
Chinese Cabbage Changes Its Release of Volatiles to Defend against Spodoptera litura

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Insect
2.3. Olfactory Preferences of M. similis (M. similis Olfactometer Experiments 1–4)
2.4. Behavioral Responses of S. litura Larvae to Chinese Cabbage and Volatile Compounds (S. litura Petri Dish Experiments 1 and 2, and S. litura Olfactometer Experiments 1 and 2)
2.5. Collection and Analysis of Volatiles via Solid Phase Microextraction
2.6. Statistical Analysis
3. Results
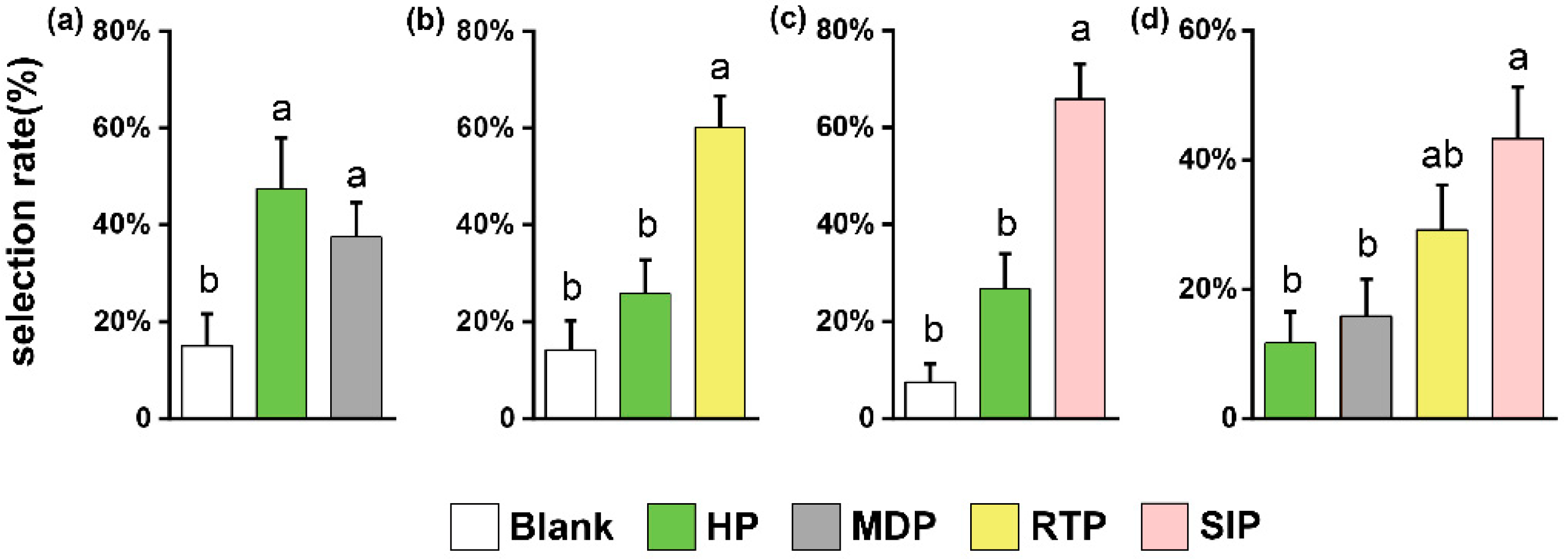
3.1. Olfactory Preferences of M. similis (M. similis Olfactometer Experiments 1–4)
3.2. Feeding and Olfactory Selecting Behavior of S. litura Larvae (S. litura Petri Dish Experiment 1 and 2, and S. litura Olfactometer Experiments 1 and 2)
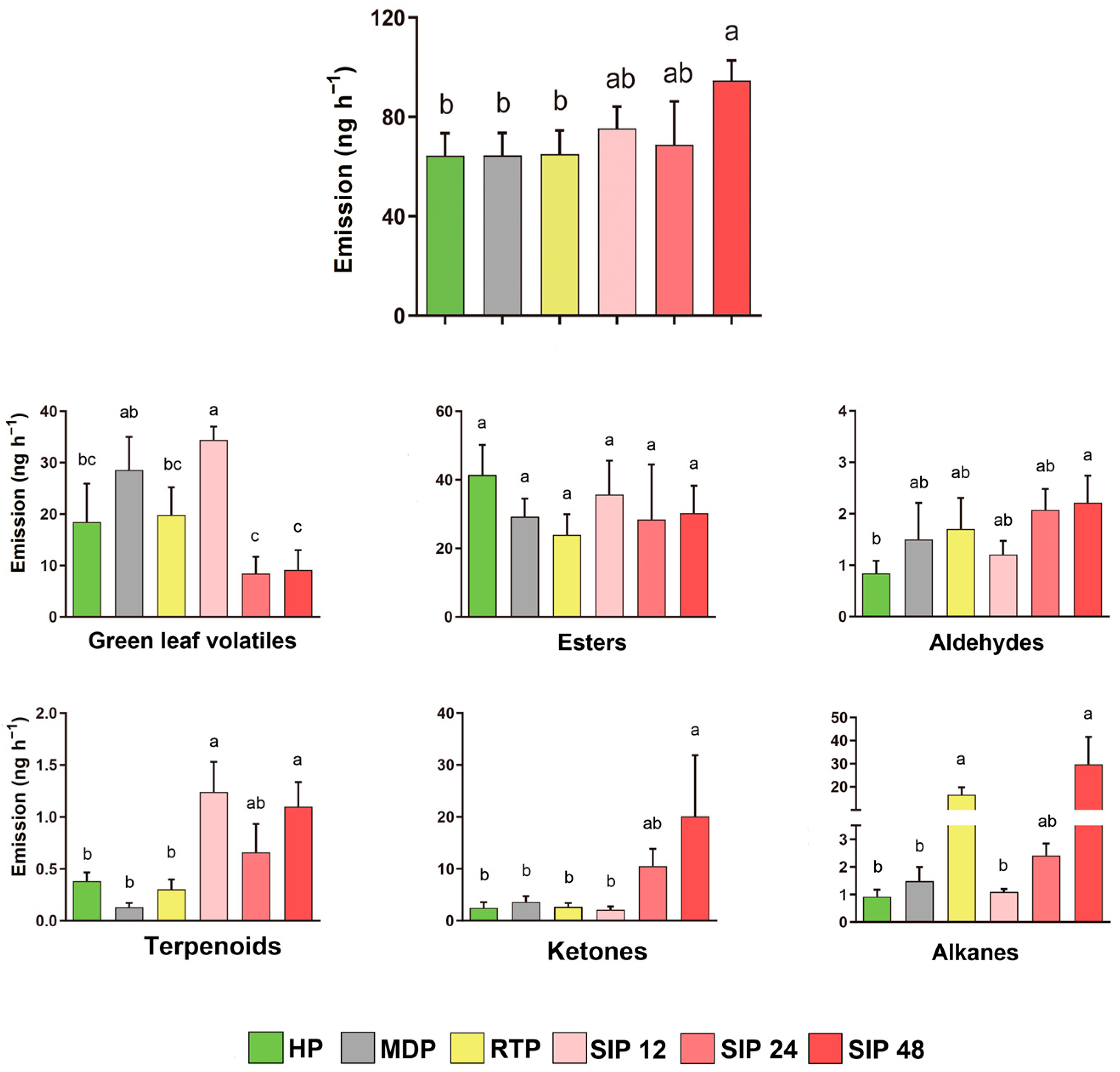
3.3. Quantities of Total and Six Classes of Volatiles Released
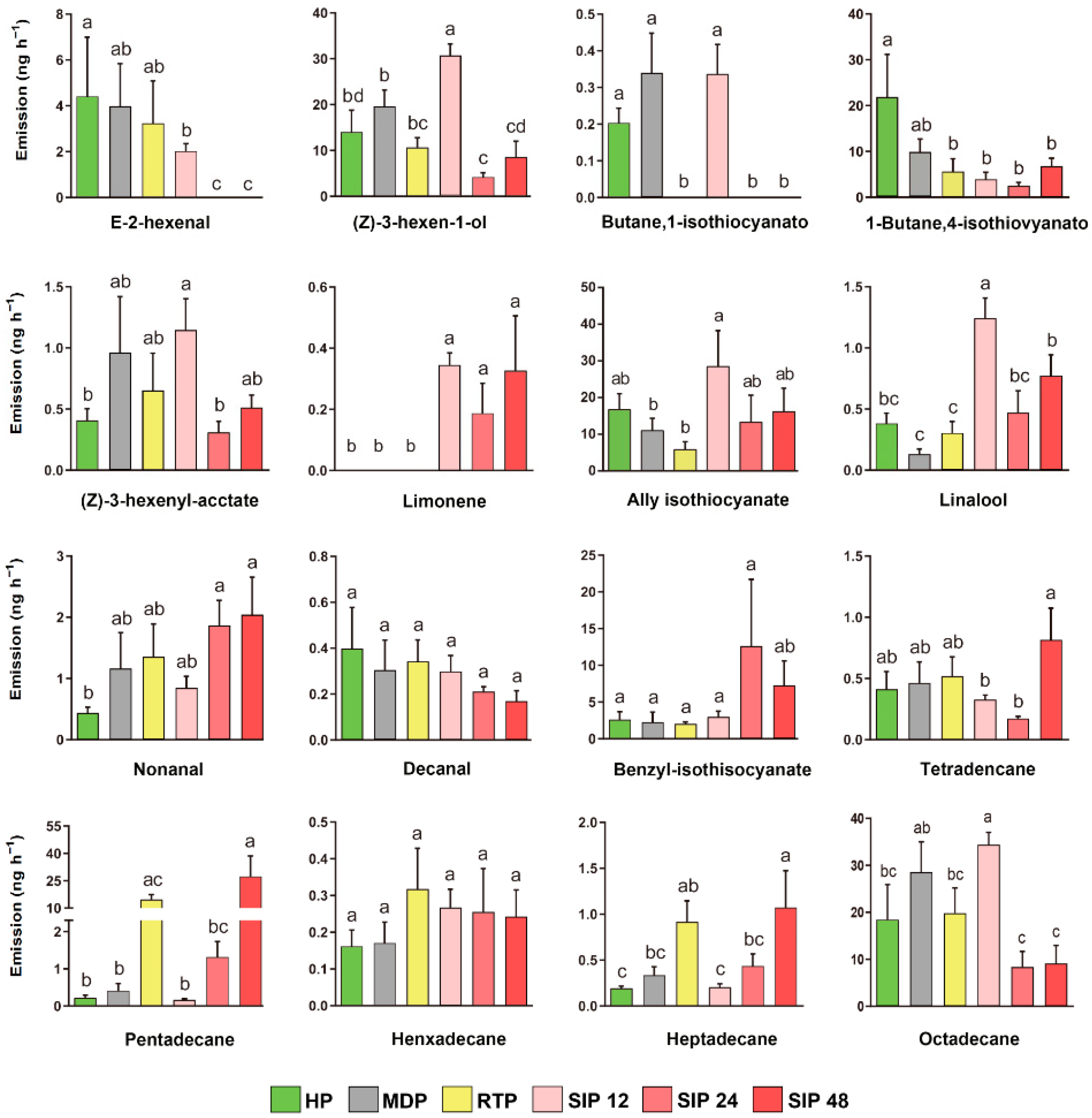
3.4. Quantities of 16 Specific Compounds Released
3.5. Relationships between the Release of Classes of Volatiles and the Host Searching Behavior of M. similis and S. litura
3.6. Relationship between the Release of Specific Volatiles by Chinese Cabbage and the Host Searching Behavior of M. similis and S. litura
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horiuchi, J.I.; Arimura, G.I.; Ozawa, R.; Shimoda, T.; Takabayashi, J.; Nishioka, T. A comparison of the responses of Tetranychus urticae (Acari: Tetranychidae) and Phytoseiulus persimilis (Acari: Phytoseiidae) to volatiles emitted from lima bean leaves with different levels of damage made by T. urticae or Spodoptera exigua (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 2003, 38, 109–116. [Google Scholar]
- Kanchiswamy, C.; Malnoy, M.; Maffei, M. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Fincheira, P.; Quiroz, A.; Tortella, G.; Diez, M.; Rubilar, O. Current advances in plant-microbe communication via volatile organic compounds as an innovative strategy to improve plant growth. Microbiol. Res. 2021, 247, 26726. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuman, M.C.; Baldwin, I.T. The layers of plant responses to insect herbivores. Annu. Rev. Entomol. 2006, 61, 373–394. [Google Scholar] [CrossRef] [PubMed]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef]
- Leitner, M.; Boland, W.; Mithöfer, A. Direct and indirect defences induced by piercing-sucking and chewing herbivores in Medicago truncatula. New Phytol. 2005, 167, 597–606. [Google Scholar] [CrossRef]
- Mumm, R.; Posthumus, M.A.; Dicke, M. Significance of terpenoids in induced indirect plant defence against herbivorous arthropods. Plant Cell Environ. 2008, 31, 575–585. [Google Scholar] [CrossRef]
- War, A.; Sharma, H.; Paulraj, M.; War, M.; Ignacimuthu, S. Herbivore induced plant volatiles: Their role in plant defense for pest management. Plant Signal. Behav. 2011, 6, 1973–1978. [Google Scholar] [CrossRef] [Green Version]
- Kessler, A.; Baldwin, T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef]
- Yosra, C.; Diane, L.; Turlings, T.C.J.; Desurmont, G.A. Impact of exotic insect herbivores on native tritrophic interactions: A case study of the African cotton leafworm, Spodoptera littoralis and insects associated with the field mustard Brassica rapa. J. Ecol. 2015, 103, 109–117. [Google Scholar]
- Desurmont, G.A.; Xu, H.; Turlings, T.C.J. Powdery mildew suppresses herbivore-induced plant volatiles and interferes with parasitoid attraction in Brassica rapa. Plant Cell Environ. 2016, 39, 1920–1927. [Google Scholar] [CrossRef] [Green Version]
- Turlings, T.C.J.; Tumlinson, J.H.; Lewis, W.J. Exploitation of herbivore-induced plant odors by host-seeking parasitic wasps. Science 1990, 250, 1251–1253. [Google Scholar] [CrossRef] [Green Version]
- Hare, J.D.; Sun, J.J. Production of induced volatiles by Datura wrightii in response to damage by insects: Effect of herbivore species and time. J. Chem. Ecol. 2011, 37, 751–764. [Google Scholar] [CrossRef]
- Stam, J.M.; Kroes, A.; Li, Y.; Gols, R.; Van Loon, J.J.A.; Poelman, E.H.; Dicke, M. Plant interactions with multiple insect herbivores: From community to genes. Annu. Rev. Plant Biol. 2014, 65, 689–713. [Google Scholar] [CrossRef]
- Gish, M.; De Moraes, C.M.; Mescher, M.C. Herbivore-induced plant volatiles in natural and agricultural ecosystems: Open questions and future prospects. Insect Sci. 2015, 9, 1–6. [Google Scholar] [CrossRef]
- Veyrat, N.; Robert, C.A.M.; Turlings, T.C.J.; Erb, M. Herbivore intoxication as a potential primary function of an inducible volatile plant signal. J. Ecol. 2016, 104, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Turlings, T.C.J.; Tumlinson, J.H. Do parasitoids use herbivore-induced plant chemical defences to locate hosts? Fla. Entomol. 1991, 74, 42–50. [Google Scholar] [CrossRef]
- Signoretti, A.G.C.; Peñaflor, M.F.G.V.; Bento, J.M.S. Fall Armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), female moths respond to herbivore-induced corn volatiles. Neotrop. Entomol. 2012, 41, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Carroll, M.J.; Schmelz, E.A.; Meagher, R.L.; Teal, P.E.A. Attraction of Spodoptera frugiperda Larvae to Volatiles from Herbivore-Damaged Maize Seedlings. J. Chem. Ecol. 2006, 32, 1911–1924. [Google Scholar] [CrossRef] [Green Version]
- Tatemoto, S.; Shimoda, T. Olfactory responses of the predatory mites (Neoseiulus cucumeris) and insects (Orius strigicollis) to two different plant species infested with onion thrips (Thrips tabaci). J. Chem. Ecol. 2008, 34, 605–613. [Google Scholar] [CrossRef]
- Pan, M.Z.; Liu, T.X. Suitability of three aphid species for Aphidius gifuensis (Hymenoptera: Braconidae): Parasitoid performance varies with hosts of origin. Biol. Control 2014, 69, 90–96. [Google Scholar] [CrossRef]
- De Moraes, C.M.; Lewis, W.J.; Paré, P.W.; Alborn, H.T.; Tumlinson, J.H. Herbivore-infested plants selectively attract parasitoids. Nature 1998, 393, 570–573. [Google Scholar] [CrossRef]
- Shiojiri, K.; Ozawa, R.; Matsui, K.; Sabelis, M.W.; Takabayashi, J. Intermittent exposure to traces of green leaf volatiles triggers a plant response. Sci. Rep. 2012, 2, 378. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.S.; Sun, W.; Lv, J.P.; Kuang, R.P. Use of sex pheromone for control of Spodoptera litura (Lepidoptera: Noctuidae). J. Entomol. Res. Soc. 2009, 11, 27–36. [Google Scholar]
- Li, S.J.; Huang, J.P.; Chang, Y.Y.; Quan, S.Y.; Yi, W.T.; Chen, Z.S.; Liu, S.Q.; Cheng, X.W.; Huang, G.H. Development of Microplitis similis (Hymenoptera: Braconidae) on two candidate host species, Spodoptera litura and Spodoptera exigua (Lepidoptera: Noctuidae). Fla. Entomol. 2015, 98, 736–741. [Google Scholar] [CrossRef]
- Yi, S.J.; Hopkins, R.J.; Chen, X.Y.; Chen, Z.M.; Wang, X.; Huang, G.H. Effects of temperature on the development and fecundity of Microplitis similis (Hymenoptera: Braconidae), a parasitoid of Spodoptera litura (Lepidoptera: Noctuidae). Physiol. Entomol. 2020, 45, 95–102. [Google Scholar] [CrossRef]
- Li, Z.Q.; Song, X.H.; Wang, M.; Wang, S.; Huang, G.H. Melanization induced by Heliothis virescens ascovirus 3 h promotes viral replication. Insect Sci. 2021, 28, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Turlings, T.C.J.; Mccall, P.J.; Alborn, H.T.; Tumlinson, J.H. An elicitor in caterpillar oral secretions that induces corn seedlings to emit chemical signals attractive to parasitic wasps. J. Chem. Ecol. 1993, 19, 411–425. [Google Scholar] [CrossRef] [Green Version]
- Turlings, T.C.J.; Davison, A.; TamÒ, C. A six-arm olfactometer permitting simultaneous observation of insect attraction and odor trapping. Physiol. Entomol. 2004, 29, 45–55. [Google Scholar] [CrossRef]
- Ahuja, I.; Rohloff, J.; Bones, A. Defence mechanisms of Brassicaceae: Implications for plant-insect interactions and potential for integrated pest management. A review. Agron. Sustain. Dev. 2010, 30, 311–348. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.L.; Felton, G.W. Foliar oxidative stress and insect herbivory: Primary compounds, secondary metabolites, and reactive oxygen species as components of induced resistance. Chem. Ecol. 1995, 21, 1511–1530. [Google Scholar] [CrossRef]
- Karban, R.; Myers, J.H. Induced plant responses to herbivory. Annu. Rev. Ecol. Syst. 1989, 20, 331–348. [Google Scholar] [CrossRef]
- Heil, M.; Koch, T.; Hilpert, A.; Fiala, B.; Boland, W.; Linsenmair, K.E. Extrafloral nectar production of the ant-associated plant, Macaranga tanarius, is an induced, indirect, defensive response elicited by jasmonic acid. Proc. Natl. Acad. Sci. USA 2001, 98, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Riefler, J.; Nowak, J.; Koschier, E.H. Components of essential oils in plant protection. Z. Arznei-Gewurzpflanzen 2009, 14, 70–76. [Google Scholar]
- Kong, W.N.; Wang, Y.; Guo, Y.F.; Chai, X.H.; Li, J.; Ma, R.Y. Behavioral effects of different attractants on adult male and female oriental fruit moths, Grapholita molesta. Pest Manag. Sci. 2020, 76, 3225–3235. [Google Scholar] [CrossRef]
- Pashalidou, F.G.; Eyman, L.; Sims, J.; Buckley, J.; Fatouros, N.E.; De Moraes, C.M.; Mescher, M.C. Plant volatiles induced by herbivore eggs prime defences and mediate shifts in the reproductive strategy of receiving plants. Ecol. Lett. 2020, 13, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Xiu, C.; Pan, H.; Liu, B.; Luo, Z.; Williams, L.; Yang, Y.; Lu, Y. Perception of and Behavioral Responses to Host Plant Volatiles for Three Adelphocoris Species. J. Chem. Ecol. 2019, 45, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Renwick, J.A.A. OvIposition behavior in lepidoptera. Annu. Rev. Entomol. 1994, 39, 377–400. [Google Scholar] [CrossRef]
- Staudt, M.; Lhoutellier, L. Volatile organic compound emission from holm oak infested by gypsy moth larvae: Evidence for distinct responses in damaged and undamaged leaves. Tree Physiol. 2007, 27, 1433–1440. [Google Scholar] [CrossRef] [Green Version]
- Arab, A.; Bento, J. Plant volatiles: New perspectives for research in Brazil. Neotrop. Entomol. 2006, 35, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alborn, H.; Turlings, T.; Jones, T.; Stenhagen, G.; Loughrin, J.; Tumlinson, J. An Elicitor of Plant Volatiles from Beet Armyworm Oral Secretion. Science 1997, 276, 945–949. [Google Scholar] [CrossRef]
- Gao, Y.; Luo, L. Research progress in tritrophic interactions among host plants, the beet armyworm Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae) and its parasitoids. Acta Entomol. Sin. 2006, 49, 333–341. [Google Scholar]
- Musser, R.O.; Hum-Musser, S.M.; Sue, M.; Eichenseer, H.; Peiffer, M.; Ervin, G.; Murphy, J.B.; Felton, G.W. Herbivory: Caterpillar saliva beats plant defences. Nature 2002, 416, 599–600. [Google Scholar] [CrossRef]
- Lange, E.S.D.; Laplanche, D.; Guo, H.; Xu, W.; Turlings, T.C.J. Spodoptera frugiperda Caterpillars Suppress Herbivore-Induced Volatile Emissions in Maize. J. Chem. Ecol. 2020, 46, 344–360. [Google Scholar] [CrossRef]
- Hare, J.D. Ecological role of volatiles produced by plants in response to damage by herbivorous insects. Annu. Rev. Entomol. 2011, 56, 161–180. [Google Scholar] [CrossRef]
- Nachappa, P.; Margolies, D.C.; Nechols, J.R.; Loughin, T. Phytoseiulus persimilis response to herbivore-induced plant volatiles as a function of mite-days. Exp. Appl. Acarol. 2006, 40, 231–239. [Google Scholar] [CrossRef]
- Miresmailli, S.; Gries, R.; Gries, G.; Zamarc, R.H.; Ismand, M.B. Population density and feeding duration of cabbage looper larvae on tomato plants alter the levels of plant volatile emissions. Pest Manag. Sci. 2011, 68, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ye, W.; Sun, M.; Xu, N.; Lou, S.; Ran, J.; Lou, Y. Three levels of defense and anti-defense responses between host plants and herbivorous insects. Chin. J. Ecol. 2015, 34, 256–262. [Google Scholar]
- Xie, M.; Chen, H.; Yi, J.; Li, K.; Zhang, J.; Su, W. Electroantennographic and behavioral responses of Anomala corpulenta to plant volatiles. Plant Prot. 2015, 41, 33–38. [Google Scholar]
- Zhang, Y.; Li, T.; Mo, J. The attractiveness of rice plant volatiles to Apanteles chilonis Munakata and Anagrus nilaparvatae Pang et Wang. Chin. J. Appl. Entomol. 2016, 53, 491–498. [Google Scholar]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Tóth, M.; Csonka, É.; Bartelt, R.J.; Cossé, A.A.; Zilkowski, B.W. Similarities in pheromonal communication of flea beetles Phyllotreta cruciferae Goeze and Ph. vittula Redtenbacher (Coleoptera, Chrysomelidae). Curr. Opin. Insect Sci. 2011, 136, 688–697. [Google Scholar]
- Liu, X.L.; Zhang, J.; Yan, Q.; Miao, C.L.; Han, W.K.; Hou, W.; Yang, K.; Hansson, B.S.; Peng, Y.C.; Guo, J.M.; et al. The molecular basis of host selection in a crucifer-specialized moth. Curr. Biol. 2020, 30, 4476–4482. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.G.; Xiao, C.; Gao, Y.L.; Dong, W.X. Behavioral responses of potato tuber moth (Phthorimaea operculella) to tobacco plant volatiles. J. Integr. Agric. 2020, 19, 325–332. [Google Scholar] [CrossRef]
- Takabayashi, J.; Dicke, M.; Posthumus, M. Volatile herbivore-induced terpenoids in plant-mite interactions: Variation caused by biotic and abiotic factors. J. Chem. Ecol. 1994, 6, 1329–1354. [Google Scholar] [CrossRef]
- Gouinguené, S.; Degen, T.; Turlings, T. Variability in herbivore-induced odour emissions among maize cultivars and their wild ancestors (teosinte). Chemoecology 2001, 1, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Turlings, T.; Erb, M. Tritrophic Interactions Mediated by Herbivore-Induced Plant Volatiles: Mechanisms, Ecological Relevance, and Application Potential. Annu. Rev. Entomol. 2018, 63, 433–452. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Linear Regression Equation | R2 | p |
|---|---|---|---|
| Green leaf volatiles | y1 = 0.0122x − 0.0576 | 0.3775 | 0.6542 |
| y2 = 0.0031x + 0.6681 | 0.1609 | 0.5782 | |
| Esters | y1 = −0.1427x + 0.6413 | 0.3946 | 0.5084 |
| y2 = 0.0148x + 0.2220 | 0.8530 | 0.0382 | |
| Aldehydes | y1 = 0.0963x + 0.1238 | 0.0587 | 0.1475 |
| y2 = −0.1305x + 0.9298 | 0.8681 | 0.0259 | |
| Terpenoids | y1 = 0.2537x + 0.1194 | 0.7103 | 0.0443 |
| y2 = −0.1545x + 0.8538 | 0.4286 | 0.3579 | |
| Ketones | y1 = −0.0035x + 0.3651 | 0.0326 | 0.2442 |
| y2 = −0.0091x + 0.8029 | 0.6722 | 0.0182 | |
| Alkanes | y1 = 0.0037x + 0.2313 | 0.0372 | 0.4786 |
| y2 = −0.0045x + 0.7619 | 0.4698 | 0.3374 |
| Number | Compound | Linear Regression Equation | R2 | p |
|---|---|---|---|---|
| 1 | E-2-hexenal | y1 = −0.1582x + 0.7733 | 0.9729 | 0.0118 |
| y2 = 0.0471x + 0.6524 | 0.9286 | 0.0195 | ||
| 2 | (Z)-3-hexen-1-ol | y1 = 0.0106x + 0.0504 | 0.3947 | 0.0931 |
| y2 = 0.0025x + 0.6880 | 0.0926 | 0.6650 | ||
| 3 | Butane, 1-isothiocyanato | y1 = 0.0308x + 0.2432 | 0.0011 | 0.4750 |
| y2 = 0.3384x + 0.6775 | 0.3567 | 0.3794 | ||
| 4 | 1-Butene, 4-isothiocyanato | y1 = −0.0148x + 0.4032 | 0.6497 | 0.0320 |
| y2 = 0.0087x + 0.6470 | 0.6842 | 0.0164 | ||
| 5 | (Z)-3-hexenyl acetate | y1 = 0.2949x + 0.0161 | 0.4216 | 0.9269 |
| y2 = −0.0011x + 0.7239 | 0.0003 | 0.9830 | ||
| 6 | Limonene | y1 = 0.1476x + 0.1863 | 0.7317 | 0.0230 |
| y2 = −0.4678x + 0.8237 | 0.6379 | 0.0123 | ||
| 7 | Allyl isothiocyanate | y1 = 0.0083x + 0.1218 | 0.2929 | 0.4685 |
| y2 = 0.0018x + 0.6892 | 0.0170 | 0.8381 | ||
| 8 | Linalool | y1 = 0.2530x + 0.1196 | 0.7105 | 0.0132 |
| y2 = −0.0775x + 0.7788 | 0.1037 | 0.7103 | ||
| 9 | Nonanal | y1 = 0.0929x + 0.1616 | 0.0626 | 0.1375 |
| y2 = −0.1147x + 0.8738 | 0.8862 | 0.0179 | ||
| 10 | Decanal | y1 = −1.870x + 0.8797 | 0.3402 | 0.5150 |
| y2 = 0.9045x + 0.4791 | 0.9739 | 0.0083 | ||
| 11 | Benzyl isothiocyanate | y1 = 0.1529x − 0.1278 | 0.1784 | 0.1890 |
| y2 = −0.0126x + 0.8033 | 0.3965 | 0.3324 | ||
| 12 | Tetradecane | y1 = −0.8443x + 0.6144 | 0.2143 | 0.9933 |
| y2 = −0.1241x + 0.7769 | 0.1323 | 0.6718 | ||
| 13 | Pentadecane | y1 = 0.0039x + 0.2349 | 0.0356 | 0.4825 |
| y2 = −0.0047x + 0.7575 | 0.4602 | 0.3447 | ||
| 14 | Hexadecane | y1 = 1.7680x − 0.1478 | 0.6716 | 0.0163 |
| y2 = −1.560x + 1.085 | 0.6310 | 0.0205 | ||
| 15 | Heptadecane | y1 = 0.0553x + 0.2272 | 0.0161 | 0.4896 |
| y2 = −0.1963x + 0.8132 | 0.6293 | 0.0228 | ||
| 16 | Octadecane | y1 = 0.4606x + 0.1868 | 0.0128 | 0.2183 |
| y2 = −0.9494x + 0.9098 | 0.5175 | 0.0263 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Y.-W.; Shi, X.-B.; Zhao, L.-C.; Yuan, G.-G.; Zhao, W.-W.; Huang, G.-H.; Chen, G. Chinese Cabbage Changes Its Release of Volatiles to Defend against Spodoptera litura. Insects 2022, 13, 73. https://doi.org/10.3390/insects13010073
Du Y-W, Shi X-B, Zhao L-C, Yuan G-G, Zhao W-W, Huang G-H, Chen G. Chinese Cabbage Changes Its Release of Volatiles to Defend against Spodoptera litura. Insects. 2022; 13(1):73. https://doi.org/10.3390/insects13010073
Chicago/Turabian StyleDu, Yuan-Wen, Xiao-Bin Shi, Lin-Chao Zhao, Ge-Ge Yuan, Wei-Wei Zhao, Guo-Hua Huang, and Gong Chen. 2022. "Chinese Cabbage Changes Its Release of Volatiles to Defend against Spodoptera litura" Insects 13, no. 1: 73. https://doi.org/10.3390/insects13010073
APA StyleDu, Y. -W., Shi, X. -B., Zhao, L. -C., Yuan, G. -G., Zhao, W. -W., Huang, G. -H., & Chen, G. (2022). Chinese Cabbage Changes Its Release of Volatiles to Defend against Spodoptera litura. Insects, 13(1), 73. https://doi.org/10.3390/insects13010073