
Geographical Variation of Honey Bee (Apis mellifera L. 1758) Populations in South-Eastern Morocco: A Geometric Morphometric Analysis

 ,
,  ,
,  ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
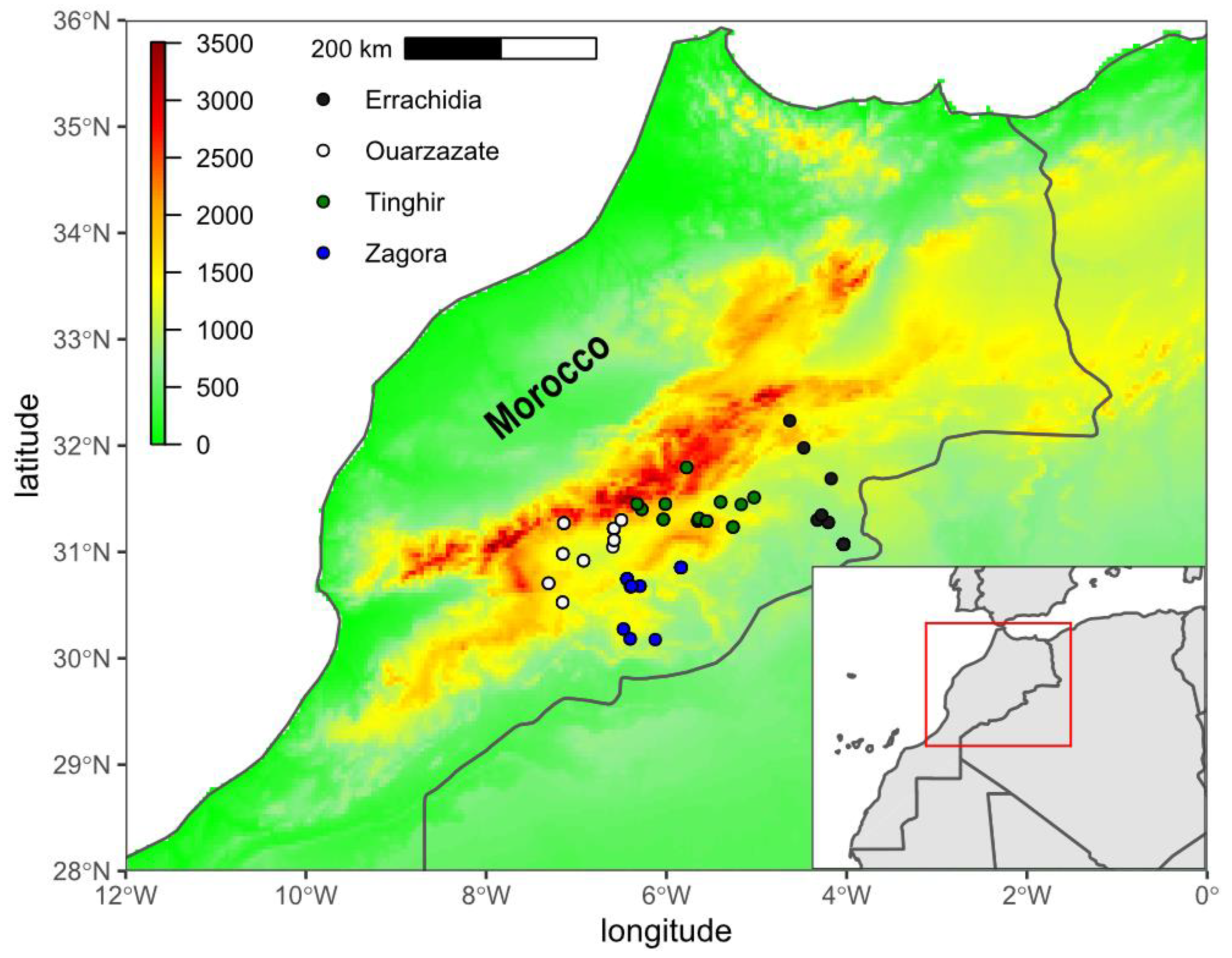
2.1. Sampling
2.2. Geometric Morphometric Analysis
2.3. Statistical Analysis
3. Results
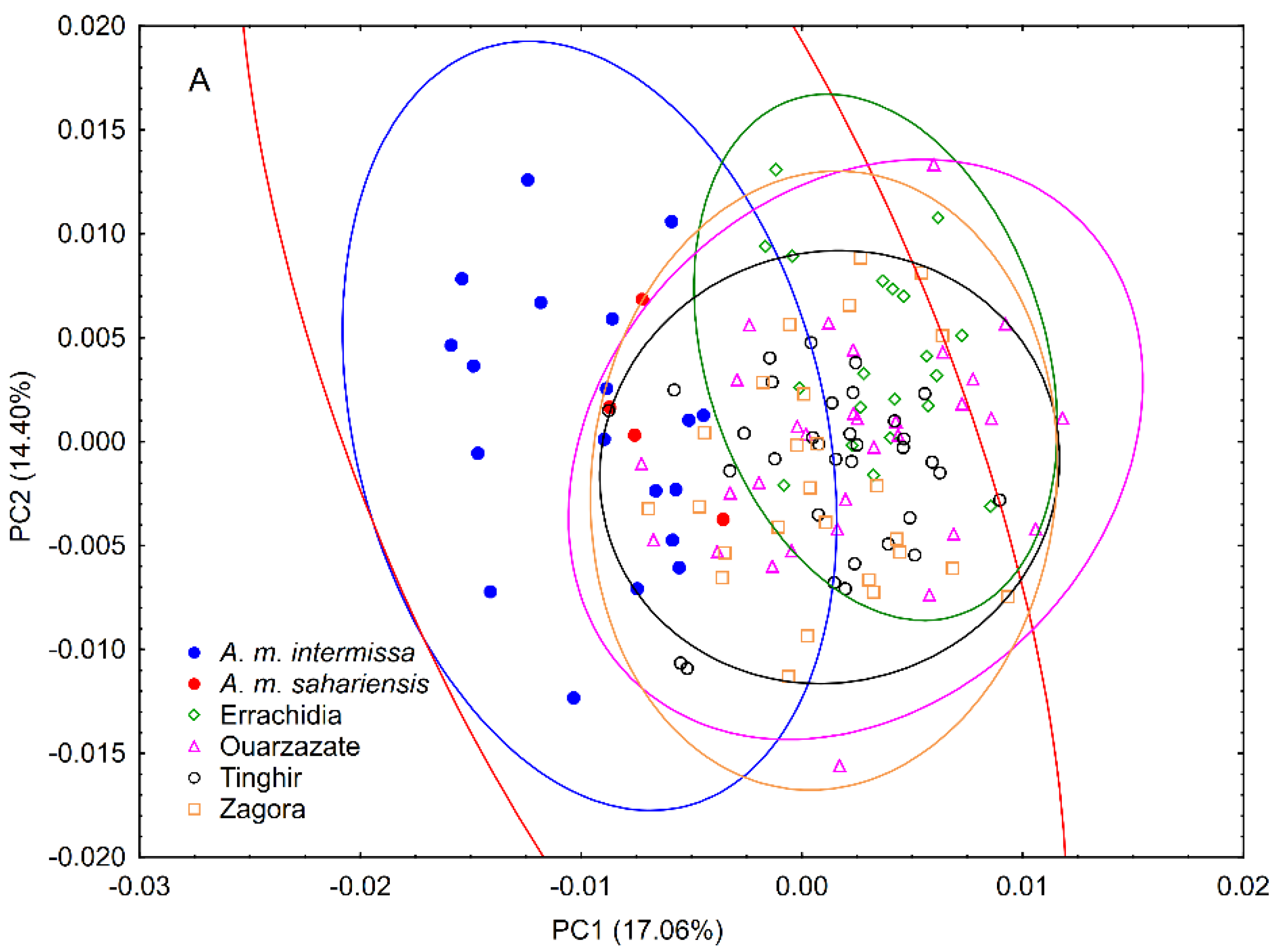
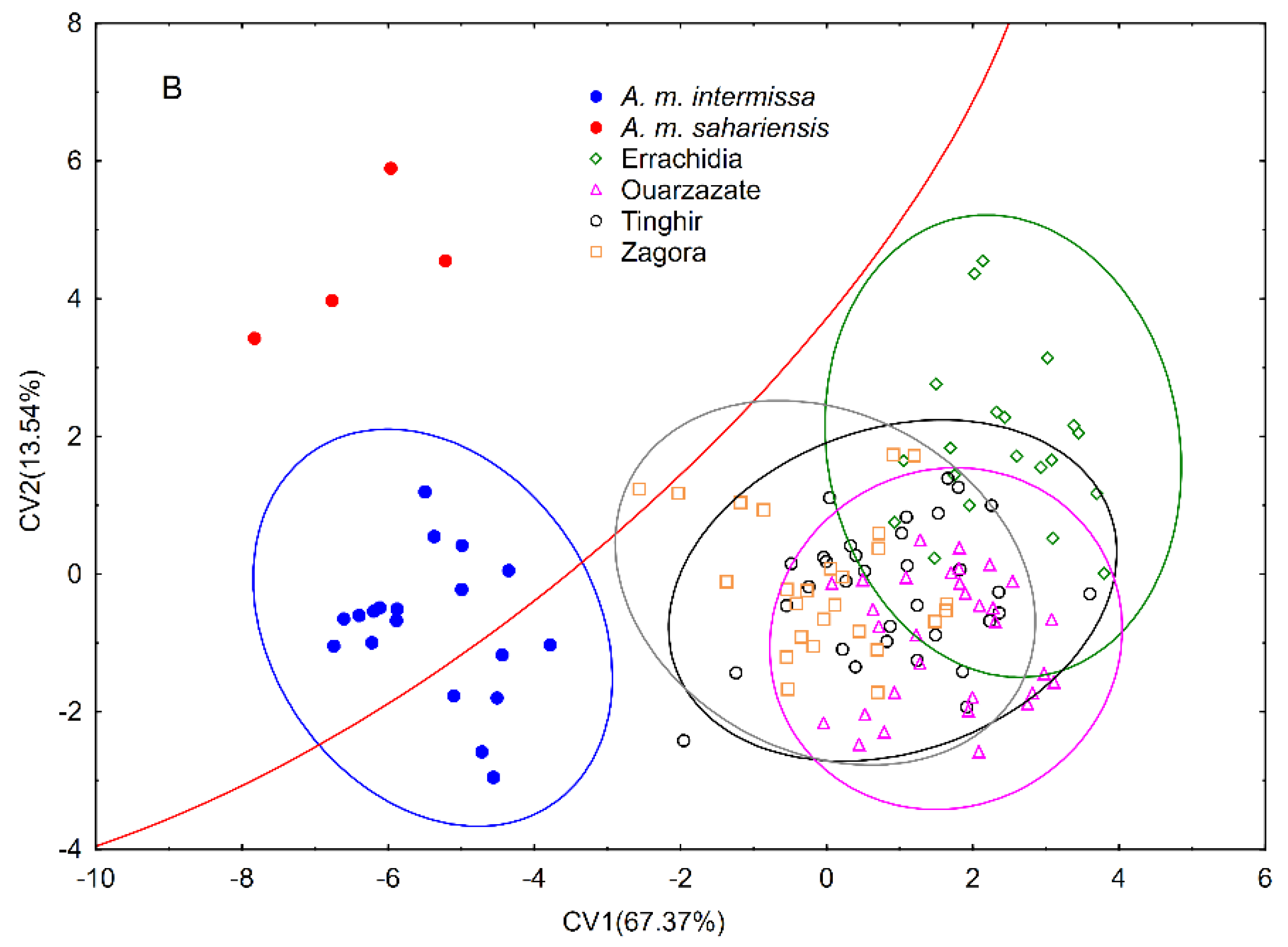
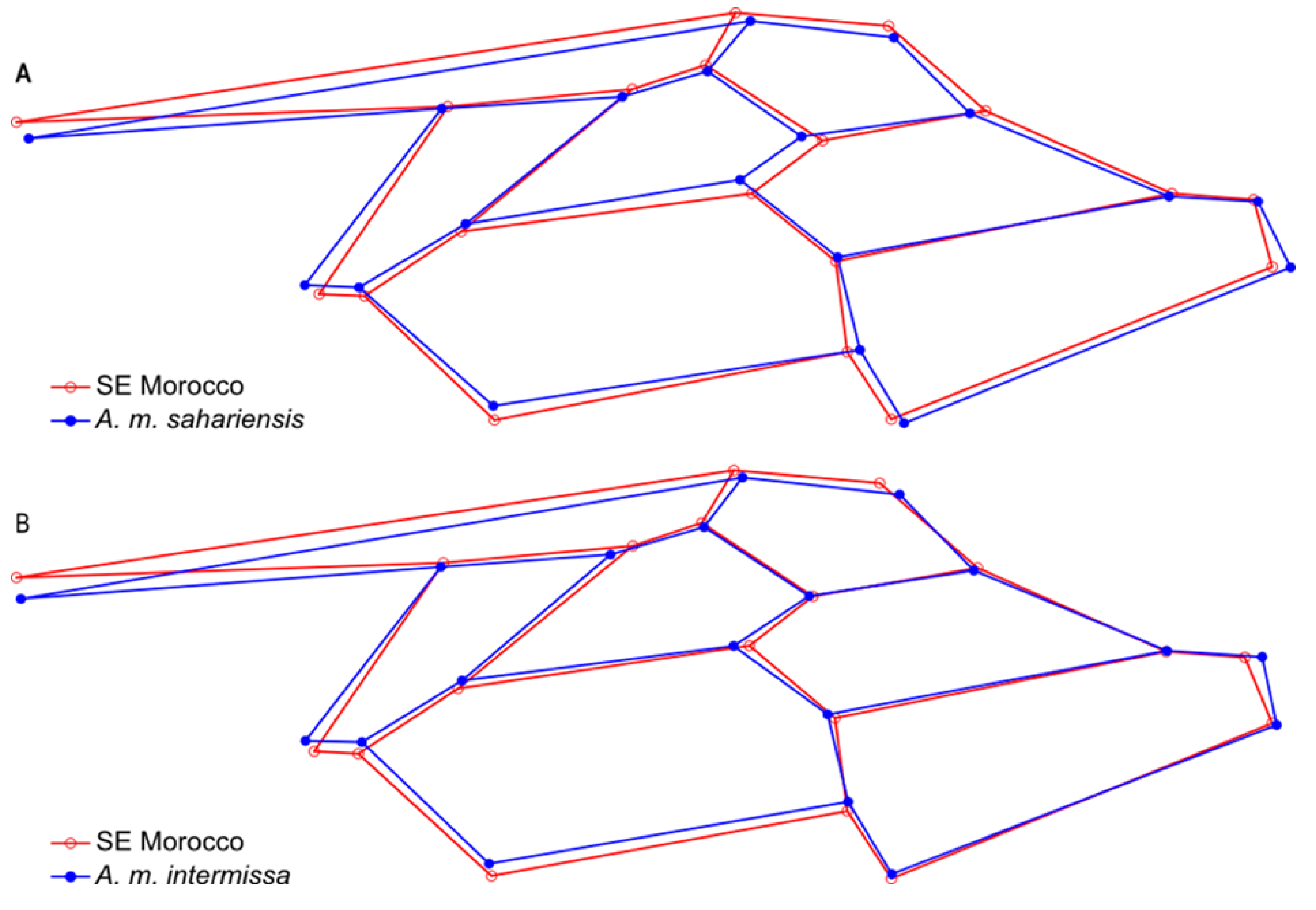
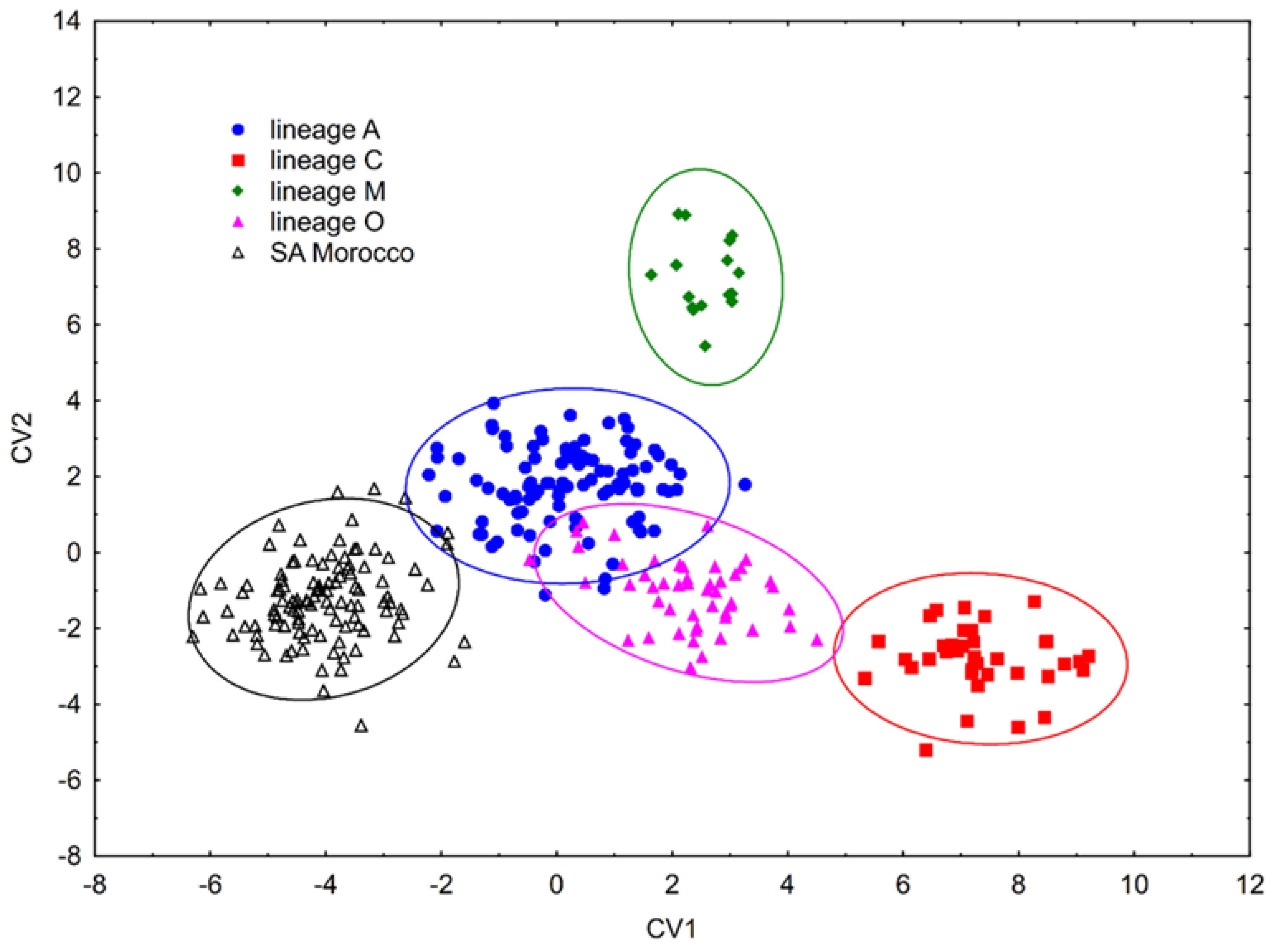
3.1. Wing Shape
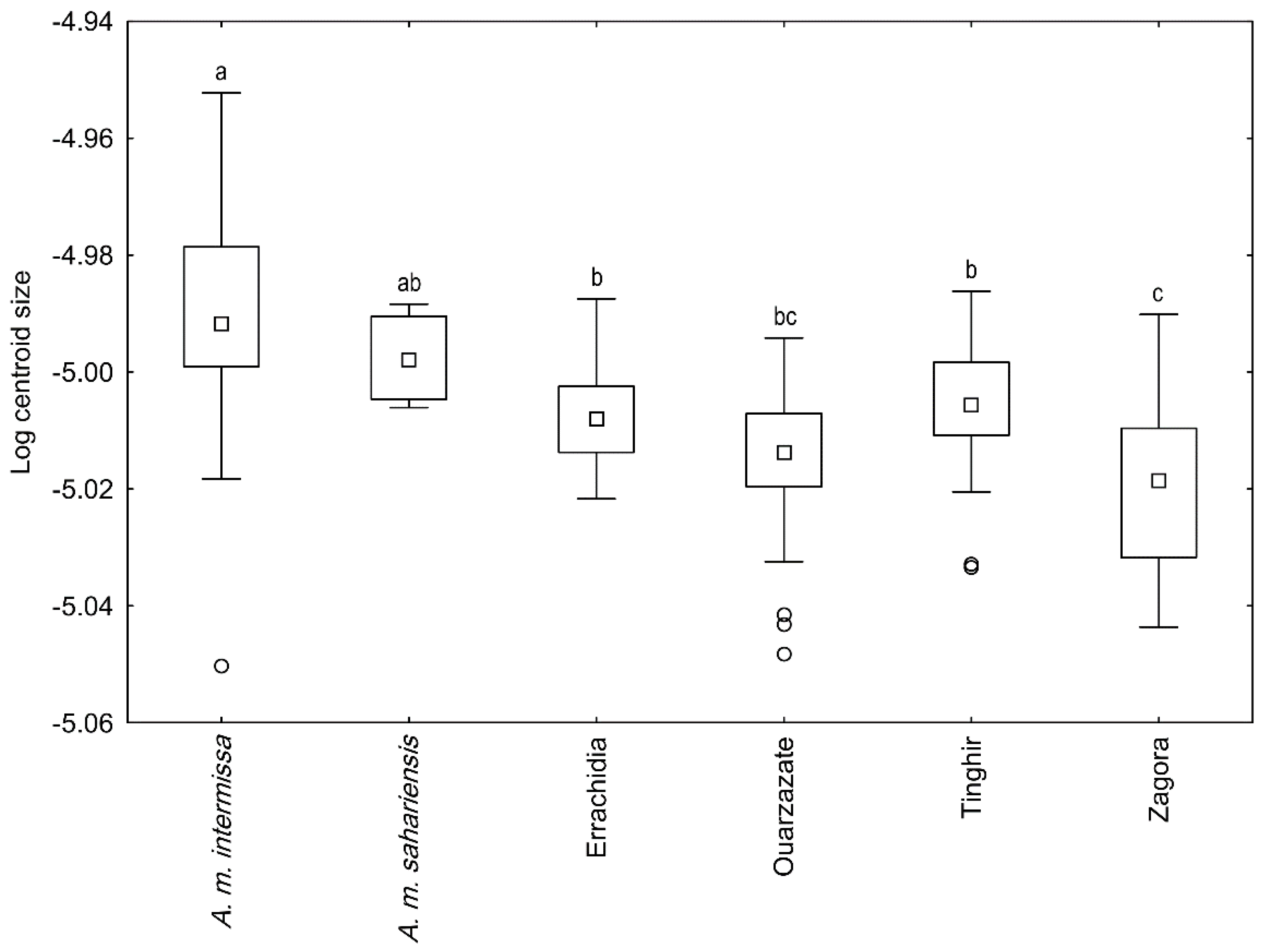
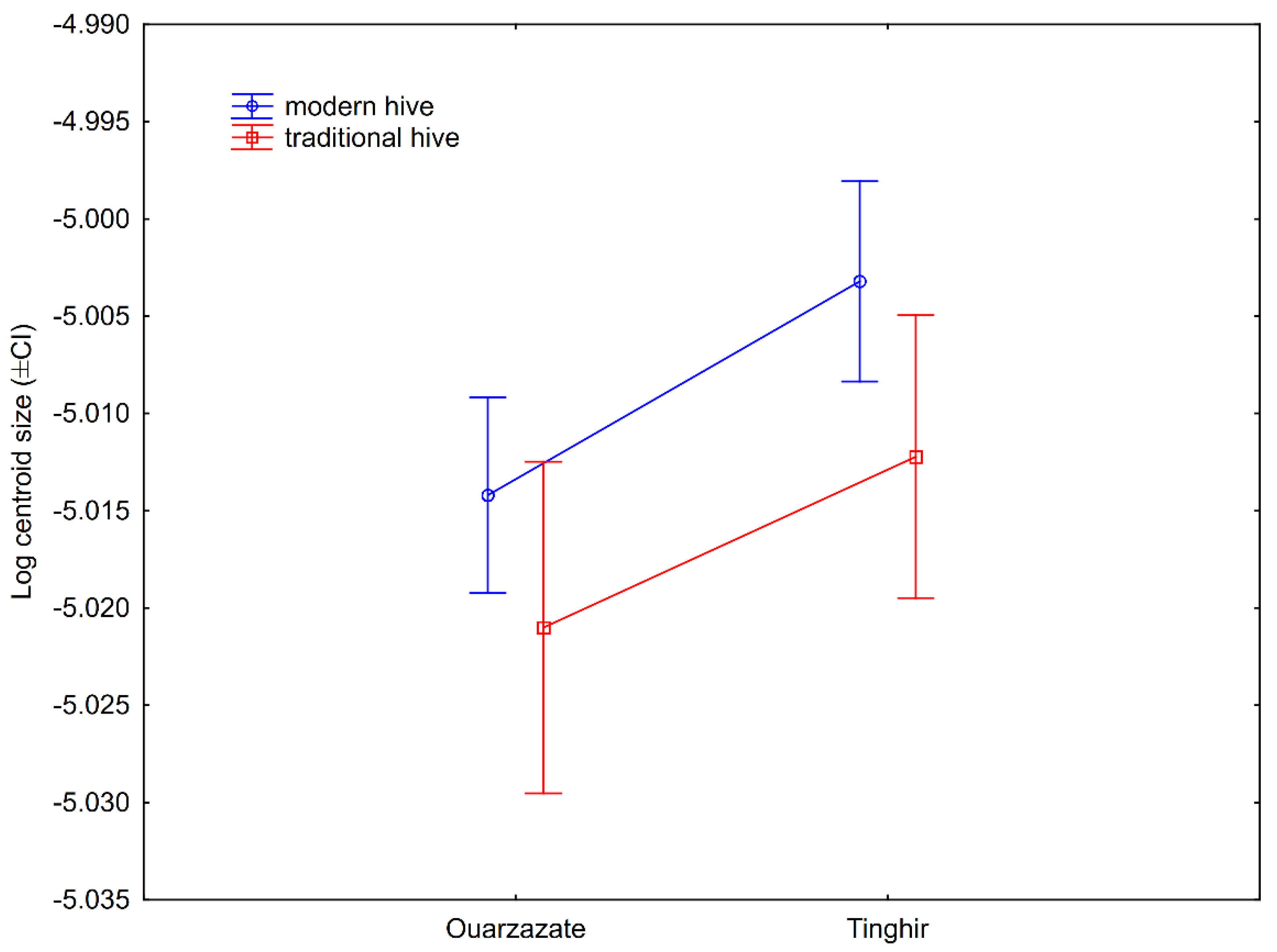
3.2. Wing Size
3.3. Standard Morphometry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruttner, F. Biogeography and Taxonomy of Honeybees; Springer: Berlin, Germany, 1988. [Google Scholar]
- Requier, F.; Garnery, L.; Kohl, P.L.; Njovu, H.K.; Pirk, C.W.W.; Crewe, R.M.; Steffan-Dewenter, I. The Conservation of Native Honey Bees Is Crucial. Trends Ecol. Evol. 2019, 34, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.S. The Taxonomy of Recent and Fossil Honey Bees (Hymenoptera: Apidae; Apis). J. Hymenopt. Res. 1999, 8, 165–196. [Google Scholar]
- Garnery, L.; Mosshine, E.H.; Oldroyd, B.P.; Cornuet, J.M. Mitochondrial DNA Variation in Moroccan and Spanish Honey Bee Populations. Mol. Ecol. 1995, 4, 465–472. [Google Scholar] [CrossRef]
- DuPraw, E.J. Non-Linnean Taxonomy and the Systematics of Honeybees. Syst. Zool. 1965, 14, 1–24. [Google Scholar] [CrossRef]
- DuPraw, E.J. The Recognition and Handling of Honeybee Specimens in Non-Linnean Taxonomy. J. Apic. Res. 1965, 4, 71–84. [Google Scholar] [CrossRef]
- Kandemir, İ.; Özkan, A.; Fuchs, S. Reevaluation of Honeybee (Apis Mellifera) Microtaxonomy: A Geometric Morphometric Approach. Apidologie 2011, 42, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Barour, C.; Baylac, M. Geometric Morphometric Discrimination of the Three African Honeybee Subspecies Apis mellifera intermissa, A. m. sahariensis and A. m. capensis (Hymenoptera, Apidae): Fore Wing and Hind Wing Landmark Configurations. J. Hymenopt. Res. 2016, 52, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Abed, F.; Bachir-Bouiadjra, B.; Dahloum, L.; Yakubu, A.; Haddad, A.; Homrani, A. Procruste Analysis of Forewing Shape in Two Endemic Honeybee Subspecies Apis mellifera intermissa and A. m. sahariensis from the Northwest of Algeria. Biodiversitas J. Biol. Divers. 2021, 22, 154–164. [Google Scholar] [CrossRef]
- Baldensperger, J. North African Bees.—II. Bee World 1923, 5, 189–190. [Google Scholar] [CrossRef]
- Okuyama, H.; Hill, J.; Martin, S.J.; Takahashi, J. The Complete Mitochondrial Genome of a Buckfast Bee, Apis mellifera (Insecta: Hymenoptera: Apidae) in Northern Ireland. Mitochondrial DNA Part B 2018, 3, 338–339. [Google Scholar] [CrossRef] [Green Version]
- Chahbar, N.; Muñoz, I.; Dall’Olio, R.; De la Rúa, P.; Serrano, J.; Doumandji, S. Population Structure of North African Honey Bees Is Influenced by Both Biological and Anthropogenic Factors. J. Insect Conserv. 2013, 17, 385–392. [Google Scholar] [CrossRef]
- Loucif-Ayad, W.; Achou, M.; Legout, H.; Alburaki, M.; Garnery, L. Genetic Assessment of Algerian Honeybee Populations by Microsatellite Markers. Apidologie 2015, 46, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Cornuet, J.M.; Daoudi, A.; Mohssine, E.H.; Fresnaye, J. Etude biométrique de populations d’abeilles Marocaines. Apidologie 1988, 19, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Hepburn, H.R.; Radloff, S.E. Morphometric and Pheromonal Analyses of Apis mellifera L. along a Transect from the Sahara to the Pyrenees. Apidologie 1996, 27, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Hepburn, H.R.; Radloff, S.E. Biogeographical Correlates of Population Variance in the Honeybees (Apis mellifera L) of Africa. Apidologie 1997, 28, 243–258. [Google Scholar] [CrossRef] [Green Version]
- De la Rúa, P.; Radloff, S.; Hepburn, R.; Serrano, J. Do Molecular Markers Support Morphometric and Pheromone Analyses? A Preliminary Case Study in Apis mellifera Populations of Morocco. Arch. Zootec. 2007, 56, 33–42. [Google Scholar]
- Kandemir, I.; Kence, M.; Kence, A. Genetic and Morphometric Variation in Honeybee (Apis mellifera L.) Populations of Turkey. Apidologie 2000, 31, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Achou, M.; Loucif-Ayad, W.; Legout, H.; Hmidan, H.; Alburaki, M.; Garnery, L. An Insightful Molecular Analysis Reveals Foreign Honeybees Among Algerian Honeybee Populations (Apis mellifera L.). J. Data Min. Genom. Proteom. 2015, 6, 166. [Google Scholar] [CrossRef] [Green Version]
- Nawrocka, A.; Kandemir, İ.; Fuchs, S.; Tofilski, A. Computer Software for Identification of Honey Bee Subspecies and Evolutionary Lineages. Apidologie 2018, 49, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Miguel, I.; Baylac, M.; Iriondo, M.; Manzano, C.; Garnery, L.; Estonba, A. Both Geometric Morphometric and Microsatellite Data Consistently Support the Differentiation of the Apis mellifera M Evolutionary Branch. Apidologie 2011, 42, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Janczyk, A.; Meixner, M.D.; Tofilski, A. Morphometric Identification of the Endemic Maltese Honey Bee (Apis mellifera ruttneri). J. Apic. Res. 2020, 1–8. [Google Scholar] [CrossRef]
- Puškadija, Z.; Kovačić, M.; Raguž, N.; Lukić, B.; Prešern, J.; Tofilski, A. Morphological Diversity of Carniolan Honey Bee (Apis mellifera carnica) in Croatia and Slovenia. J. Apic. Res. 2020, 60, 326–336. [Google Scholar] [CrossRef]
- Francoy, T.M.; Wittmann, D.; Drauschke, M.; Müller, S.; Steinhage, V.; Bezerra-Laure, M.A.F.; De Jong, D.; Gonçalves, L.S. Identification of Africanized Honey Bees through Wing Morphometrics: Two Fast and Efficient Procedures. Apidologie 2008, 39, 488–494. [Google Scholar] [CrossRef] [Green Version]
- Tofilski, A. Using Geometric Morphometrics and Standard Morphometry to Discriminate Three Honeybee Subspecies. Apidologie 2008, 39, 558–563. [Google Scholar] [CrossRef] [Green Version]
- Crane, E. The World History of Beekeeping and Honey Hunting; Routledge: New York, NY, USA, 1999. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An Integrated Software Package for Geometric Morphometrics. Mol. Ecol. Res. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Dryden, I.L.; Mardia, K.V. Statistical Shape Analysis; John Wiley and Sons: Chichester, UK, 1998; Volume 4. [Google Scholar]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Franck, P.; Garnery, L.; Loiseau, A.; Oldroyd, B.P.; Hepburn, H.R.; Solignac, M.; Cornuet, J.-M. Genetic Diversity of the Honeybee in Africa: Microsatellite and Mitochondrial Data. Heredity 2001, 86, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Węgrzynowicz, P.; Gerula, D.; Tofilski, A.; Panasiuk, B.; Bieńkowska, M. Maternal Inheritance in Hybrids of Three Honey Bee Subspecies. J. Apic. Sci. 2019, 63, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Francoy, T.M.; Gonçalves, L.S.; De Jong, D. Rapid Morphological Changes in Populations of Hybrids between Africanized and European Honey Bees. Genet. Mol. Res. 2012, 11, 3349–3356. [Google Scholar] [CrossRef]
- Rinderer, T.E.; Daly, H.V.; Sylvester, H.A.; Collins, A.M.; Buco, S.M.; Hellmich, R.L.; Danka, R.G. Morphometric Differences Among Africanized and European Honey Bees and Their F1 Hybrids (Hymenoptera: Apidae). Ann. Entomol. Soc. Am. 1990, 83, 346–351. [Google Scholar] [CrossRef]
- Zhu, X.; Xu, X.; Zhou, S.; Wang, Q.; Chen, L.; Hao, Z.; Zhou, B. Low Temperature Exposure (20 °C) during the Sealed Brood Stage Induces Abnormal Venation of Honey Bee Wings. J. Apic. Res. 2018, 57, 458–465. [Google Scholar] [CrossRef]
- Dukku, U.H.; Danailu, G. An Appraisal of Subspecific Classification of Apis mellifera L. in Parts of West and Central Africa through Landmark-Based Geometric Morphometric Analysis of Forewings. J. Apic. Res. 2020, 59, 722–729. [Google Scholar] [CrossRef]
- Shaibi, T.; Fuchs, S.; Moritz, R.F.A. Morphological Study of Honeybees (Apis mellifera) from Libya. Apidologie 2009, 40, 97–105. [Google Scholar] [CrossRef]
- Barour, C.; Tahar, A.; Baylac, M. Forewing Shape Variation in Algerian Honey Bee Populations of Apis mellifera intermissa (Buttel-Reepen, 1906) (Hymenoptera: Apidae): A Landmark-Based Geometric Morphometrics Analysis. Afr. Entomol. 2011, 19, 11–22. [Google Scholar] [CrossRef]
- Büchler, R.; Costa, C.; Hatjina, F.; Andonov, S.; Meixner, M.D.; Conte, Y.L.; Uzunov, A.; Berg, S.; Bienkowska, M.; Bouga, M.; et al. The Influence of Genetic Origin and Its Interaction with Environmental Effects on the Survival of Apis mellifera L. Colonies in Europe. J. Apic. Res. 2014, 53, 205–214. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Errachidia | Ouarzazate | Tinghir | Zagora | A. m. intermissa | A. m. sahariensis |
|---|---|---|---|---|---|---|
| Errachidia | - | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Ouarzazate | 3.4948 | - | <0.0001 | <0.0001 | <0.0001 | 0.0001 |
| Tinghir | 3.2692 | 2.2285 | - | <0.0001 | <0.0001 | <0.0001 |
| Zagora | 4.1742 | 2.7590 | 2.6875 | - | <0.0001 | <0.0001 |
| A. m. intermissa | 8.2451 | 7.2371 | 6.5403 | 6.0323 | - | <0.0001 |
| A. m. sahariensis | 10.1513 | 10.0084 | 9.2805 | 8.5097 | 6.6533 | - |
| Measurement | Errachidia (±SD) | Ouarzazate (±SD) | Tinghir (±SD) | Zagora (±SD) | Mean of SE Morocco (±SD) | A. m. intermissa (±SD) | A. m. sahariensis (±SD) | P1 | P2 | DuPraw 1965a (±CI) |
|---|---|---|---|---|---|---|---|---|---|---|
| CuI | 2.53 ± 0.25 | 2.46 ± 0.29 | 2.58 ± 0.21 | 2.35 ± 0.24 | 2.48 ± 0.25 | 2.27 ± 0.18 | 2.39 ± 0.25 | 0.002 * | 0.730 | - |
| A1 | 26.63 ± 1.31 | 25.23 ± 1.59 | 26.02 ± 1.38 | 24.66 ± 1.7 | 25.64 ± 1.49 | 24.12 ± 2.3 | 24.53 ± 1.7 | 0.002 * | 0.460 | 22.7 ± 1.4 |
| A4 | 31.41 ± 1.21 | 32.08 ± 1.48 | 31.90 ± 1.08 | 32.03 ± 1.45 | 31.85 ± 1.30 | 31.58 ± 1.17 | 31.78 ± 1.82 | 0.610 | 0.980 | 31.1 ± 1.5 |
| B4 | 106.15 ± 2.67 | 105.35 ± 3.49 | 105.36 ± 2.82 | 104.43 ± 2.82 | 105.32 ± 2.95 | 103.12 ± 3.62 | 102.47 ± 4.6 | 0.018 * | 0.190 | 102 ± 4.3 |
| D7 | 99.65 ± 1.33 | 100.84 ± 1.58 | 99.96 ± 1.28 | 100.11 ± 2.00 | 100.14 ± 1.55 | 99.99 ± 2.14 | 100.59 ± 1.09 | 0.880 | 0.880 | - |
| E9 | 20.25 ± 0.62 | 20.56 ± 0.91 | 20.05 ± 0.64 | 20.35 ± 0.85 | 20.3 ± 0.75 | 19.95 ± 0.73 | 18.89 ± 0.34 | 0.160 | 0.001 * | 19.8 ± 0.8 |
| G7 | 23.61 ± 0.34 | 23.46 ± 0.63 | 23.51 ± 0.47 | 23.28 ± 0.71 | 23.46 ± 0.54 | 23.28 ± 0.66 | 22.89 ± 0.23 | 0.410 | 0.120 | 23.4 ± 0.8 |
| G18 | 99.62 ± 1.78 | 99.47 ± 2.01 | 99.88 ± 1.44 | 100.52 ± 1.92 | 99.87 ± 1.79 | 99.51 ± 2.44 | 99.14 ± 1.81 | 0.720 | 0.730 | 97.5 ± 1.2 |
| H12 | 18.73 ± 0.86 | 18.9 ± 0.97 | 18.66 ± 0.96 | 18.47 ± 1.1 | 18.69 ± 0.97 | 18.2 ± 1.55 | 17.51 ± 1.24 | 0.150 | 0.080 | 17.5 ± 0.9 |
| J10 | 51.48 ± 1.78 | 51.09 ± 1.48 | 51.55 ± 2.00 | 51.53 ± 1.88 | 51.41 ± 1.78 | 48.7 ± 2.87 | 49.96 ± 1.36 | 0.000 * | 0.319 | 54.1 ± 1.6 |
| J16 | 89.54 ± 1.69 | 89.7 ± 2.36 | 90.3 ± 1.94 | 90.28 ± 2.28 | 89.96 ± 2.07 | 93.74 ± 2.00 | 93.75 ± 2.55 | 0.000 * | 0.001 * | 97.2 ± 3.0 |
| K19 | 80.31 ± 1.36 | 79.73 ± 1.55 | 80.65 ± 1.81 | 78.76 ± 1.73 | 79.86 ± 1.61 | 77.79 ± 1.97 | 77.61 ± 2.13 | 0.000 * | 0.039 * | - |
| L13 | 17.32 ± 0.91 | 17.21 ± 0.89 | 17.04 ± 0.6 | 17.44 ± 0.87 | 17.25 ± 0.82 | 15.49 ± 0.92 | 16 ± 0.42 | 0.000 * | 0.010 * | - |
| M17 | 44.62 ± 1.18 | 45.53 ± 1.81 | 45.31 ± 1.33 | 45.07 ± 1.72 | 45.13 ± 1.51 | 44.27 ± 2.63 | 42.26 ± 1.02 | 0.090 | 0.003 * | 46.4 ± 2.0 |
| N23 | 79.92 ± 1.28 | 81.2 ± 1.98 | 81.57 ± 1.41 | 81.34 ± 1.48 | 81.01 ± 1.54 | 83.28 ± 2.4 | 84.2 ± 2.67 | 0.000 * | 0.003 * | 94 ± 1.9 |
| O26 | 36.03 ± 1.77 | 36.48 ± 2.28 | 37.12 ± 2.10 | 36.84 ± 2.4 | 36.62 ± 2.14 | 40.12 ± 2.62 | 35.08 ± 2.38 | 0.000 * | 0.340 | 37.4 ± 2.9 |
| Q21 | 34.79 ± 0.56 | 34.96 ± 0.67 | 35.16 ± 0.78 | 35.49 ± 0.94 | 35.10 ± 0.74 | 34.44 ± 1.06 | 34.35 ± 0.65 | 0.003 * | 0.160 | 35.9 ± 0.9 |
| LG | 1.99 ± 0.02 | 1.96 ± 0.03 | 1.97 ± 0.03 | 1.95 ± 0.03 | 1.97 ± 0.03 | 1.99 ± 0.05 | 1.99 ± 0.02 | 0.050 | 0.420 | 1.99 ± 0.03 |
| AO | 4.33 ± 0.04 | 4.28 ± 0.06 | 4.33 ± 0.05 | 4.28 ± 0.07 | 4.31 ± 0.06 | 4.42 ± 0.1 | 4.42 ± 0.05 | 0.000 * | 0.004 * | 4.44 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aglagane, A.; Tofilski, A.; Er-Rguibi, O.; Laghzaoui, E.-M.; Kimdil, L.; El Mouden, E.H.; Fuchs, S.; Oleksa, A.; Aamiri, A.; Aourir, M. Geographical Variation of Honey Bee (Apis mellifera L. 1758) Populations in South-Eastern Morocco: A Geometric Morphometric Analysis. Insects 2022, 13, 288. https://doi.org/10.3390/insects13030288
Aglagane A, Tofilski A, Er-Rguibi O, Laghzaoui E-M, Kimdil L, El Mouden EH, Fuchs S, Oleksa A, Aamiri A, Aourir M. Geographical Variation of Honey Bee (Apis mellifera L. 1758) Populations in South-Eastern Morocco: A Geometric Morphometric Analysis. Insects. 2022; 13(3):288. https://doi.org/10.3390/insects13030288
Chicago/Turabian StyleAglagane, Abdessamad, Adam Tofilski, Omar Er-Rguibi, El-Mustapha Laghzaoui, Latifa Kimdil, El Hassan El Mouden, Stefan Fuchs, Andrzej Oleksa, Ahmed Aamiri, and Mohamed Aourir. 2022. "Geographical Variation of Honey Bee (Apis mellifera L. 1758) Populations in South-Eastern Morocco: A Geometric Morphometric Analysis" Insects 13, no. 3: 288. https://doi.org/10.3390/insects13030288
APA StyleAglagane, A., Tofilski, A., Er-Rguibi, O., Laghzaoui, E. -M., Kimdil, L., El Mouden, E. H., Fuchs, S., Oleksa, A., Aamiri, A., & Aourir, M. (2022). Geographical Variation of Honey Bee (Apis mellifera L. 1758) Populations in South-Eastern Morocco: A Geometric Morphometric Analysis. Insects, 13(3), 288. https://doi.org/10.3390/insects13030288