



Attraction of Insects to Ornamental Lighting Used on Cultural Heritage Buildings: A Case Study in an Urban Area

 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Location and Sampling Set-Up
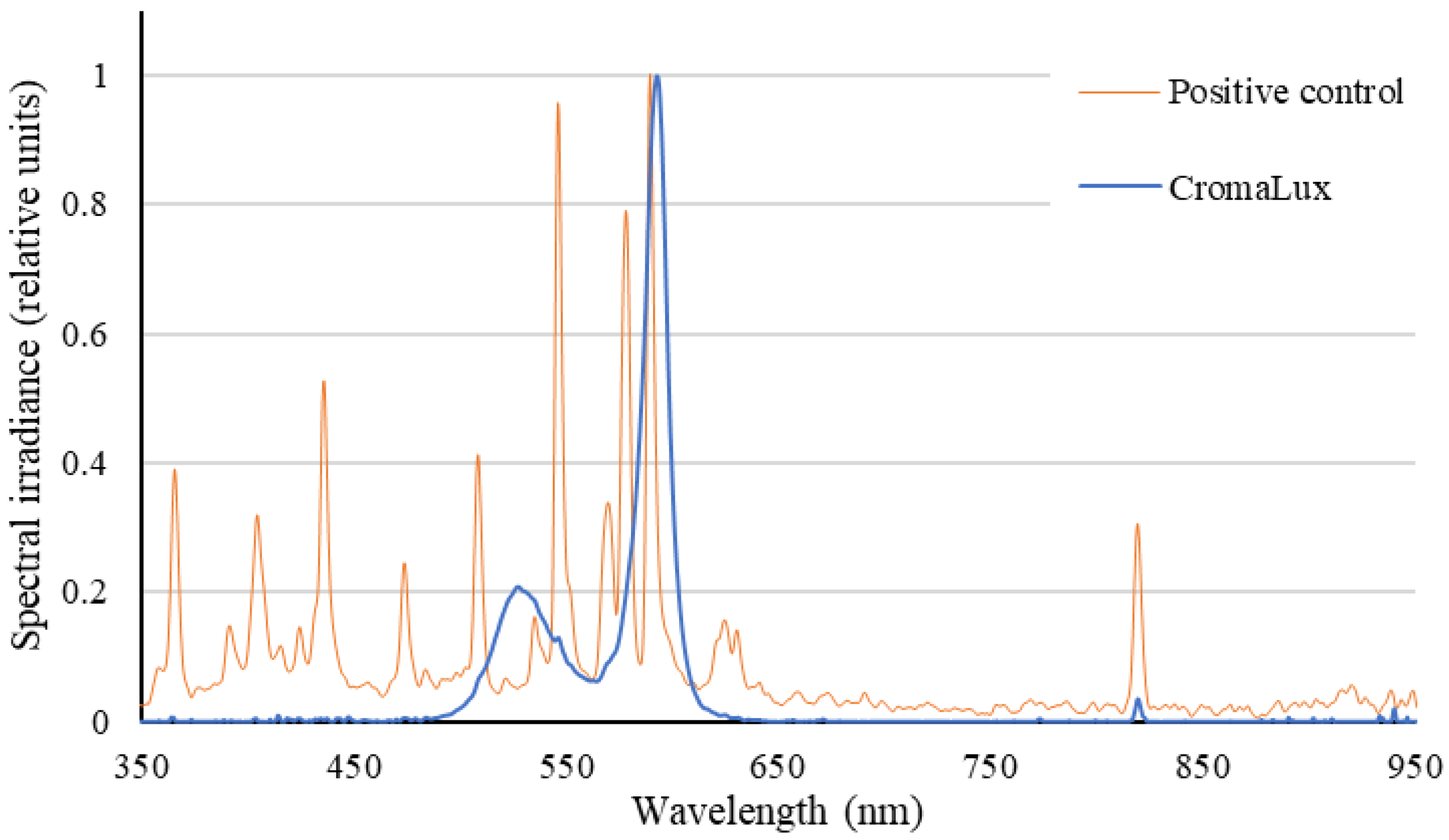
2.2. Artificial Lighting and Environmental Conditions
2.3. Insect Collection and Identification
2.4. Biodiversity and Statistical Analysis
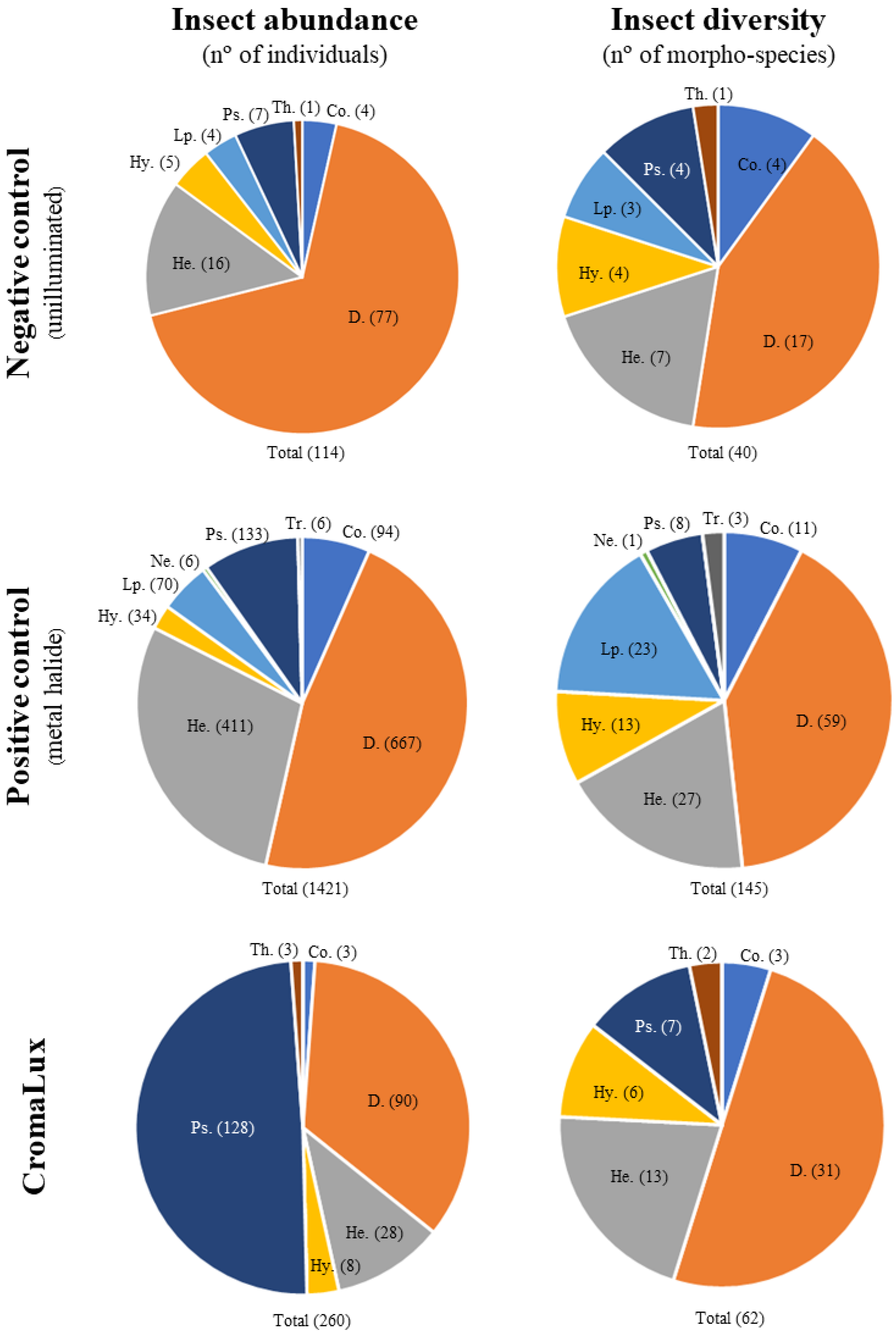
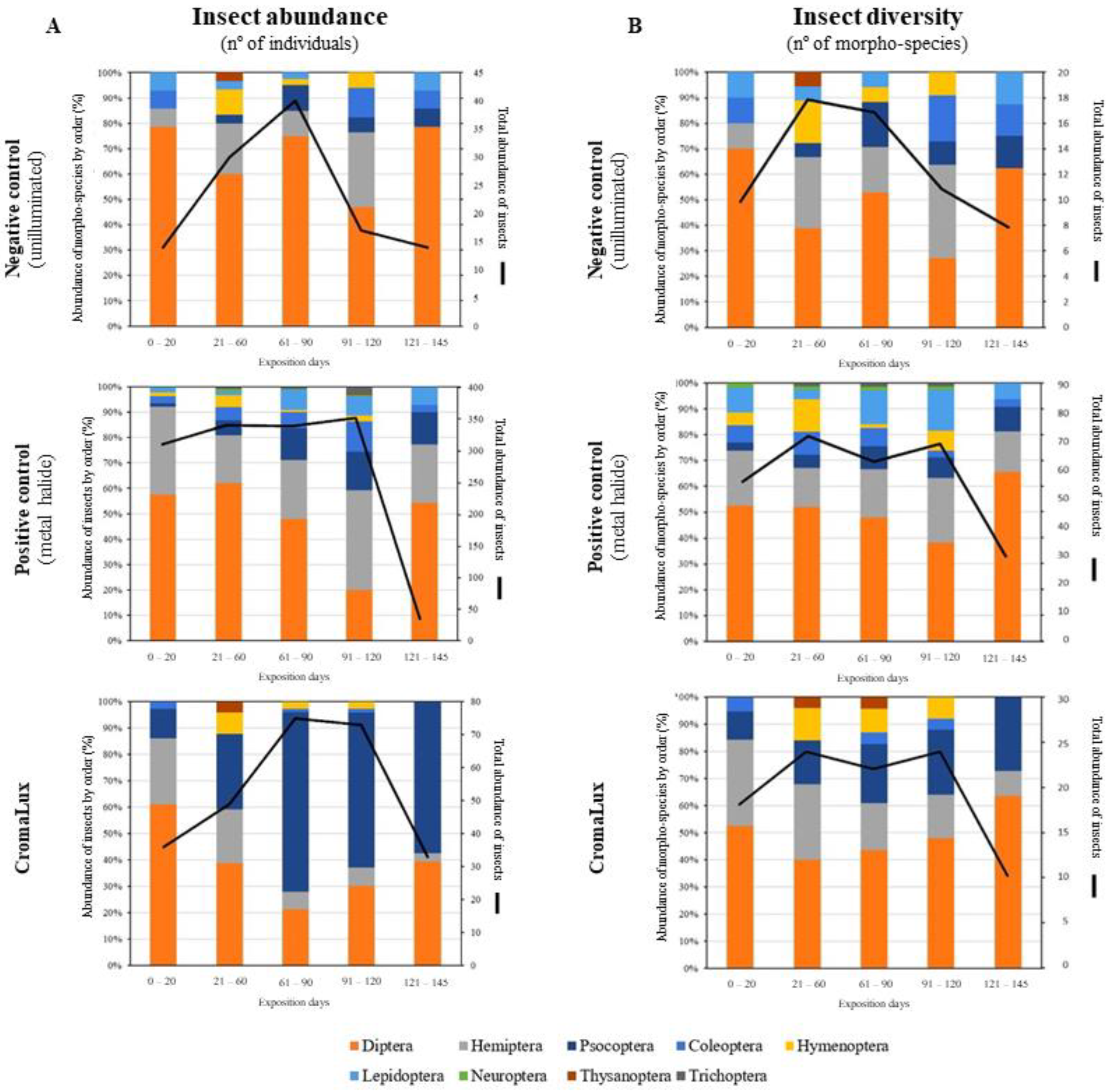
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazur, D.; Wachta, H.; Leśko, K. Research of cohesion principle in illuminations of monumental Objects. In Analysis and Simulation of Electrical and Computer Systems, 1st ed.; Mazur, D., Golebiowski, M., Korkosz, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 395–406. [Google Scholar]
- Zielinska-Dabkowska, K.M.; Xavia, K. Historic Urban Settings, LED Illumination and its Impact on Nighttime Perception, Visual Appearance, and Cultural Heritage Identity. In Proceedings of the 5th International Multidisciplinary Scientific Conference on Social Sciences and Arts, Florence, Italy, 23–26 October 2018; SGEM: Sofia, Bulgaria, 2018. [Google Scholar]
- Falchi, F.; Cinzano, P.; Duriscoe, D.; Kyba, C.C.; Elvidge, C.D.; Baugh, K.; Portnov, B.; Rybnikova, N.; Furgoni, R. The new world atlas of artificial night sky brightness. Sci. Adv. 2016, 2, e1600377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hölker, F.; Wolter, C.; Perkin, E.K.; Tockner, K. Light pollution as a biodiversity threat. Trends Ecol. Evol. 2010, 25, 681–682. [Google Scholar] [CrossRef]
- Sanders, D.; Gaston, K.J. How ecological communities respond to artificial light at night. J. Exp. Zool. 2018, 329, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Touzot, M.; Lengagne, T.; Secondi, J.; Desouhant, E.; Thery, M.; Dumet, A.; Duchamp, C.; Mondy, N. Artificial light at night alters the sexual behaviour and fertilisation success of the common toad. Environ. Pollut. 2020, 259, 113883. [Google Scholar] [CrossRef]
- Kyba CC, M.; Mohar, A.; Pintar, G.; Stare, J. Reducing the environmental footprint of church lighting: Matching facade shape and lowering luminance with the EcoSky LED. Int. J. Sustain. Light. 2018, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kyba, C.C.M.; Kuester, T.; de Miguel, A.S.; Baugh, K.; Jechow, A.; Hölker, F.; Bennie, J.; Elvidge, C.D.; Gaston, K.J.; Guanter, L. Artificially lit surface of Earth at night increasing in radiance and extent. Sci. Adv. 2017, 3, e1701528. [Google Scholar] [CrossRef] [Green Version]
- Mohar, A.; Zagmajster, M.; Verovnik, R.; Skaberne, B. Nature-Friendlier Lighting of Objects of Cultural Heritage (Churches). Dark-Sky Slovenia, Liubliana. 2014. Available online: https://www.anl.bayern.de/publikationen/anliegen/additional_data/an37200notizen_2015_kulturdenkmaeler_life_bericht_engl.pdf (accessed on 8 November 2022).
- Jechow, A.; Ribas, S.J.; Domingo, R.C.; Hölker, F.; Kolláth, Z.; Kyba, C. Tracking the dynamics of skyglow with differential photometry using a digital camera with fisheye lens. J. Quant. Spectrosc. Radiat. Transf. 2018, 209, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Owens, A.C.; Cochard, P.; Durrant, J.; Farnworth, B.; Perkin, E.K.; Seymoure, B. Light pollution is a driver of insect declines. Biol. Conserv. 2020, 241, 108259. [Google Scholar] [CrossRef]
- Gaston, K.J.; Visser, M.E.; Hölker, F. The biological impacts of artificial light at night: The research challenge. Philos. Trans. R. Soc. 2015, 370, 20140133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, A.C.; Lewis, S.M. The impact of artificial light at night on nocturnal insects: A review and synthesis. Ecol. Evol. 2018, 8, 11337–11358. [Google Scholar] [CrossRef] [PubMed]
- van Grunsven, R.H.; Becker, J.; Peter, S.; Heller, S.; Hölker, F. Long-term comparison of attraction of flying insects to streetlights after the transition from traditional light sources to light-emitting diodes in urban and peri-urban settings. Sustainability 2019, 11, 6198. [Google Scholar] [CrossRef] [Green Version]
- Boyes, D.H.; Evans, D.M.; Fox, R.; Parsons, M.S.; Pocock, M.J. Street lighting has detrimental impacts on local insect populations. Sci. Adv. 2021, 7, eabi8322. [Google Scholar] [CrossRef] [PubMed]
- Acharya, L.; Fenton, M.B. Bat attacks and moth defensive behavior around street lights. Can. J. Zool. 1999, 77, 27–33. [Google Scholar] [CrossRef]
- Boyes, D.H.; Evans, D.M.; Fox, R.; Parsons, M.S.; Pocock, M.J. Is light pollution driving moth population declines? A review of causal mechanisms across the life cycle. Insect Conserv. Divers. 2021, 14, 167–187. [Google Scholar] [CrossRef]
- Deichmann, J.L.; Ampudia Gatty, C.; Andía Navarro, J.M.; Alonso, A.; Linares-Palomino, R.; Longcore, T. Reducing the blue spectrum of artificial light at night minimises insect attraction in a tropical lowland forest. Insect Conserv. Divers. 2021, 14, 247–259. [Google Scholar] [CrossRef]
- Harris, J.B. Electric lamps, past and present. J. Eng. Educ. 1993, 2, 161–170. [Google Scholar] [CrossRef]
- Davies, T.W.; Bennie, J.; Inger, R.; De Ibarra, N.H.; Gaston, K.J. Artificial light pollution: Are shifting spectral signatures changing the balance of species interactions? Glob. Chang. Biol. 2013, 19, 1417–1423. [Google Scholar] [CrossRef] [Green Version]
- Gendre, M. Two Centuries of Electric Light Source Innovations. 2003. Available online: https://www.einlightred.tue.nl/lightsources/history/light_history.pdf (accessed on 8 November 2022).
- Rea, M.S.; Bullough, J.D.; Akashi, Y. Several views of metal halide and high-pressure sodium lighting for outdoor applications. Light. Res. Technol. 2009, 41, 297–320. [Google Scholar] [CrossRef]
- Verovnik, R.; Fišer, Ž.; Zakšek, V. How to reduce the impact of artificial lighting on moths: A case study on cultural heritage sites in Slovenia. J. Nat. Conserv. 2015, 28, 105–111. [Google Scholar] [CrossRef]
- Otieno, J.A.; Weller, J.; Poehling, H.M. Efficacy of LED-enhanced blue sticky traps combined with the synthetic lure Lurem-TR for trapping of western fower thrips (Frankliniella occidentalis). J. Pest. Sci. 2018, 91, 1301–1314. [Google Scholar] [CrossRef]
- Shi, L.; He, H.; Yang, G.; Huang, H.; Vasseur, L.; You, M. Are yellow sticky cards and light traps effective on tea green leafhoppers and their predators in Chinese Tea plantations? Insects 2021, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Shreeve, T. The effect of weather on the life cycle of the speckled wood butterfly Pararge aegeria. Ecol. Entomol. 1986, 11, 325–332. [Google Scholar] [CrossRef]
- Scott JEpstein, M. Factors affecting phenology in a temperate insect community. Am. Midl. Nat. 1987, 103–118. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Wilson, R. Intra-and interspecific variation in the responses of insect phenology to climate. J. Anim. Ecol. 2021, 90, 248–259. [Google Scholar] [CrossRef]
- Barrientos, J.A. (Ed.) Curso Práctico de Entomología; Univ. Autònoma de Barcelona: Barcelona, Spain, 2004; Volume 41. [Google Scholar]
- Malicky, H. Atlas of European Trichoptera, 2nd ed.; Springer: Dordrecht, The Netherlands, 2004; p. 385. [Google Scholar]
- Oosterbroek, P. The European Families of the Diptera: Identification-Diagnosis-Biology; KNNV Publishing: Utrecht, The Netherlands, 2006; p. 205. [Google Scholar]
- Leraut, P. Insectos de España y Europa; Lynx Edicions: Barcelona, Spain, 2007; p. 527. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; p. 144. [Google Scholar]
- Hutcheson, K. A test for comparing diversities based on the Shannon formula. J. Theor. Biol. 1970, 29, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Conover, W.J.; Iman, R.L. On some alternative procedures using ranks for the analysis of experimental designs. Commun. Stat.-Theory Methods 1976, 5, 1349–1368. [Google Scholar] [CrossRef]
- Conover, W. Practical Nonparametric Statistics; John Wiley & Sons: New York, NY, USA, 1980; p. 493. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- van der Kooi, C.J.; Stavenga, D.G.; Arikawa, K.; Belušič, G.; Kelber, A. Evolution of insect color vision: From spectral sensitivity to visual ecology. Annu. Rev. Entomol. 2021, 66, 435–461. [Google Scholar] [CrossRef]
- Schlangen, L.; Lang, D.; Novotny, P.; Plischke, H.; Smolders, K.; Beersma, D.; Wulff, K.; Foster, R.; Cajochen, C.; Nikunen, H.; et al. Lighting for Health and Well-Being in Education, Work Places, Nursing Homes, Domestic Applications, and Smart Cities. 2014. Available online: https://lightingforpeople.eu/2016/wp-content/uploads/2016/03/SSLerate-3.2-3.4-v4.pdf (accessed on 1 December 2022).
- Briscoe, D.B.; Chittka, L. The evolution of color vision in insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef] [Green Version]
- Brehm, G. A new LED lamp for the collection of nocturnal Lepidoptera and a spectral comparison of light-trapping lamps. Nota Lepidopterol. 2017, 40, 87–108. [Google Scholar] [CrossRef] [Green Version]
- Brehm, G.; Niermann, J.; Nino LM, J.; Enseling, D.; Jüstel, T.; Axmacher, J.C.; Warrant, E.; Fiedler, K. Moths are strongly attracted to ultraviolet and blue radiation. Insect Conserv. Divers. 2021, 14, 188–198. [Google Scholar] [CrossRef]
- Sanmartín, P.; Vázquez-Nion, D.; Arines, J.; Cabo-Domínguez, L.; Prieto, B. Controlling growth and colour of phototrophs by using simple and inexpensive coloured lighting: A preliminary study in the Light4Heritage project towards future strategies for outdoor illumination. Int. Biodeterior. Biodegrad. 2017, 122, 107–115. [Google Scholar] [CrossRef]
- Sanmartín, P. New perspectives against biodeterioration through public lighting. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Joseph, E., Ed.; Springer: Cham, Switzerland, 2021; pp. 155–171. [Google Scholar]
- Longcore, T.; Aldern, H.L.; Eggers, J.F.; Flores, S.; Franco, L.; Hirshfield-Yamanishi, E.; Laina Petrinec, L.N.; Yan, W.A.; Barroso, A.M. Tuning the white light spectrum of light emitting diode lamps to reduce attraction of nocturnal arthropods. Philos. Trans. R. Soc. 2015, 370, 20140125. [Google Scholar]
- Hakami, A.R.; Khan, K.A.; Ghramh, H.A.; Ahmad, Z.; Al-Zayd, A. Impact of artificial light intensity on nocturnal insect diversity in urban and rural areas of the Asir province, Saudi Arabia. PLoS ONE 2020, 15, e0242315. [Google Scholar] [CrossRef] [PubMed]
- Stavenga, D.G.; Wehling, M.F.; Belušič, G. Functional interplay of visual, sensitizing and screening pigments in the eyes of Drosophila and other red-eyed dipteran flies. J. Physiol. 2017, 595, 5481–5494. [Google Scholar] [CrossRef] [Green Version]
- Kokkinn, M.J.; Williams, W.D. An experimental study of phototactic responses of Tanytarsus barbitarsis Freeman (Diptera: Chironomidae). Mar. Freshw. Res. 1989, 40, 693–702. [Google Scholar] [CrossRef]
- Mellor, H.E.; Hamilton, J. Navigation of Lutzomyia longipalpis (Diptera: Psychodidae) under dusk or starlight conditions. Bull. Entomol. Res. 2003, 93, 315–322. [Google Scholar] [CrossRef]
- Döring, T.F.; Kirchner, S.M.; Skorupski, P.; Hardie, J. Spectral sensitivity of the green photoreceptor of winged pea aphids. Physiol. Entomol. 2011, 36, 392–396. [Google Scholar] [CrossRef]
- McNeill, C.A.; Allan, S.A.; Koehler, P.G.; Pereira, R.M.; Weeks, E. Vision in the common bed bug Cimex lectularius L. (Hemiptera: Cimicidae): Eye morphology and spectral sensitivity. Med. Vet. Entomol. 2016, 30, 426–434. [Google Scholar]
- Diaz-Montano, J.; Campbell, J.F.; Phillips, T.W.; Throne, J.E. Evaluation of light attraction for the stored-product psocids, Liposcelis entomophila, Liposcelis paeta, and Liposcelis brunnea. J. Econ. Entomol. 2018, 111, 1476–1480. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Campbell, J.F.; Phillips, T.W.; Cohnstaedt, L.W.; Throne, J.E. Evaluation of light attraction for the stored-product psocid, Liposcelis bostrychophila. J. Pest. Sci. 2016, 89, 923–933. [Google Scholar] [CrossRef] [Green Version]
- van Langevelde, F.; Ettema, J.A.; Donners, M.; WallisDeVries, M.F.; Groenendijk, D. Effect of spectral composition of artificial light on the attraction of moths. Biol. Conserv. 2011, 144, 2274–2281. [Google Scholar] [CrossRef]
- Wakefield, A.; Broyles, M.; Stone, E.L.; Harris, S.; Jones, G. Quantifying the attractiveness of broad-spectrum street lights to aerial nocturnal insects. J. Appl. Ecol. 2018, 55, 714–722. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | N° of Replacements | Rain (L m−2) | Sunshine Duration (h) | Mean Relative Humidity (%) | Average Air Temperature (°C) |
|---|---|---|---|---|---|
| [1] 1–21 June | 3 | 3.18 ± 6.98 | 8.10 ± 4.93 | 78.95 ± 7.91 | 16.25 ± 3.32 |
| [2] 21–30 June | 3 | 0.58 ± 1.38 | 7.16 ± 4.38 | 80.56 ± 7.62 | 16.93 ± 2.23 |
| [3] 30–28 July | 4 | 0.44 ± 1.30 | 8.15 ± 3.41 | 81.25 ± 5.13 | 18.30 ± 1.54 |
| [4] 28–26 August | 3 | 2.76 ± 5.01 | 6.10 ± 3.33 | 83.03 ± 6.96 | 17.32 ± 2.08 |
| [5] 26 September–20 October | 2 | 4.27 ± 8.58 | 5.12 ± 3.69 | 87.88 ± 6.31 | 14.72 ± 1.26 |
| Order | % of Insects Trapped | N° Specimens | N° of Morphospecies |
|---|---|---|---|
| Diptera | 47.1 | 842 | 65 |
| Hemiptera | 25.0 | 457 | 28 |
| Psocoptera | 15.0 | 268 | 8 |
| Coleoptera | 5.5 | 100 | 14 |
| Lepidoptera | 4.0 | 74 | 24 |
| Hymenoptera | 2.6 | 47 | 14 |
| Neuroptera | 0.3 | 6 | 1 |
| Trichoptera | 0.3 | 6 | 3 |
| Thysanoptera | 0.2 | 4 | 3 |
| Total | 100 | 1804 | 160 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez, A.; Martín, L.; Arines, J.; Carballeira, R.; Sanmartín, P. Attraction of Insects to Ornamental Lighting Used on Cultural Heritage Buildings: A Case Study in an Urban Area. Insects 2022, 13, 1153. https://doi.org/10.3390/insects13121153
Méndez A, Martín L, Arines J, Carballeira R, Sanmartín P. Attraction of Insects to Ornamental Lighting Used on Cultural Heritage Buildings: A Case Study in an Urban Area. Insects. 2022; 13(12):1153. https://doi.org/10.3390/insects13121153
Chicago/Turabian StyleMéndez, Anxo, Luis Martín, Justo Arines, Rafael Carballeira, and Patricia Sanmartín. 2022. "Attraction of Insects to Ornamental Lighting Used on Cultural Heritage Buildings: A Case Study in an Urban Area" Insects 13, no. 12: 1153. https://doi.org/10.3390/insects13121153
APA StyleMéndez, A., Martín, L., Arines, J., Carballeira, R., & Sanmartín, P. (2022). Attraction of Insects to Ornamental Lighting Used on Cultural Heritage Buildings: A Case Study in an Urban Area. Insects, 13(12), 1153. https://doi.org/10.3390/insects13121153