
Predicting Climate Change Effects on the Potential Distribution of Two Invasive Cryptic Species of the Bemisia tabaci Species Complex in China

 , ,
, , 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
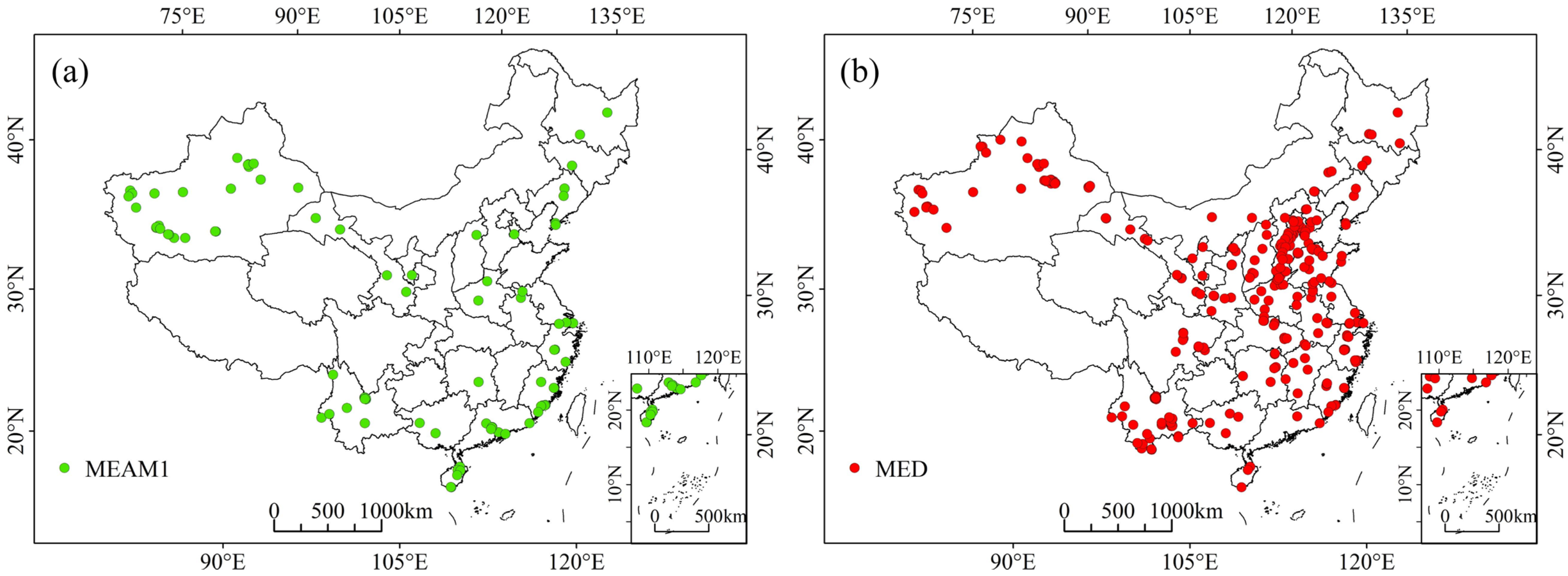
2.1. Occurrence Data
2.2. Environmental Variables
2.3. Model Development
2.4. Model Evaluation
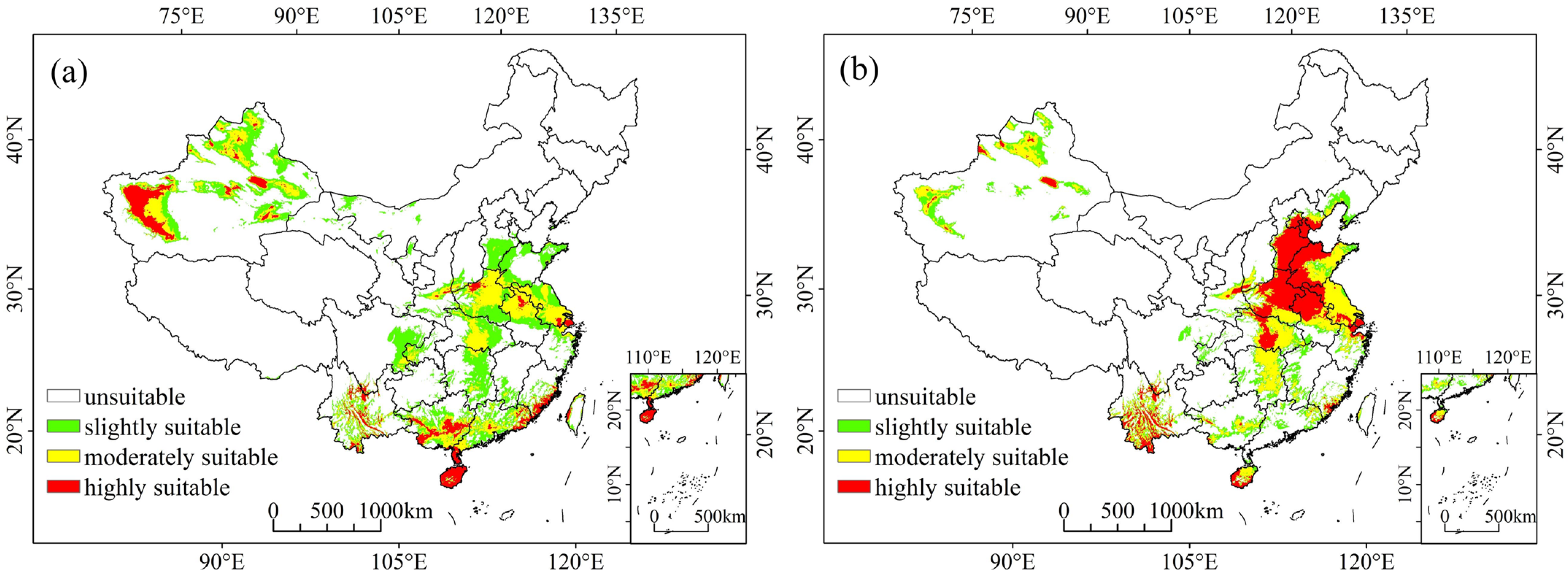
2.5. Habitat Suitability Classification
2.6. Distribution Change Estimation
3. Results
3.1. Predicted Current Potential Distribution
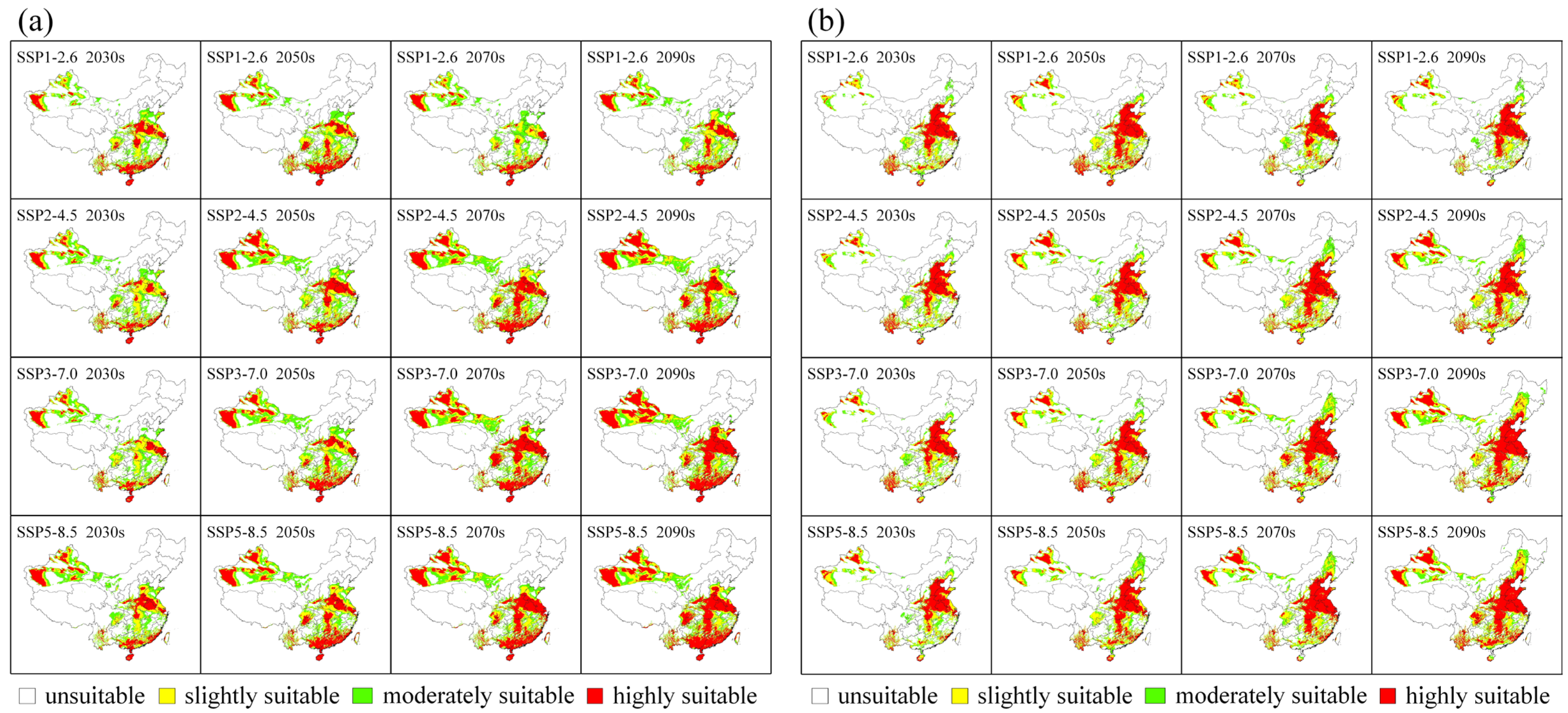
3.2. Predicted Future Potential Distribution
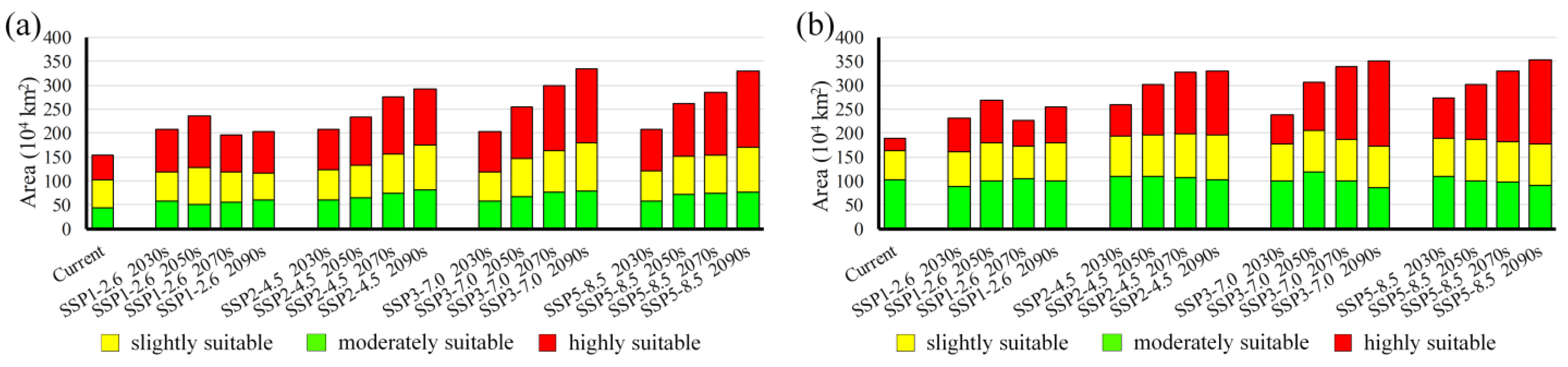
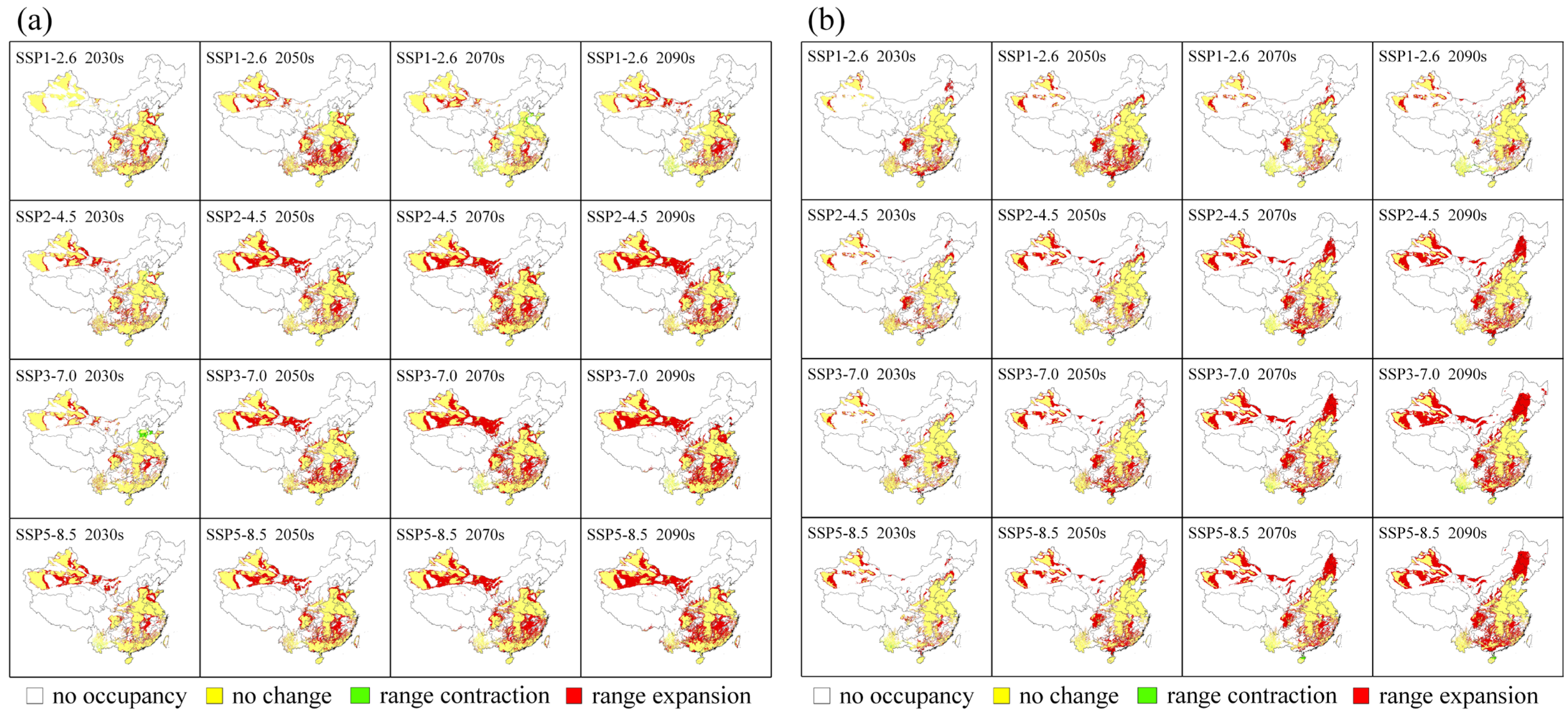
3.3. Habitat Changes under Future Climate Scenarios
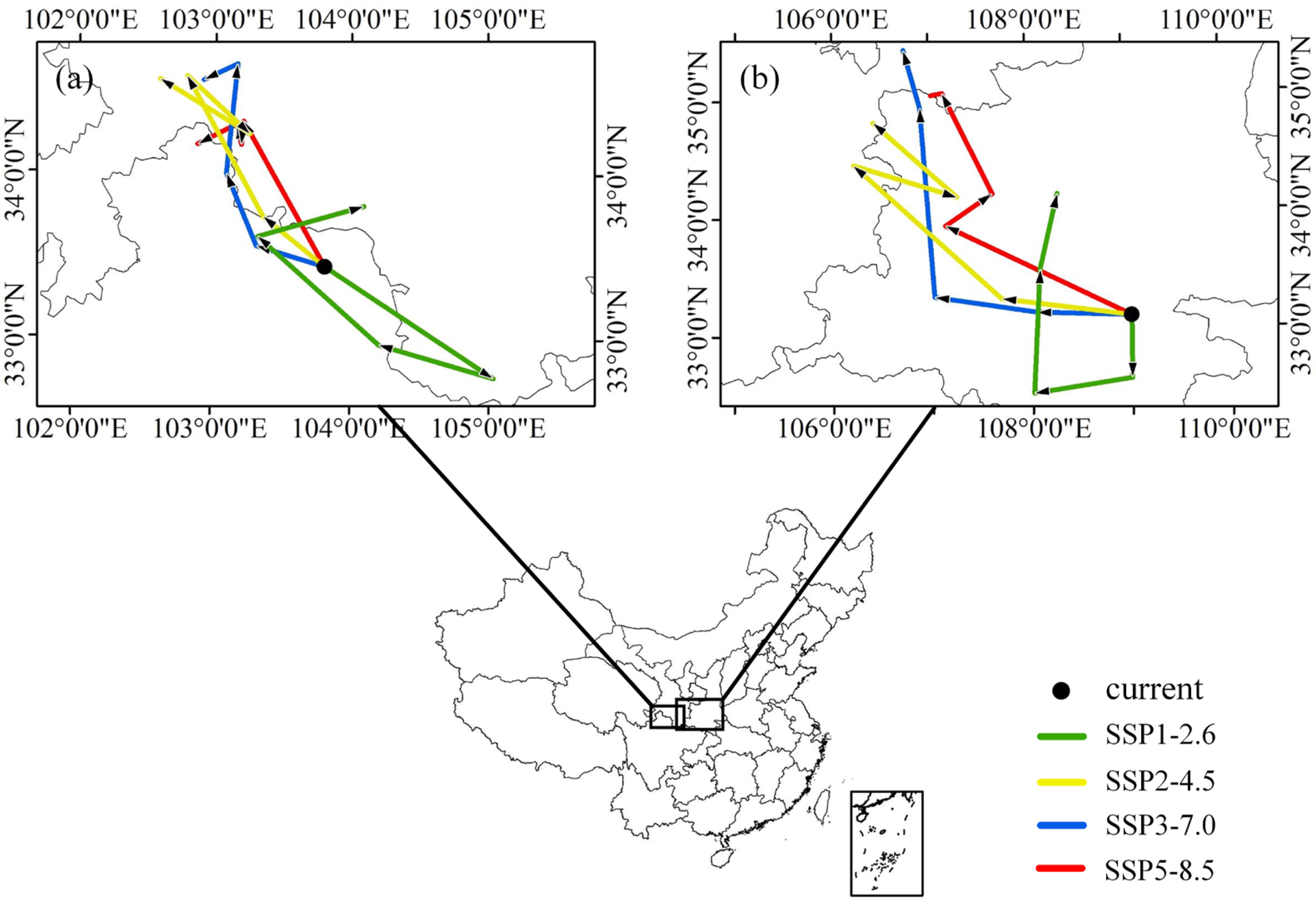
3.4. Centroid Shifts between Current and Future Distribution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Early, R.; Sax, D.F. Climatic niche shifts between species’ native and naturalized ranges raise concern for ecological forecasts during invasions and climate change. Glob. Ecol. Biogeogr. 2014, 23, 1356–1365. [Google Scholar] [CrossRef]
- Wainright, C.A.; Muhlfeld, C.C.; Elser, J.J.; Bourret, S.L.; Devlin, S.P. Species invasion progressively disrupts the trophic structure of native food webs. Proc. Natl. Acad. Sci. USA 2021, 118, e2102179118. [Google Scholar] [CrossRef] [PubMed]
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; van Kleunen, M.; Kuhn, I.; et al. Projecting the continental accumulation of alien species through to 2050. Glob. Chang. Biol. 2021, 27, 970–982. [Google Scholar] [CrossRef] [PubMed]
- Diagne, C.; Leroy, B.; Vaissiere, A.C.; Gozlan, R.E.; Roiz, D.; Jaric, I.; Salles, J.M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef]
- UNDP. The Sustainable Development Goals. Available online: https://www.undp.org/sustainable-development-goals (accessed on 5 June 2021).
- Cornelissen, B.; Neumann, P.; Schweiger, O. Global warming promotes biological invasion of a honey bee pest. Glob. Chang. Biol. 2019, 25, 3642–3655. [Google Scholar] [CrossRef] [Green Version]
- Saranya, K.R.L.; Lakshmi, T.V.; Reddy, C.S. Predicting the potential sites of Chromolaena odorata and Lantana camara in forest landscape of Eastern Ghats using habitat suitability models. Ecol. Inform. 2021, 66, 101455. [Google Scholar] [CrossRef]
- Zettlemoyer, M.A.; Schultheis, E.H.; Lau, J.A. Phenology in a warming world: Differences between native and non-native plant species. Ecol. Lett. 2019, 22, 1253–1263. [Google Scholar] [CrossRef]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change for invasive species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef]
- Guan, B.C.; Guo, H.J.; Chen, S.S.; Li, D.M.; Liu, X.; Gong, X.; Ge, G. Shifting ranges of eleven invasive alien plants in China in the face of climate change. Ecol. Inform. 2020, 55, 101024. [Google Scholar] [CrossRef]
- Wan, F.H.; Yang, N.W. Invasion and Management of Agricultural Alien Insects in China. Annu. Rev. Entomol. 2016, 61, 77–98. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A Statement of Species Status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Gautam, S. The Role of Bemisia tabaci in the Transmission of Vegetable Viruses in the Farmscape of Georgia. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2019. [Google Scholar]
- Ferreira Rodrigues, R.H.; Silva, L.B.; Silva, M.C.F.; Lopes, J.W.B.; Araujo Lima, E.; Sobreira Barbosa, R.; Oliveira Almeida, L.F. Population fluctuation and distribution of bemisia tabaci MEAM1 (Hemiptera: Aleyrodidae) in soybean crops. Front. Agron. 2022, 4, 958498. [Google Scholar] [CrossRef]
- Macfadyen, S.; Tay, W.T.; Hulthen, A.D.; Paull, C.; Kalyebi, A.; Jacomb, F.; Parry, H.; Sseruwagi, P.; Seguni, Z.; Abu Omongo, C.; et al. Landscape factors and how they influence whitefly pests in cassava fields across East Africa. Landsc. Ecol. 2021, 36, 45–67. [Google Scholar] [CrossRef]
- Chu, D.; Zhang, Y.J.; Brown, J.K.; Cong, B.; Xu, B.Y.; Wu, Q.J.; Zhu, G.R. The introduction of the exotic Q biotype of Bemisia tabaci from the Mediterranean region into China on ornamental crops. Fla. Entomol. 2006, 89, 168–174. [Google Scholar] [CrossRef]
- Luo, C.; Yao, Y.; Wang, R.; Yan, F.; Hu, D.; Zhang, Z. The use of mitochondrial cytochrome oxidase I (mt CO I) gene sequences for the identification of biotypes of Bemisia tabaci (Gennadius) in China. Acta Entomol. Sin. 2002, 45, 759–763. [Google Scholar]
- Chu, D.; Wan, F.H.; Zhang, Y.J.; Brown, J.K. Change in the Biotype Composition of Bemisia tabaci in Shandong Province of China From 2005 to 2008. Environ. Entomol. 2010, 39, 1028–1036. [Google Scholar] [CrossRef]
- Hu, J.; De Barro, P.; Zhao, H.; Wang, J.; Nardi, F.; Liu, S.S. An extensive field survey combined with a phylogenetic analysis reveals rapid and widespread invasion of two alien whiteflies in China. PLoS ONE 2011, 6, e16061. [Google Scholar] [CrossRef]
- Pan, H.P.; Chu, D.; Ge, D.Q.; Wang, S.L.; Wu, Q.J.; Xie, W.; Jiao, X.G.; Liu, B.M.; Yang, X.; Yang, N.; et al. Further Spread of and Domination by Bemisia tabaci (Hemiptera: Aleyrodidae) Biotype Q on Field Crops in China. J. Econ. Entomol. 2011, 104, 978–985. [Google Scholar] [CrossRef]
- Rao, Q.; Luo, C.; Zhang, H.; Guo, X.; Devine, G.J. Distribution and dynamics of Bemisia tabaci invasive biotypes in central China. Bull. Entomol. Res. 2011, 101, 81–88. [Google Scholar] [CrossRef]
- Guo, C.L.; Zhu, Y.Z.; Zhang, Y.J.; Keller, M.A.; Liu, T.X.; Chu, D. Invasion Biology and Management of Sweetpotato Whitefly (Hemiptera: Aleyrodidae) in China. J. Integr. Pest Manag. 2021, 12, pmaa024. [Google Scholar] [CrossRef]
- Liu, B.M.; Yan, F.M.; Chu, D.; Pan, H.P.; Jiao, X.G.; Xie, W.; Wu, Q.J.; Wang, S.L.; Xu, B.Y.; Zhou, X.G.; et al. Difference in Feeding Behaviors of Two Invasive Whiteflies on Host Plants with Different Suitability: Implication for Competitive Displacement. Int. J. Biol. Sci. 2012, 8, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.B.; Li, J.; Liu, Y.Q.; Crowder, D.W.; Liu, S.S. Effects of reproductive interference on the competitive displacement between two invasive whiteflies. Bull. Entomol. Res. 2014, 104, 334–346. [Google Scholar] [CrossRef] [Green Version]
- Horowitz, A.R.; Ghanim, M.; Roditakis, E.; Nauen, R.; Ishaaya, I. Insecticide resistance and its management in Bemisia tabaci species. J. Pest Sci. 2020, 93, 893–910. [Google Scholar] [CrossRef]
- Xiao, N.; Pan, L.L.; Zhang, C.R.; Shan, H.W.; Liu, S.S. Differential tolerance capacity to unfavourable low and high temperatures between two invasive whiteflies. Sci. Rep. 2016, 6, 24306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.T.; Cai, L.; Shen, Y.; Xu, L.L.; Du, Y.Z. Competitive Displacement between Bemisia tabaci MEAM1 and MED and Evidence for Multiple Invasions of MED. Insects 2020, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.T.; Lin, C.T.; Wang, Y.Z.; Zhang, Y.B.; Ji, L.Q. Ecological niche complexity of invasive and native cryptic species of the Bemisia tabaci species complex in China. J. Pest Sci. 2022, 95, 1245–1259. [Google Scholar] [CrossRef]
- Gautam, S.; Mugerwa, H.; Buck, J.W.; Dutta, B.; Coolong, T.; Adkins, S.; Srinivasan, R. Differential Transmission of Old and New World Begomoviruses by Middle East-Asia Minor 1 (MEAM1) and Mediterranean (MED) Cryptic Species of Bemisia tabaci. Viruses 2022, 14, 1104. [Google Scholar] [CrossRef]
- Mugerwa, H.; Gautam, S.; Catto, M.A.; Dutta, B.; Brown, J.K.; Adkins, S.; Srinivasan, R. Differential Transcriptional Responses in Two Old World Bemisia tabaci Cryptic Species Post Acquisition of Old and New World Begomoviruses. Cells 2022, 11, 2060. [Google Scholar] [CrossRef]
- Shi, X.B.; Tang, X.; Zhang, X.; Zhang, D.Y.; Li, F.; Yan, F.; Zhang, Y.J.; Zhou, X.G.; Liu, Y. Transmission Efficiency, Preference and Behavior of Bemisia tabaci MEAM1 and MED under the Influence of Tomato Chlorosis Virus. Front. Plant Sci. 2018, 8, 2271. [Google Scholar] [CrossRef] [Green Version]
- Chi, Y.; Pan, L.L.; Bouvaine, S.; Fan, Y.Y.; Liu, Y.Q.; Liu, S.S.; Seal, S.; Wang, X.W. Differential transmission of Sri Lankan cassava mosaic virus by three cryptic species of the whitefly Bemisia tabaci complex. Virology 2020, 540, 141–149. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Yang, N.W.; Sun, L.Y.; Wan, F.H. Host instar suitability in two invasive whiteflies for the naturally occurring parasitoid Eretmocerus hayati in China. J. Pest Sci. 2015, 88, 225–234. [Google Scholar] [CrossRef]
- Kumar, S.; Neven, L.G.; Yee, W.L. Evaluating correlative and mechanistic niche models for assessing the risk of pest establishment. Ecosphere 2014, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudik, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Mudereri, B.T.; Kimathi, E.; Chitata, T.; Moshobane, M.C.; Abdel-Rahman, E.M. Landscape-scale biogeographic distribution analysis of the whitefly, Bemisia tabaci (Gennadius, 1889) in Kenya. Int. J. Trop. Insect Sci. 2021, 41, 1585–1599. [Google Scholar] [CrossRef]
- Paredes-Montero, J.R.; Ibarra, M.A.; Arias-Zambrano, M.; Peralta, E.L.; Brown, J.K. Phylo-biogeographical distribution of whitefly Bemisia tabaci (Insecta: Aleyrodidae) mitotypes in Ecuador. Ecosphere 2020, 11, e03154. [Google Scholar] [CrossRef]
- Ramos, R.S.; Kumar, L.; Shabani, F.; Picanco, M.C. Mapping global risk levels of Bemisia tabaci in areas of suitability for open field tomato cultivation under current and future climates. PLoS ONE 2018, 13, 20. [Google Scholar] [CrossRef]
- Prabhulinga, T.; Rameash, K.; Madhu, T.N.; Vivek, S.; Suke, R. Maximum entropy modelling for predicting the potential distribution of cotton whitefly Bemisia tabaci (Gennadius) in North India. J. Entomol. Zool. Stud. 2017, 5, 1002–1006. [Google Scholar]
- Ren, S.; Qiu, B.; Ge, F.; Zhang, Y.; Du, Y.; Chen, X.; Guo, J.; Lin, K.; Peng, Z.; Yao, S.; et al. Research progress of the monitoring, forecast and sustainable management of whitefly pests in China. Chin. J. Appl. Entomol. 2011, 48, 7–15. [Google Scholar]
- Zhao, J.; Li, Z.; Zhang, H.; Chen, H.; Weng, Q. Prediction of potential distribution of tobacco whitefly Bemisia tabaci in China based on MaxEnt niche model and GIS. Acta Phytophylacica Sin. 2019, 46, 1292–1300. [Google Scholar]
- Hu, J.; Jiang, Z.L.; Nardi, F.; Liu, Y.Y.; Luo, X.R.; Li, H.X.; Zhang, Z.K. Members of Bemisia tabaci (Hemiptera: Aleyrodidae) Cryptic Species and the Status of Two Invasive Alien Species in the Yunnan Province (China). J. Insect Sci. 2014, 14, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Zhang, X.; Jiang, Z.; Zhang, F.; Liu, Y.; Li, Z.; Zhang, Z. New putative cryptic species detection and genetic network analysis of Bemisia tabaci (Hempitera: Aleyrodidae) in China based on mitochondrial COI sequences. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2018, 29, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.Z.; Fu, K.Y.; Guo, W.C.; Jiang, W.H.; Ahmat, T.; Ding, X.H.; He, J.; Wang, X.W. CAP Analysis of the Distribution of the Introduced Bemisia tabaci (Hemiptera: Aleyrodidae) Species Complex in Xinjiang, China and the Southerly Expansion of the Mediterranean Species. J. Insect Sci. 2021, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Thuiller, W.; Albert, C.; Araujo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgely, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, B.C.; Tebaldi, C.; van Vuuren, D.P.; Eyring, V.; Friedlingstein, P.; Hurtt, G.; Knutti, R.; Kriegler, E.; Lamarque, J.F.; Lowe, J.; et al. The Scenario Model Intercomparison Project (ScenarioMIP) for CMIP6. Geosci. Model Dev. 2016, 9, 3461–3482. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.W.; Lu, Y.X.; Fang, Y.J.; Xin, X.G.; Li, L.; Li, W.P.; Jie, W.H.; Zhang, J.; Liu, Y.M.; Zhang, L.; et al. The Beijing Climate Center Climate System Model (BCC-CSM): The main progress from CMIP5 to CMIP6. Geosci. Model Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef] [Green Version]
- Owens, H.L.; Campbell, L.P.; Dornak, L.L.; Saupe, E.E.; Barve, N.; Soberon, J.; Ingenloff, K.; Lira-Noriega, A.; Hensz, C.M.; Myers, C.E.; et al. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol. Model. 2013, 263, 10–18. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MAXENT ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Papes, M.; Soberon, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Ramos, R.S.; Kumar, L.; Shabani, F.; Picanco, M.C. Risk of spread of tomato yellow leaf curl virus (TYLCV) in tomato crops under various climate change scenarios. Agric. Syst. 2019, 173, 524–535. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Beaury, E.M.; Fusco, E.J.; Jackson, M.R.; Laginhas, B.B.; Morelli, T.L.; Allen, J.M.; Pasquarella, V.J.; Bradley, B.A. Incorporating climate change into invasive species management: Insights from managers. Biol. Invasions 2020, 22, 233–252. [Google Scholar] [CrossRef]
- Mainali, K.P.; Warren, D.L.; Dhileepan, K.; McConnachie, A.; Strathie, L.; Hassan, G.; Karki, D.; Shrestha, B.B.; Parmesan, C. Projecting future expansion of invasive species: Comparing and improving methodologies for species distribution modeling. Glob. Chang. Biol. 2015, 21, 4464–4480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uden, D.R.; Allen, C.R.; Angeler, D.G.; Corral, L.; Fricke, K.A. Adaptive invasive species distribution models: A framework for modeling incipient invasions. Biol. Invasions 2015, 17, 2831–2850. [Google Scholar] [CrossRef]
- Zhou, M.; Zhou, G.; Lu, X.; Zhou, L.; Ji, Y. Projection of temperature and precipitation changes over China under global warming of 1.5 and 2 °C. Acta Meteorol. Sin. 2019, 77, 728–744. [Google Scholar]
- Gilioli, G.; Pasquali, S.; Parisi, S.; Winter, S. Modelling the potential distribution of Bemisia tabaci in Europe in light of the climate change scenario. Pest Manag. Sci. 2014, 70, 1611–1623. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Bai Run, E.; Zhai, Q.; Wang, B.; Yan, F. New record of the Bemisia tabaci Q biotype in Tibet, China. Chin. J. Appl. Entomol. 2016, 53, 213–217. [Google Scholar]
- Xue, Y.T. Geographical Distribution, Genetic Structure of Bemisia Tabaci (Hemiptera:Aleyrodidae) in China, and Diversity of Its Secondary Endosymbionts. Master’s Thesis, Southwest University, Chongqing, China, 2018. [Google Scholar]
- Gautam, S.; Crossley, M.S.; Dutta, B.; Coolong, T.; Simmons, A.M.; da Silva, A.; Snyder, W.E.; Srinivasan, R. Low Genetic Variability in Bemisia tabaci MEAM1 Populations within Farmscapes of Georgia, USA. Insects 2020, 11, 834. [Google Scholar] [CrossRef]
- Gu, C.J.; Tu, Y.L.; Liu, L.S.; Bo, W.; Zhang, Y.L.; Yu, H.B.; Wang, X.L.; Zhuoga, Y.J.; Zhang, B.H.; Cui, B.H. Predicting the potential global distribution of Ageratina adenophora under current and future climate change scenarios. Ecol. Evol. 2021, 11, 12092–12113. [Google Scholar] [CrossRef]
- Ramasamy, M.; Das, B.; Ramesh, R. Predicting climate change impacts on potential worldwide distribution of fall armyworm based on CMIP6 projections. J. Pest Sci. 2022, 95, 841–854. [Google Scholar] [CrossRef]
- Peng, L.Q.; Tang, M.; Liao, J.H.; Qing, H.F.; Zhao, Z.K.; Pike, D.A.; Chen, W. Potential effects of climate change on the distribution of invasive bullfrogs Lithobates catesbeianus in China. Acta Herpetol. 2020, 15, 87–94. [Google Scholar] [CrossRef]
- Thurman, L.L.; Stein, B.A.; Beever, E.A.; Foden, W.; Geange, S.R.; Green, N.; Gross, J.E.; Lawrence, D.J.; LeDee, O.; Olden, J.D.; et al. Persist in place or shift in space? Evaluating the adaptive capacity of species to climate change. Front. Ecol. Environ. 2020, 18, 520–528. [Google Scholar] [CrossRef]
- Ma, G.; Ma, C.S. Potential distribution of invasive crop pests under climate change: Incorporating mitigation responses of insects into models. Curr. Opin. Insect Sci. 2022, 49, 15–21. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Type | Variable Code | Description |
|---|---|---|
| Bioclimatic variables | bio1 | Annual mean temperature |
| bio2 | Mean diurnal range (mean of monthly (max temp.−min temp.)) | |
| bio3 | Isothermality (bio2/bio7) (×100) | |
| bio4 | Temperature seasonality (standard deviation ×100) | |
| bio5 | Maximum temperature of the warmest month | |
| bio6 | Minimum temperature of the coldest month | |
| bio7 | Temperature annual range (bio5−bio6) | |
| bio8 | Mean temperature of the wettest quarter | |
| bio9 | Mean temperature of the driest quarter | |
| bio10 | Mean temperature of the warmest quarter | |
| bio11 | Mean temperature of the coldest quarter | |
| bio12 | Annual precipitation | |
| bio13 | Precipitation of the wettest month | |
| bio14 | Precipitation of the driest month | |
| bio15 | Precipitation seasonality (coefficient of variation) | |
| bio16 | Precipitation of the wettest quarter | |
| bio17 | Precipitation of the driest quarter | |
| bio18 | Precipitation of the warmest quarter | |
| bio19 | Precipitation of the coldest quarter | |
| Elevation | elev | Ground height above sea level |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, Y.; Lin, C.; Wang, Y.; Liu, W.; Wan, F.; Zhang, Y.; Ji, L. Predicting Climate Change Effects on the Potential Distribution of Two Invasive Cryptic Species of the Bemisia tabaci Species Complex in China. Insects 2022, 13, 1081. https://doi.org/10.3390/insects13121081
Xue Y, Lin C, Wang Y, Liu W, Wan F, Zhang Y, Ji L. Predicting Climate Change Effects on the Potential Distribution of Two Invasive Cryptic Species of the Bemisia tabaci Species Complex in China. Insects. 2022; 13(12):1081. https://doi.org/10.3390/insects13121081
Chicago/Turabian StyleXue, Yantao, Congtian Lin, Yaozhuo Wang, Wanxue Liu, Fanghao Wan, Yibo Zhang, and Liqiang Ji. 2022. "Predicting Climate Change Effects on the Potential Distribution of Two Invasive Cryptic Species of the Bemisia tabaci Species Complex in China" Insects 13, no. 12: 1081. https://doi.org/10.3390/insects13121081
APA StyleXue, Y., Lin, C., Wang, Y., Liu, W., Wan, F., Zhang, Y., & Ji, L. (2022). Predicting Climate Change Effects on the Potential Distribution of Two Invasive Cryptic Species of the Bemisia tabaci Species Complex in China. Insects, 13(12), 1081. https://doi.org/10.3390/insects13121081