
Biodiversity of Forensically Relevant Blowflies (Diptera: Calliphoridae) at the Anthropology Research Facility in Knoxville, Tennessee, USA

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
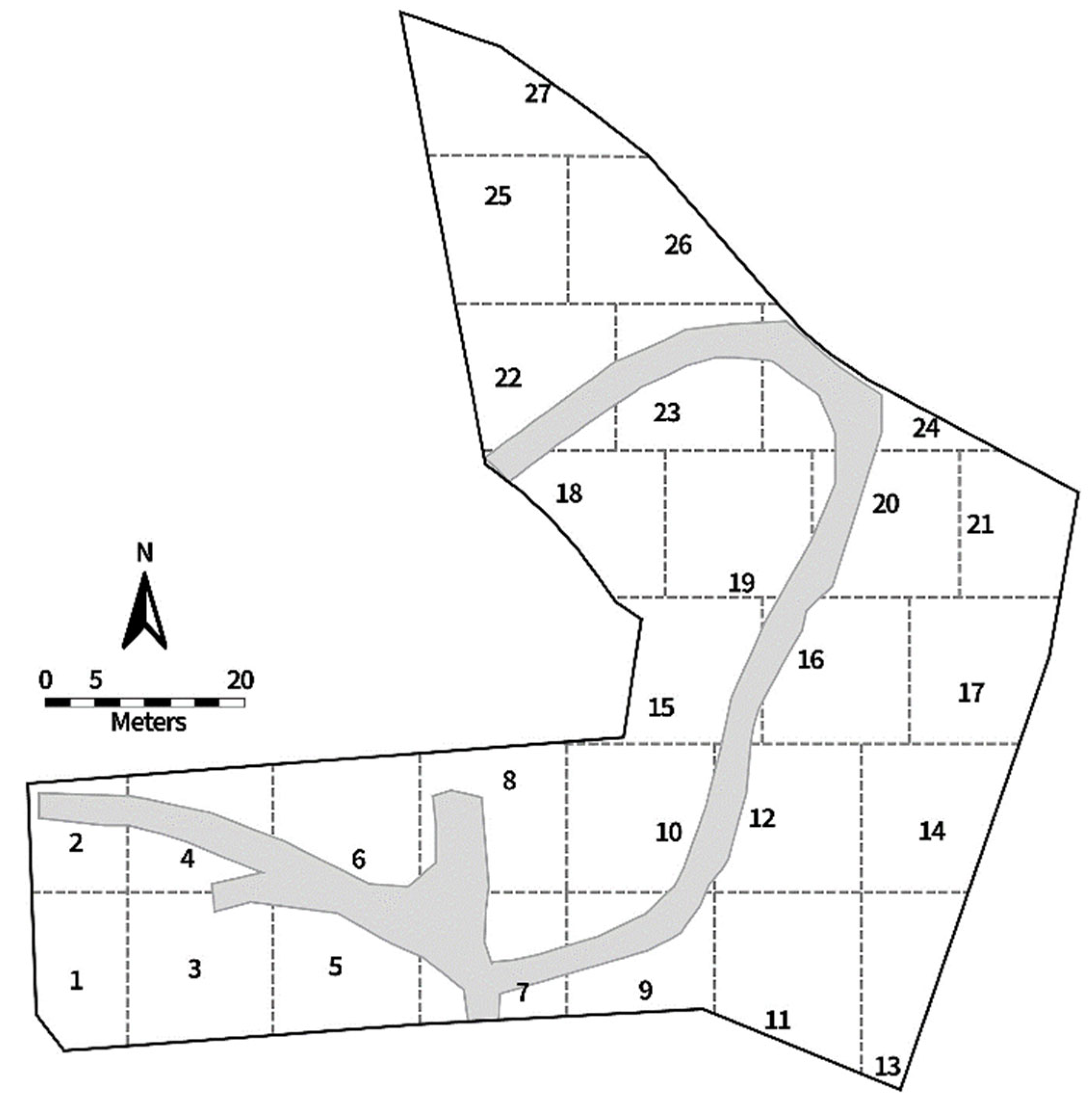
2.1. Experimental Location and Duration
2.2. Trap Design and Placement
2.3. Processing
2.4. Temperature Data
2.5. Data Analyses
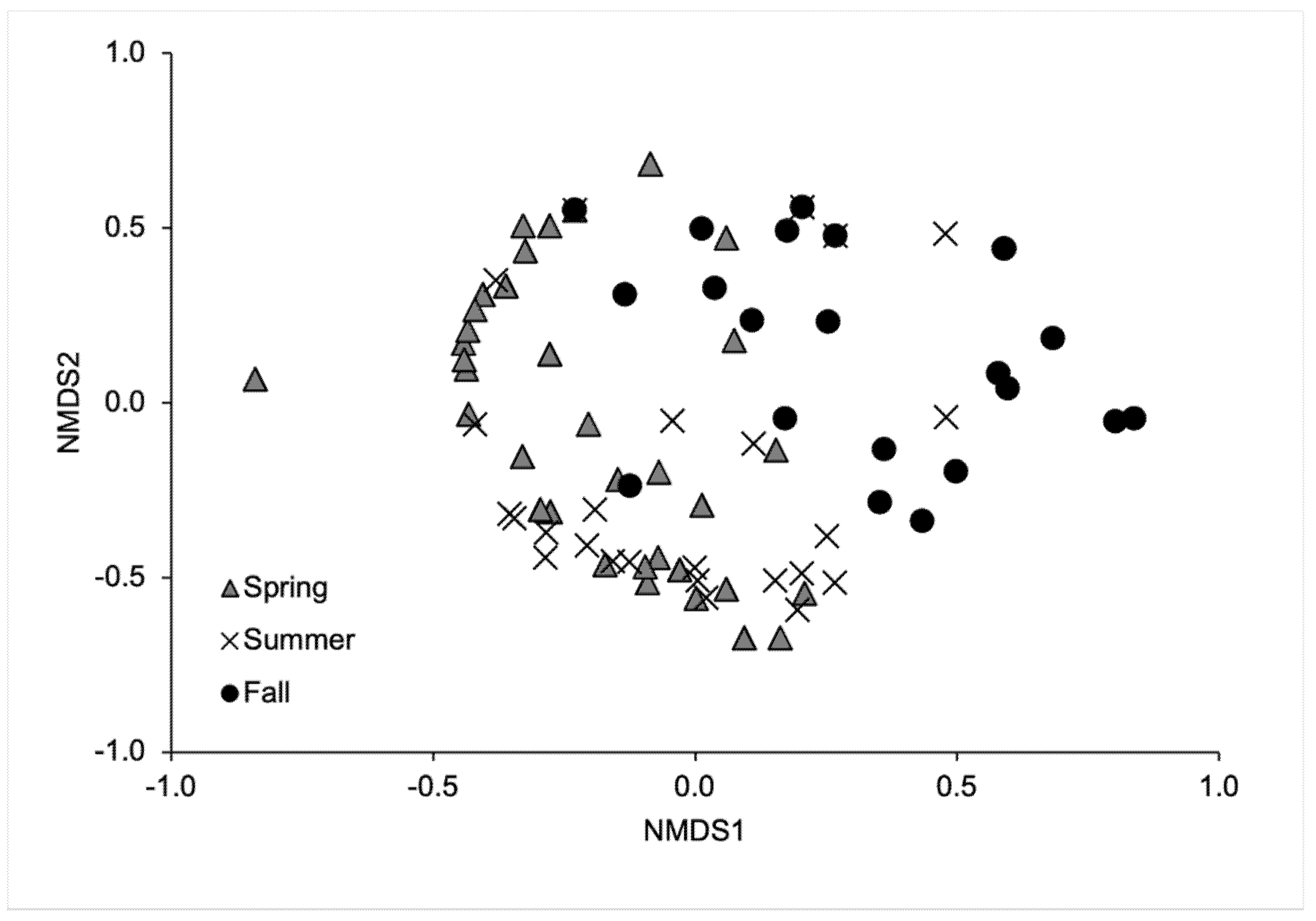
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gennard, D. Forensic Entomology: An Introduction, 2nd ed.; John Wiley & Sons: West Sussex, UK, 2012; pp. 1–11. [Google Scholar]
- Van Laerhoven, S.L. Ecological theory of community assembly and its application in forensic entomology. In Forensic Entomology: The Utility of Arthropods in Legal Investigations, 3rd ed.; Byrd, J.H., Tomberlin, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 387–404. [Google Scholar]
- Tomberlin, J.K.; Mohr, R.; Benbow, M.E.; Tarone, A.M.; Van Laerhoven, S. A roadmap for bridging basic and applied research in forensic entomology. Annu. Rev. Entomol. 2011, 56, 401–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.S.; VanLaerhoven, S.L. Initial studies on insect succession on carrion in southwestern British Columbia. J. Forensic Sci. 1996, 41, 617–625. [Google Scholar] [CrossRef]
- Weidner, L.M.; Monzon, M.A.; Hamilton, G.C. Death eaters respond to the dark mark of decomposition day and night: Observations of initial insect activity on piglet carcasses. Int. J. Legal Med. 2016, 130, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Brundage, A.; Bros, S.; Honda, J.Y. Seasonal and habitat abundance and distribution of some forensically important blow flies (Diptera: Calliphoridae) in Central California. Forensic Sci. Int. 2011, 212, 115–120. [Google Scholar] [CrossRef]
- Farinha, A.; Dourado, C.G.; Centeio, N.; Oliveira, A.R.; Dias, D.; Rebelo, M.T. Small bait traps as accurate predictors of dipteran early colonizers in forensic studies. J. Insect Sci. 2014, 14, 77. [Google Scholar] [CrossRef] [Green Version]
- Weidner, L.M.; Jennings, D.E.; Tomberlin, J.K.; Hamilton, G.C. Seasonal and geographic variation in biodiversity of forensically important blow flies (Diptera: Calliphoridae) in New Jersey, USA. J. Med. Entomol. 2015, 52, 937–946. [Google Scholar] [CrossRef]
- Weidner, L.M.; Gemmellaro, M.D.; Tomberlin, J.K.; Hamilton, G.C. Evaluation of bait traps as a means to predict initial blow fly (Diptera: Calliphoridae) communities associated with decomposing swine remains in New Jersey, USA. Forensic Sci. Int. 2017, 278, 95–100. [Google Scholar] [CrossRef]
- LeBlanc, K.; Boudreau, D.R.; Moreau, G. Small bait traps may not accurately reflect the composition of necrophagous Diptera associated to remains. Insects 2021, 12, 261. [Google Scholar] [CrossRef]
- Early, M.; Goff, M.L. Arthropod succession patterns in exposed carrion on the island of O’ahu, Hawaiian Islands, USA. J. Med. Entomol. 1986, 23, 520–531. [Google Scholar] [CrossRef]
- Tabor, K.L.; Fell, R.D.; Brewster, C.C. Insect fauna visiting carrion in Southwest Virginia. Forensic Sci. Int. 2005, 150, 73–80. [Google Scholar] [CrossRef]
- Archer, M.S.; Elgar, M.A. Effects of decomposition on carcass attendance in a guild of carrion-breeding flies. Med. Vet. Entomol. 2003, 17, 263–271. [Google Scholar] [CrossRef]
- Feddern, N.; Mitchell, E.A.D.; Amendt, J.; Szelecz, I.; Seppey, C.V.W. Decomposition and insect colonization patterns of pig cadavers lying on forest soil and suspended above ground. Forensic Sci. Med. Pathol. 2019, 15, 342–351. [Google Scholar] [CrossRef]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Tippanun, J. Survey of forensically-relevant fly species in Chiang Mai, Northern Thailand. J. Vector Ecol. 2003, 28, 135–138. [Google Scholar]
- Anderson, G.S. The use of insects in death investigations: An analysis of cases in British Columbia over a five-year period. Can. Soc. Forensic Sci. 1995, 28, 277–292. [Google Scholar] [CrossRef]
- Goff, M.L.; Early, M.; Odom, C.B.; Tullis, K. A preliminary checklist of arthropods associated with exposed carrion in the Hawaiian Islands. Proc. Hawaii Entomol. Soc. 1986, 26, 53–57. [Google Scholar]
- Sanford, M.R. Insects and associated arthropods analyzed during medicolegal death investigations in Harris County, Texas, USA: January 2013–April 2016. PLoS ONE 2017, 12, e0179404. [Google Scholar] [CrossRef]
- Sanford, M.R. Comparing species composition of passive trapping of adult flies with larval collections from the body during scene-based medicolegal death investigations. Insects 2017, 8, 36. [Google Scholar] [CrossRef]
- Weidner, L.M.; Nigoghosian, G.; Yoho, K.C.; Parrott, J.J.; Hans, K.R. An analysis of forensically important insects associated with human remains in Indiana, USA. J. Forensic Sci. 2020, 65, 2036–2041. [Google Scholar] [CrossRef]
- Vanin, S.; Gherardi, M.; Bugelli, V.; Di Paolo, M. Insects found on a human cadaver in central Italy including the blowfly Calliphora loewi (Diptera: Calliphoridae), a new species of forensic interest. Forensic Sci. Int. 2011, 207, e30–e33. [Google Scholar] [CrossRef]
- Oliveira, T.C.; Vasconcelos, S.D. Insects (Diptera) associated with cadavers at the Institute of Legal Medicine in Pernambuco, Brazil: Implications for forensic entomology. Forensic Sci. Int. 2010, 198, 97–102. [Google Scholar] [CrossRef]
- Shin, S.E.; Lee, H.J.; Park, J.H.; Ko, K.S.; Kim, Y.H.; Kim, K.R.; Park, S.H. The first survey of forensically important entomofauna collected from medicolegal autopsies in South Korea. BioMed. Res. Int. 2015, 606728. [Google Scholar] [CrossRef]
- Talebzadeh, F.; Ghadipasha, M.; Gharedaghi, J.; Yeksan, N.; Akbarzadeh, K.; Oshaghi, M.A. Insect Fauna of Human Cadavers in Tehran District. J. Arthropod-Borne Dis. 2017, 11, 363–370. Available online: https://jad.tums.ac.ir/index.php/jad/article/view/717 (accessed on 10 October 2020).
- Moemenbellah-Fard, M.D.; Keshavarzi, D.; Fereidooni, M.; Soltani, A. First survey of forensically important insects from human corpses in Shiraz, Iran. J. Forensic Leg. Med. 2018, 54, 62–68. [Google Scholar] [CrossRef]
- Al-Qahtni, A.H.; Mashaly, A.M.; Alajmi, R.A.; Alshehri, A.A.; Al-Musawi, Z.M.; Al-Khalifa, M.S. Forensic insects attracted to human cadavers in a vehicular environment in Riyadh, Saudi Arabia. Saudi J. Biol. Sci. 2019, 26, 1499–1502. [Google Scholar] [CrossRef]
- Al-Khalifa, M.S.; Mashaly, A.M.; Al-Qahtni, A.H. Insect species colonized indoor and outdoor human corpses in Riyadh, Saudi Arabia. J. King Saud Univ. Sci. 2020, 32, 1812–1817. [Google Scholar] [CrossRef]
- Lutz, L.; Zehner, R.; Verhoff, M.A.; Bratzke, H.; Amendt, J. It is all about the insects: A retrospective on 20 years of forensic entomology highlights the importance of insects in legal investigations. Int. J. Legal. Med. 2021, 135, 2637–2651. [Google Scholar] [CrossRef]
- Matuszewski, S.; Hall, M.J.R.; Moreau, G.; Schoenly, K.G.; Tarone, A.M.; Villet, M.H. Pigs vs people: The use of pigs as analogues for humans in forensic entomology and taphonomy research. Int. J. Legal Med. 2020, 134, 793–810. [Google Scholar] [CrossRef] [Green Version]
- Forbes, S. Body farms. Forensic Sci. Med. Pat. 2017, 13, 477–479. [Google Scholar] [CrossRef]
- Wallman, J.F. Body farms. Forensic Sci. Med. Pat. 2017, 13, 487–489. [Google Scholar] [CrossRef]
- Wescott, D.J. Recent advances in forensic anthropology: Decomposition research. Forensic Sci. Res. 2018, 3, 278–293. [Google Scholar] [CrossRef] [Green Version]
- Pecsi, E.L.; Bronchti, G.; Crispino, F.; Forbes, S.L. Perspectives on the establishment of a Canadian human taphonomic facility: The experience of REST [ES]. Forensic Sci. Int. Synergy 2020, 2, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Varlet, V.; Joye, C.; Forbes, S.L.; Grabherr, S. Revolution in death sciences: Body farms and taphonomics blooming. A review investigating the advantages, ethical and legal aspects in a Swiss context. Int. J. Legal Med. 2020, 134, 1875–1895. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, W.C.; Bass, W.M. Insect activity and its relationship to decay rates of human cadavers in East Tennessee. J. Forensic Sci. 1983, 28, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Shahid, S.A.; Hall, R.D.; Haskell, N.H.; Merritt, R.W. Chrysomya rufifacies (Macquart) (Diptera: Calliphoridae) established in the vicinity of Knoxville, Tennessee, USA. J. Forensic Sci. 2000, 45, 896–897. [Google Scholar] [CrossRef]
- Shahid, S.A.; Schoenly, K.; Haskell, N.H.; Hall, R.D.; Zhang, W. Carcass enrichment does not alter decay rates or arthropod community structure: A test of the arthropod saturation hypothesis at the Anthropology Research Facility in Knoxville, Tennessee. J. Med. Entomol. 2003, 40, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y. First oviposition timing of blow flies on human cadavers. In Proceedings of the 65th American Academy of Forensic Sciences, Washington, DC, USA, 18–23 February 2013; pp. 314–315. [Google Scholar]
- Jeong, Y. Decomposition pattern of human heads as related to insect activity. In Proceedings of the 65th American Academy of Forensic Sciences, Washington, DC, USA, 18–23 February 2013; pp. 378–379. [Google Scholar]
- Jeong, Y.; Hauther, K. When do blowflies lay eggs on human bodies? In Proceedings of the 2018 North American Forensic Entomology Association, Orlando, FL, USA, 23–25 July 2018. [Google Scholar]
- Schoenly, K.G.; Haskell, N.H.; Hall, R.D.; Gbur, J.R. Comparative performance and complementarity of four sampling methods and arthropod preference tests from human and porcine remains at the Forensic Anthropology Center in Knoxville, Tennessee. J. Med. Entomol. 2007, 44, 881–894. [Google Scholar] [CrossRef]
- Peel, M.; Finlayson, B.; McMahon, T. Updated world map of the Köppen–Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.; Jantz, L.M.; Smith, J. Investigation into seasonal scavenging patterns of raccoons on human decomposition. J. Forensic. Sci. 2016, 61, 467–471. [Google Scholar] [CrossRef]
- Jones, N.; Whitworth, T.; Marshall, S.A. Blow flies of North America: Keys to the subfamilies and genera of Calliphoridae, and to the species of the subfamilies Calliphorinae, Luciliinae and Chrysomyinae. Can. J. Arth. Identif. 2019, 39, 1–191. [Google Scholar] [CrossRef]
- Jewiss-Gaines, A.; Marshall, S.A.; Whitworth, T.L. Cluster flies (Calliphoridae: Polleninae: Pollenia) of North America. Can. J. Arth. Identif. 2012, 19, 1–19. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org (accessed on 10 October 2020).
- Reed, H.B. A study of dog carcass communities in Tennessee, with special reference to the insects. Am. Midl. Nat. 1958, 59, 213–245. [Google Scholar] [CrossRef]
- Owings, C.G.; Mckee-Zech, H.; Steadman, D.W. First record of the oriental latrine fly, Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae), in Tennessee, USA. Acta Parasitol. 2021, 66, 1079–1081. [Google Scholar] [CrossRef]
- Schoenly, K. Demographic bait trap. Environ. Entomol. 1981, 10, 615–617. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Institution | City/Country | Establishment Year |
|---|---|---|
| ARF/University of Tennessee | Knoxville, TN, USA | 1980 |
| FOREST/Western Carolina University | Cullowhee, NC, USA | 2007 |
| FARF/Texas State University | San Marcos, TX, USA | 2008 |
| STAFS/Sam Houston State University | Huntsville, TX, USA | 2008 |
| CFAR/Southern Illinois University | Carbondale, IL, USA | 2012 |
| FIRS/Colorado Mesa University | Grand Junction, CO, USA | 2013 |
| AFTER/University of Technology Sydney | Yarramundi, New South Wales, Australia | 2016 |
| ARISTA/Amsterdam’s Academic Medical Center | Amsterdam, Netherlands | 2017 |
| FIRST/University of South Florida | Tampa, FL, USA | 2018 |
| FROST/Northern Michigan University | Marquette, MI, USA | 2018 |
| REST[ES]/University of Québec–Trois Rivières | Québec, Canada | 2019 |
| Dates of Trap Deployment and Collection | Temperature (°C) | Humidity (%) | Precipitation (mm) | |||
|---|---|---|---|---|---|---|
| Average | Range | Average | Range | Trap | Monthly | |
| 3/4–3/5/18 * | 13.9 | 10.9–14.3 | 33.3 | 32.0–40.0 | 0 | 144.5 |
| 3/19–3/20/18 | 15.7 | 12.3–21.6 | 83.2 | 51.0–99.0 | 8.4 | |
| 4/4–4/5/18 | 8.3 | 1.3–15.3 | 49.3 | 25.0–79.0 | 10.9 | 108.7 |
| 4/19–4/20/18 | 9.0 | 2.6–17.1 | 60.4 | 32.0–92.0 | 0 | |
| 5/4–5/5/18 | 22.8 | 18.1–29.0 | 70.8 | 46.0–95.0 | 0 | 87.6 |
| 5/19–5/20/18 | 24.7 | 19.1–30.9 | 78.1 | 48.0–99.0 | 23.1 | |
| 6/4–6/5/18 | 21.2 | 15.9–28.2 | 64.0 | 36.0–89.0 | 0.5 | 107.2 |
| 6/19–6/20/18 | 27.5 | 22.3–34.9 | 70.1 | 45.0–90.0 | 0 | |
| 7/4–7/5/18 | 27.9 | 23.4–33.1 | 74.1 | 58.0–89.0 | 0 | 127.5 |
| 7/19–7/20/18 | 25.7 | 21.2–32.3 | 77.2 | 53.0–92.0 | 0 | |
| 8/3–8/4/18 | 23.8 | 20.8–29.6 | 89.7 | 66.0–96.0 | 39.6 | 118.9 |
| 8/19–8/20/18 | 25.6 | 23.5–30.1 | 88.1 | 68.0–95.0 | 0.5 | |
| 9/2–9/3/18 | 25.6 | 21.3–37.1 | 82.0 | 43.0–97.0 | 25.4 | 191.0 |
| 9/20–9/21/18 | 27.1 | 22.2–39.5 | 73.1 | 37.0–93.0 | 0 | |
| 10/4–10/5/18 | 24.5 | 20.4–33.3 | 83.5 | 53.0–98.0 | 0 | 76.2 |
| 10/18–10/19/18 | 12.6 | 6.7–22.7 | 75.1 | 34.0–99.0 | 0 | |
| 11/3–11/4/18 | 9.3 | 3.8–18.1 | 78.8 | 44.0–99.0 | 0 | 142.2 |
| 11/17–11/18/18 | 8.2 | 1.4–18.1 | 88.8 | 43.0–99.0 | 0 | |
| 12/6–12/7/18 † | 5.6 | 3.6–11.2 | 59.4 | 36.0–77.0 | 0 | 189.5 |
| 12/19–12/20/18 † | 7.3 | 4.6–14.3 | 81.1 | 46.0–99.0 | 18.0 | |
| 1/3–1/4/19 † | 9.5 | 8.4–12.0 | 98.3 | 90.0–99.0 | 21.6 | 136.4 |
| 1/19–1/20/19 † | 5.5 | −2.7–12.4 | 94.9 | 79.0–99.0 | 40.1 | |
| 2/4/–2/5/19 † | 11.7 | 8.1–18.0 | 88.6 | 70.0–99.0 | 1.5 | 325.9 |
| 2/20–2/21/19 † | 9.2 | 8.6–10.6 | 98.9 | 94.0–99.0 | 47.4 | |
| 3/4–3/5/19 † | −1.0 | −5.6–7.6 | 64.1 | 25.0–86.0 | 38.6 | 109.2 |
| 3/19–3/20/19 † | 9.1 | −1.0–25.9 | 51.2 | 13.0–93.0 | 0 | |
| 4/4–4/5/19 | 18.1 | 10.7–29.9 | 59.6 | 19.0–99.0 | 14.9 | 111.5 |
| 4/20–4/21/19 | 10.4 | 7.2–25.4 | 87.0 | 31.0–99.0 | 2.5 | |
| Genus | Species * | Overall Totals (%) | Spring (%) | Summer (%) | Fall (%) |
|---|---|---|---|---|---|
| Calliphora | 110 (3.5%) | 66 (4.6%) | 20 (1.5%) | 24 (6.4%) | |
| C. livida | 15 (0.5%) | 13 (0.9%) | 1 (0.1%) | 1 (0.3%) | |
| C. vicina | 63 (2.0%) | 38 (2.7%) | 18 (1.3%) | 7 (1.9%) | |
| C. vomitoria | 32(1.0%) | 15 (1.1%) | 1 (0.1%) | 16 (4.3%) | |
| Chrysomya | 25 (0.8%) | - | 1 (0.1%) | 24 (6.4%) | |
| Ch. megacephala | 23 (0.7%) | - | 1 (0.1%) | 22 (5.9%) | |
| Ch. rufifacies | 2 (0.1%) | - | - | 2 (0.5%) | |
| Cochliomyia | 21 (0.7%) | 3 (0.2%) | 18 (1.3%) | - | |
| Co. macellaria | 21 (0.7%) | 3 (0.2%) | 18 (1.3%) | - | |
| Cynomya | 2 (0.1%) | 1 (0.1%) | - | 1 (0.3%) | |
| Cy. cadaverina | 2 (0.1%) | 1 (0.1%) | - | 1 (0.3%) | |
| Lucilia | 844 (26.7%) | 211 (14.8%) | 401 (29.5%) | 232 (61.8%) | |
| L. coeruleiviridis | 649 (20.6%) | 138 (9.7%) | 290 (21.3%) | 221 (58.9%) | |
| L. cuprina | 3 (0.1%) | - | - | 3 (0.8%) | |
| L. illustris | 170 (5.4%) | 72 (5.1%) | 96 (7.1%) | 2 (0.5%) | |
| L. sericata | 22 (0.7%) | 1 (0.1%) | 15 (1.1%) | 6 (1.6%) | |
| Phormia | 2080 (65.9%) | 1119 (78.6%) | 870 (64.0%) | 91 (24.2%) | |
| P. regina | 2080 (65.9%) | 1119 (78.6%) | 870 (64.0%) | 91 (24.2%) | |
| Protophormia | 74 (2.3%) | 22 (1.5%) | 49 (3.6%) | 3 (0.8%) | |
| Pr. terraenovae | 74 (2.3%) | 22 (1.5%) | 49 (3.6%) | 3 (0.8%) |
| Season | Richness | Simpson’s Index of Diversity |
|---|---|---|
| Spring | 10 | 0.368 |
| Summer | 10 | 0.538 |
| Fall | 12 | 0.589 |
| Winter | 0 | - |
| Species * | Reed [50] | Shahid et al. [36] | Schoenly et al. [41] | Present Study |
|---|---|---|---|---|
| C. livida | + | – | – | + |
| C. vicina | + | + | – | + |
| C. vomitoria | + | – | – | + |
| C. terraenovae | + | – | – | – |
| Ch. megacephala | – | – | – | + |
| Ch. rufifacies | – | + | – | + |
| Co. macellaria | + | + | + | + |
| Cy. cadaverina | + | – | – | + |
| L. coeruleiviridis | + | + | + | + |
| L. cuprina | + | – | – | + |
| L. illustris | + | + | + | + |
| L. sericata | + | + | + | + |
| L. cluvia | – | + | – | – |
| P. regina | + | + | + | + |
| Pr. terraenovae | – | – | – | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.; Weidner, L.M.; Pergande, S.; Gemmellaro, D.; Jennings, D.E.; Hans, K.R. Biodiversity of Forensically Relevant Blowflies (Diptera: Calliphoridae) at the Anthropology Research Facility in Knoxville, Tennessee, USA. Insects 2022, 13, 109. https://doi.org/10.3390/insects13020109
Jeong Y, Weidner LM, Pergande S, Gemmellaro D, Jennings DE, Hans KR. Biodiversity of Forensically Relevant Blowflies (Diptera: Calliphoridae) at the Anthropology Research Facility in Knoxville, Tennessee, USA. Insects. 2022; 13(2):109. https://doi.org/10.3390/insects13020109
Chicago/Turabian StyleJeong, Yangseung, Lauren M. Weidner, Simon Pergande, Denise Gemmellaro, David E. Jennings, and Krystal R. Hans. 2022. "Biodiversity of Forensically Relevant Blowflies (Diptera: Calliphoridae) at the Anthropology Research Facility in Knoxville, Tennessee, USA" Insects 13, no. 2: 109. https://doi.org/10.3390/insects13020109
APA StyleJeong, Y., Weidner, L. M., Pergande, S., Gemmellaro, D., Jennings, D. E., & Hans, K. R. (2022). Biodiversity of Forensically Relevant Blowflies (Diptera: Calliphoridae) at the Anthropology Research Facility in Knoxville, Tennessee, USA. Insects, 13(2), 109. https://doi.org/10.3390/insects13020109