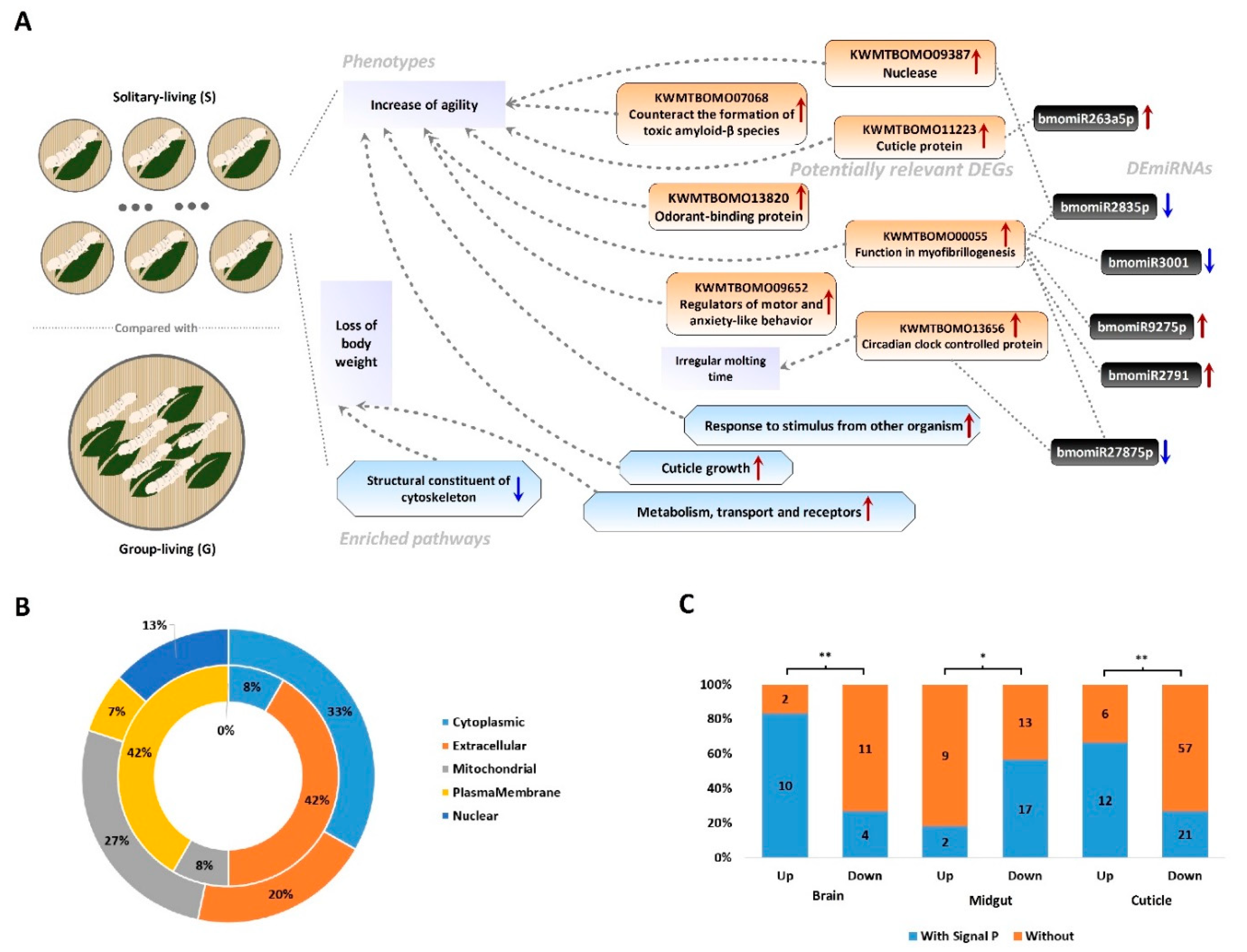
3.3. Potential Molecular Mechanisms Associated with the Phenotypic Change Led by Solitary Living
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xia, N.; Li, H. Loneliness, social isolation, and cardiovascular health. Antioxid. Redox Signal. 2018, 28, 837–851. [Google Scholar] [CrossRef] [Green Version]
- Perissinotto, C.M.; Stijacic Cenzer, I.; Covinsky, K.E. Loneliness in older persons: A predictor of functional decline and death. Arch. Intern. Med. 2012, 172, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Hawkley, L.C.; Waite, L.J.; Cacioppo, J.T. Loneliness, health, and mortality in old age: A national longitudinal study. Soc. Sci. Med. 2012, 74, 907–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaker, E.D.; Pinsky, J.; Castelli, W.P. Myocardial infarction and coronary death among women: Psychosocial predictors from a 20-year follow-up of women in the framingham study. Am. J. Epidemiol. 1992, 135, 854–864. [Google Scholar] [CrossRef] [PubMed]
- Cacioppo, S.; Capitanio, J.P.; Cacioppo, J.T. Toward a neurology of loneliness. Psychol. Bull. 2014, 140, 1464–1504. [Google Scholar] [CrossRef] [PubMed]
- Cacioppo, J.T.; Cacioppo, S.; Cole, S.W.; Capitanio, J.P.; Goossens, L.; Boomsma, D.I. Loneliness across phylogeny and a call for comparative studies and animal models. Perspect. Psychol. Sci. A J. Assoc. Psychol. Sci. 2015, 10, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Cacioppo, S.; Cacioppo, J.T. Why may allopregnanolone help alleviate loneliness? Med. Hypotheses 2015, 85, 947–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacioppo, J.T.; Cacioppo, S.; Capitanio, J.P.; Cole, S.W. The neuroendocrinology of social isolation. Annu. Rev. Psychol. 2015, 66, 733–767. [Google Scholar] [CrossRef] [Green Version]
- Siuda, D.; Wu, Z.; Chen, Y.; Guo, L.; Linke, M.; Zechner, U.; Xia, N.; Reifenberg, G.; Kleinert, H.; Forstermann, U.; et al. Social isolation-induced epigenetic changes in midbrain of adult mice. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2014, 65, 247–255. [Google Scholar]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar]
- Sachser, N.; Durschlag, M.; Hirzel, D. Social relationships and the management of stress. Psychoneuroendocrinology 1998, 23, 891–904. [Google Scholar] [CrossRef]
- Capitanio, J.P.; Mendoza, S.P.; Lerche, N.W.; Mason, W.A. Social stress results in altered glucocorticoid regulation and shorter survival in simian acquired immune deficiency syndrome. Proc. Natl. Acad. Sci. USA 1998, 95, 4714–4719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachser, N.; Lick, C. Social stress in guinea pigs. Physiol. Behav. 1989, 46, 137–144. [Google Scholar] [CrossRef]
- Kappeler, P.M.; Cremer, S.; Nunn, C.L. Sociality and health: Impacts of sociality on disease susceptibility and transmission in animal and human societies. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittwer, B.; Hefetz, A.; Simon, T.; Murphy, L.E.K.; Elgar, M.A.; Pierce, N.E.; Kocher, S.D. Solitary bees reduce investment in communication compared with their social relatives. Proc. Natl. Acad. Sci. USA 2017, 114, 6569–6574. [Google Scholar] [CrossRef] [Green Version]
- Madabhavi, I.; Sarkar, M.; Kadakol, N. Covid-19: A review. Monaldi Arch. Chest Dis. Arch. Monaldi Per Le Mal. Del Torace 2020, 90, 2. [Google Scholar] [CrossRef]
- Mehrkam, L.R.; Hall, N.J.; Haitz, C.; Wynne, C.D.L. The influence of breed and environmental factors on social and solitary play in dogs (canis lupus familiaris). Learn. Behav. 2017, 45, 367–377. [Google Scholar] [CrossRef]
- Vanthournout, B.; Greve, M.; Bruun, A.; Bechsgaard, J.; Overgaard, J.; Bilde, T. Benefits of group living include increased feeding efficiency and lower mass loss during desiccation in the social and inbreeding spider stegodyphus dumicola. Front. Physiol. 2016, 7, 18. [Google Scholar] [CrossRef]
- Kang, L.; Chen, X.; Zhou, Y.; Liu, B.; Zheng, W.; Li, R.; Wang, J.; Yu, J. The analysis of large-scale gene expression correlated to the phase changes of the migratory locust. Proc. Natl. Acad. Sci. USA 2004, 101, 17611–17615. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Wang, X.; Ma, Z.; Xue, L.; Han, J.; Yu, D.; Kang, L. Csp and takeout genes modulate the switch between attraction and repulsion during behavioral phase change in the migratory locust. PLoS Genet. 2011, 7, e1001291. [Google Scholar] [CrossRef]
- Ichinose, T.; Tanimoto, H. Dynamics of memory-guided choice behavior in drosophila. Proceedings of the Japan Academy. Ser. B Phys. Biol. Sci. 2016, 92, 346–357. [Google Scholar]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Ballgown bridges the gap between transcriptome assembly and expression analysis. Nat. Biotechnol. 2015, 33, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of rna-seq experiments with hisat, stringtie and ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Kawamoto, M.; Jouraku, A.; Toyoda, A.; Yokoi, K.; Minakuchi, Y.; Katsuma, S.; Fujiyama, A.; Kiuchi, T.; Yamamoto, K.; Shimada, T. High-quality genome assembly of the silkworm, bombyx mori. Insect Biochem. Mol. Biol. 2019, 107, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chan, C.K. Analysis of rna-seq data using tophat and cufflinks. Methods Mol. Biol. 2016, 1374, 339–361. [Google Scholar] [PubMed]
- Pollier, J.; Rombauts, S.; Goossens, A. Analysis of rna-seq data with tophat and cufflinks for genome-wide expression analysis of jasmonate-treated plants and plant cultures. Methods Mol. Biol. 2013, 1011, 305–315. [Google Scholar]
- Langdon, W.B. Performance of genetic programming optimised bowtie2 on genome comparison and analytic testing (gcat) benchmarks. BioData Min. 2015, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. Htseq—A python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. Degseq: An r package for identifying differentially expressed genes from rna-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Kruger, J.; Rehmsmeier, M. Rnahybrid: Microrna target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef]
- Sturm, M.; Hackenberg, M.; Langenberger, D.; Frishman, D. Targetspy: A supervised machine learning approach for microrna target prediction. BMC Bioinform. 2010, 11, 292. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An r package for comparing biological themes among gene clusters. Omics A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Guan, Z.; Liu, G.; Wang, Y.; Zhang, Z. SGID: A comprehensive and interactive database of the silkworm. Database J. Biol. Databases Curation 2019, 2019, baz134. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Yu, C.S.; Lin, C.J.; Hwang, J.K. Predicting subcellular localization of proteins for gram-negative bacteria by support vector machines based on n-peptide compositions. Protein Sci. 2004, 13, 1402–1406. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, H. Predicting secretory proteins with signalp. Methods Mol. Biol. 2017, 1611, 59–73. [Google Scholar]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Signalp 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Claridge-Chang, A.; Roorda, R.D.; Vrontou, E.; Sjulson, L.; Li, H.; Hirsh, J.; Miesenbock, G. Writing memories with light-addressable reinforcement circuitry. Cell 2009, 139, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kaminska, K. Microrna in cardiovascular biology and disease. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2017, 26, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Diaz, G.; Zamboni, F.; Tice, A.; Farci, P. Integrated ordination of miRNA and mRNA expression profiles. BMC Genom. 2015, 16, 767. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.J.; Underhill, D.M. Peptidoglycan recognition by the innate immune system. Nature reviews. Immunology 2018, 18, 243–254. [Google Scholar] [PubMed]
- Arentsen, T.; Khalid, R.; Qian, Y.; Diaz Heijtz, R. Sex-dependent alterations in motor and anxiety-like behavior of aged bacterial peptidoglycan sensing molecule 2 knockout mice. Brain Behav. Immun. 2018, 67, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.C. Neuroinflammation and the cgas-sting pathway. J. Neurophysiol. 2019, 121, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Helmfors, L.; Boman, A.; Civitelli, L.; Nath, S.; Sandin, L.; Janefjord, C.; McCann, H.; Zetterberg, H.; Blennow, K.; Halliday, G.; et al. Protective properties of lysozyme on beta-amyloid pathology: Implications for alzheimer disease. Neurobiol. Dis. 2015, 83, 122–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balter, L.J.T.; Raymond, J.E.; Aldred, S.; Drayson, M.T.; Veldhuijzen van Zanten, J.; Higgs, S.; Bosch, J.A. Loneliness in healthy young adults predicts inflammatory responsiveness to a mild immune challenge in vivo. Brain Behav. Immun. 2019, 82, 298–301. [Google Scholar] [CrossRef]
- Goossens, L.; van Roekel, E.; Verhagen, M.; Cacioppo, J.T.; Cacioppo, S.; Maes, M.; Boomsma, D.I. The genetics of loneliness: Linking evolutionary theory to genome-wide genetics, epigenetics, and social science. Perspect. Psychol. Sci. A J. Assoc. Psychol. Sci. 2015, 10, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.P. Loneliness and eating disorders. J. Psychol. 2012, 146, 243–257. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Tan, Y.; Xiao, S.; Guan, Z.; Zhao, W.; Dai, Z.; Liu, G.; Zhang, Z. Solitary Living Brings a Decreased Weight and an Increased Agility to the Domestic Silkworm, Bombyx mori. Insects 2021, 12, 809. https://doi.org/10.3390/insects12090809
Zhu Z, Tan Y, Xiao S, Guan Z, Zhao W, Dai Z, Liu G, Zhang Z. Solitary Living Brings a Decreased Weight and an Increased Agility to the Domestic Silkworm, Bombyx mori. Insects. 2021; 12(9):809. https://doi.org/10.3390/insects12090809
Chicago/Turabian StyleZhu, Zhenglin, Yuting Tan, Siyu Xiao, Zhufen Guan, Wen Zhao, Zhijun Dai, Gexin Liu, and Ze Zhang. 2021. "Solitary Living Brings a Decreased Weight and an Increased Agility to the Domestic Silkworm, Bombyx mori" Insects 12, no. 9: 809. https://doi.org/10.3390/insects12090809
APA StyleZhu, Z., Tan, Y., Xiao, S., Guan, Z., Zhao, W., Dai, Z., Liu, G., & Zhang, Z. (2021). Solitary Living Brings a Decreased Weight and an Increased Agility to the Domestic Silkworm, Bombyx mori. Insects, 12(9), 809. https://doi.org/10.3390/insects12090809