
The Development and Evaluation of Insect Traps for the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae), Vector of Citrus Huanglongbing

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. ACP Traps
2.2. Stem Traps
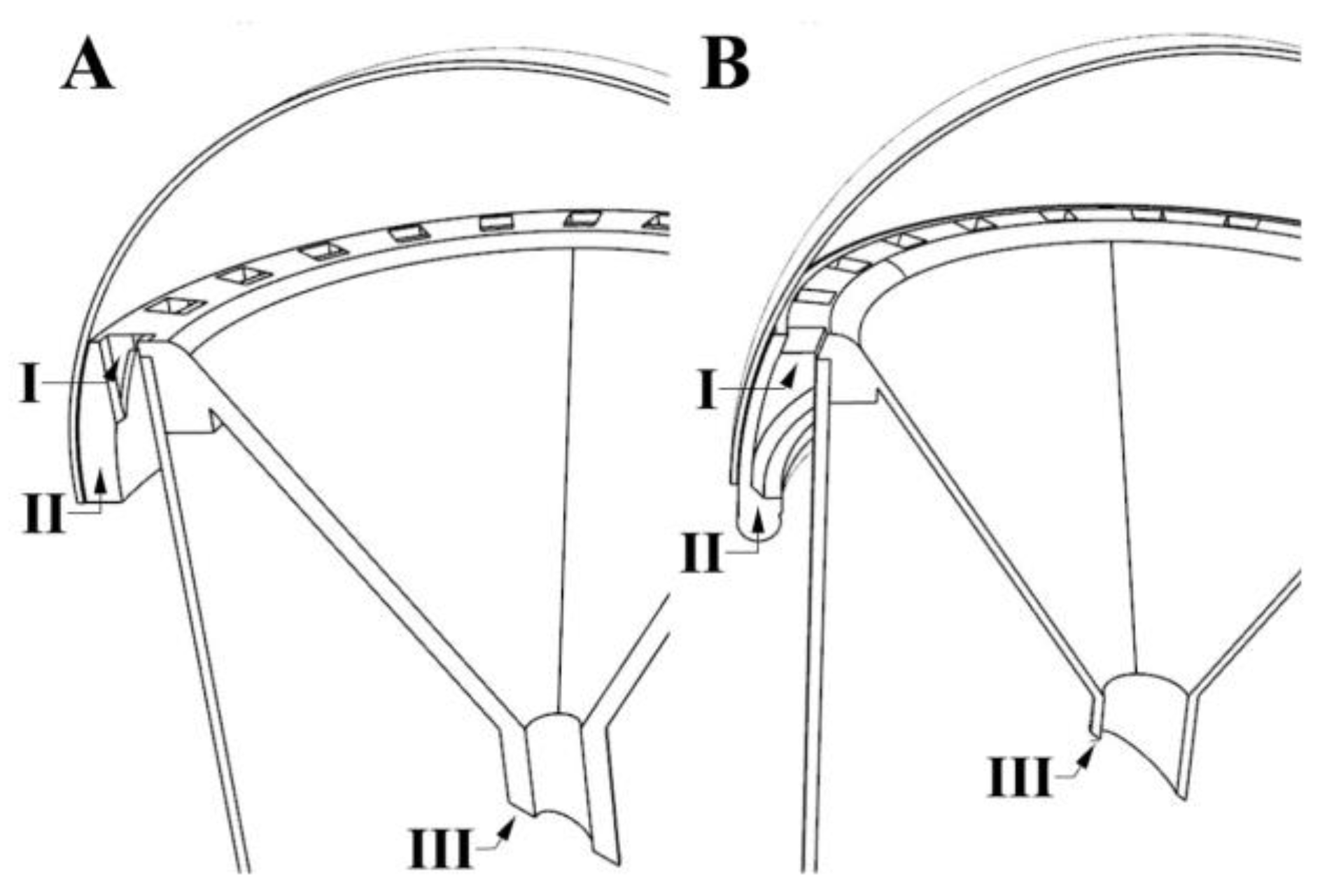
2.3. Cylinder Traps
2.4. 3D Design and Printing
2.5. Field Trials in Florida
2.6. Trap Evaluations in Field Cages in Pomona, CA
2.7. Trap Evaluations in Citrus Groves in Temecula, CA
2.8. By-Catch
3. Results
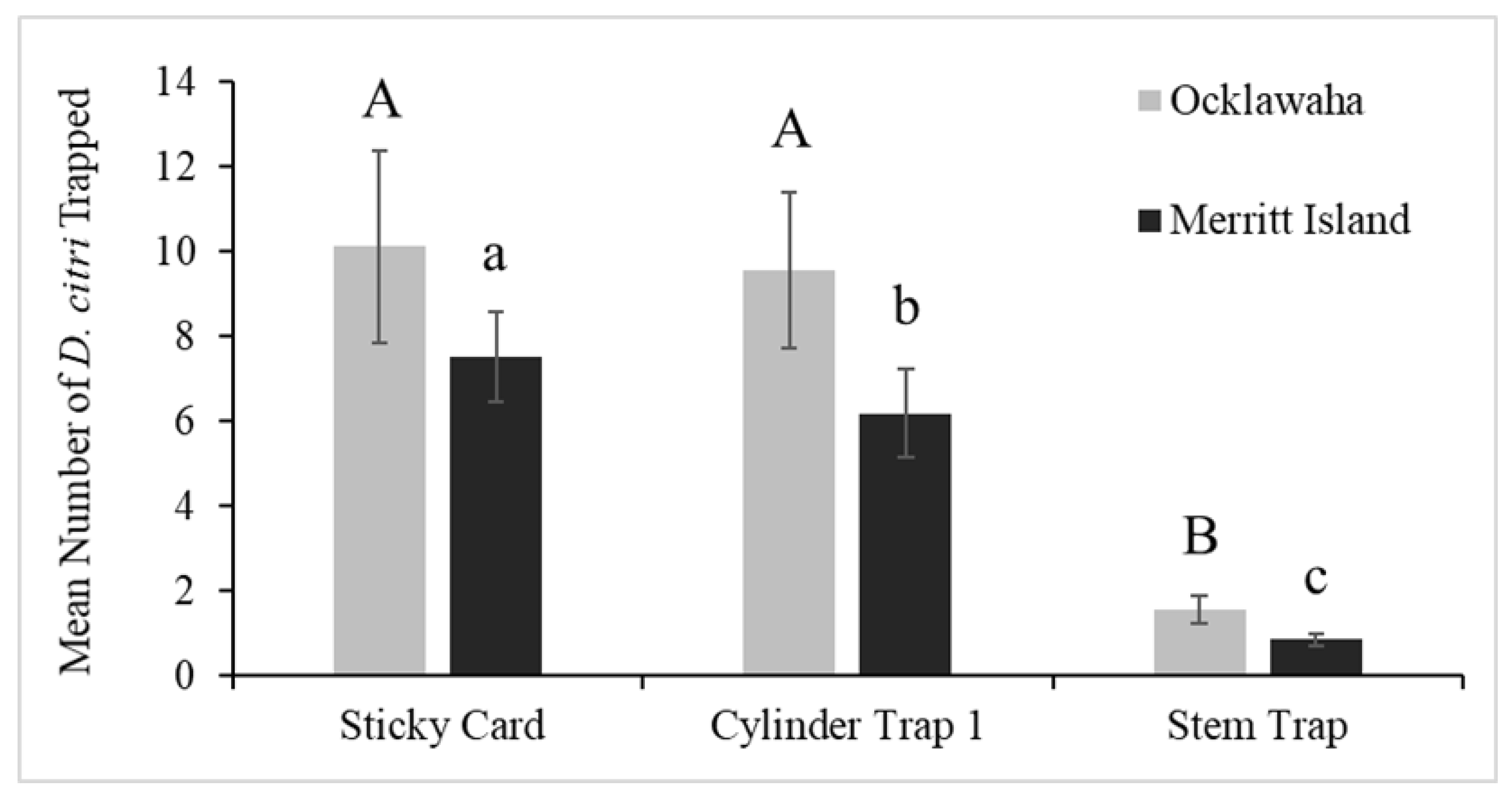
3.1. Field Trials in Merritt Island and Ocklawaha, Florida
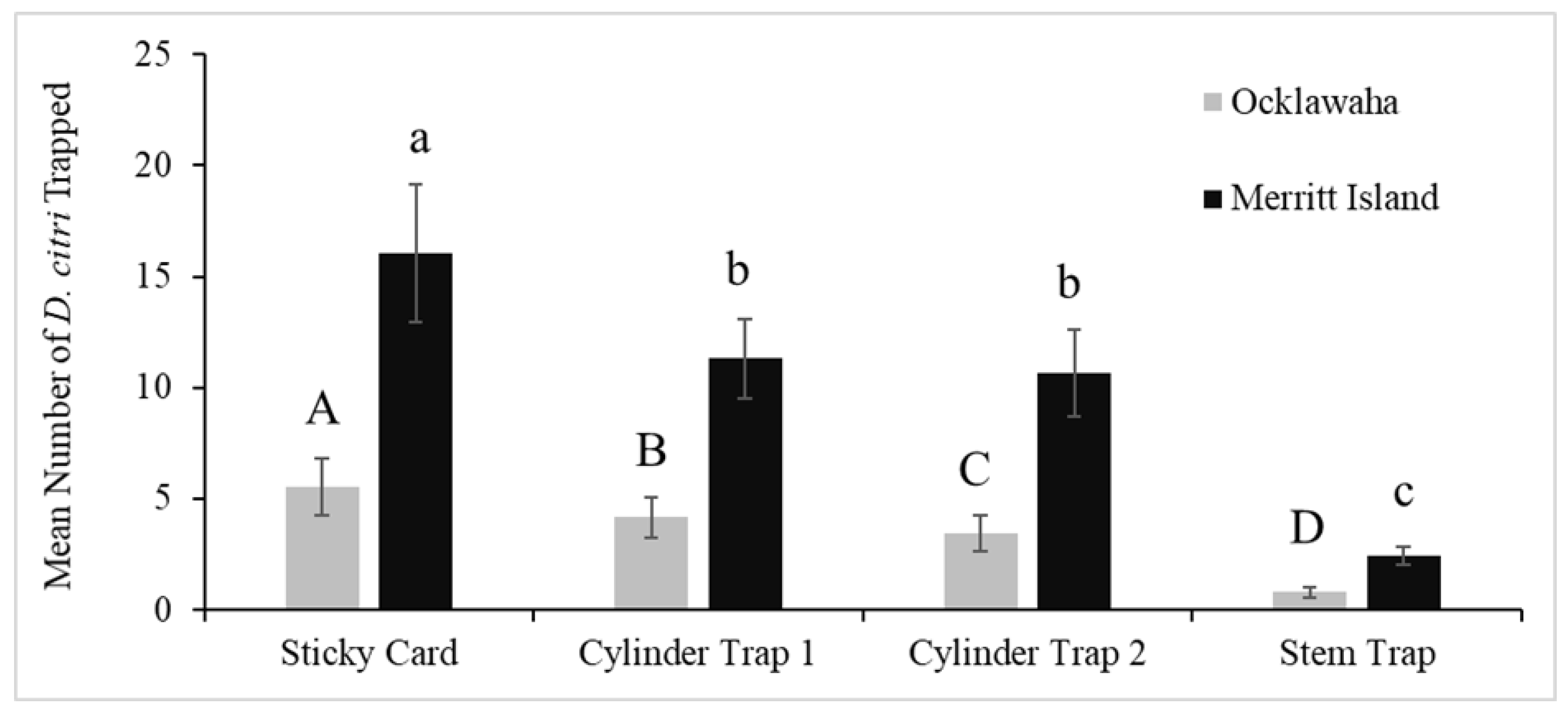
3.2. Trap Evaluations in Field Cages in Pomona, California
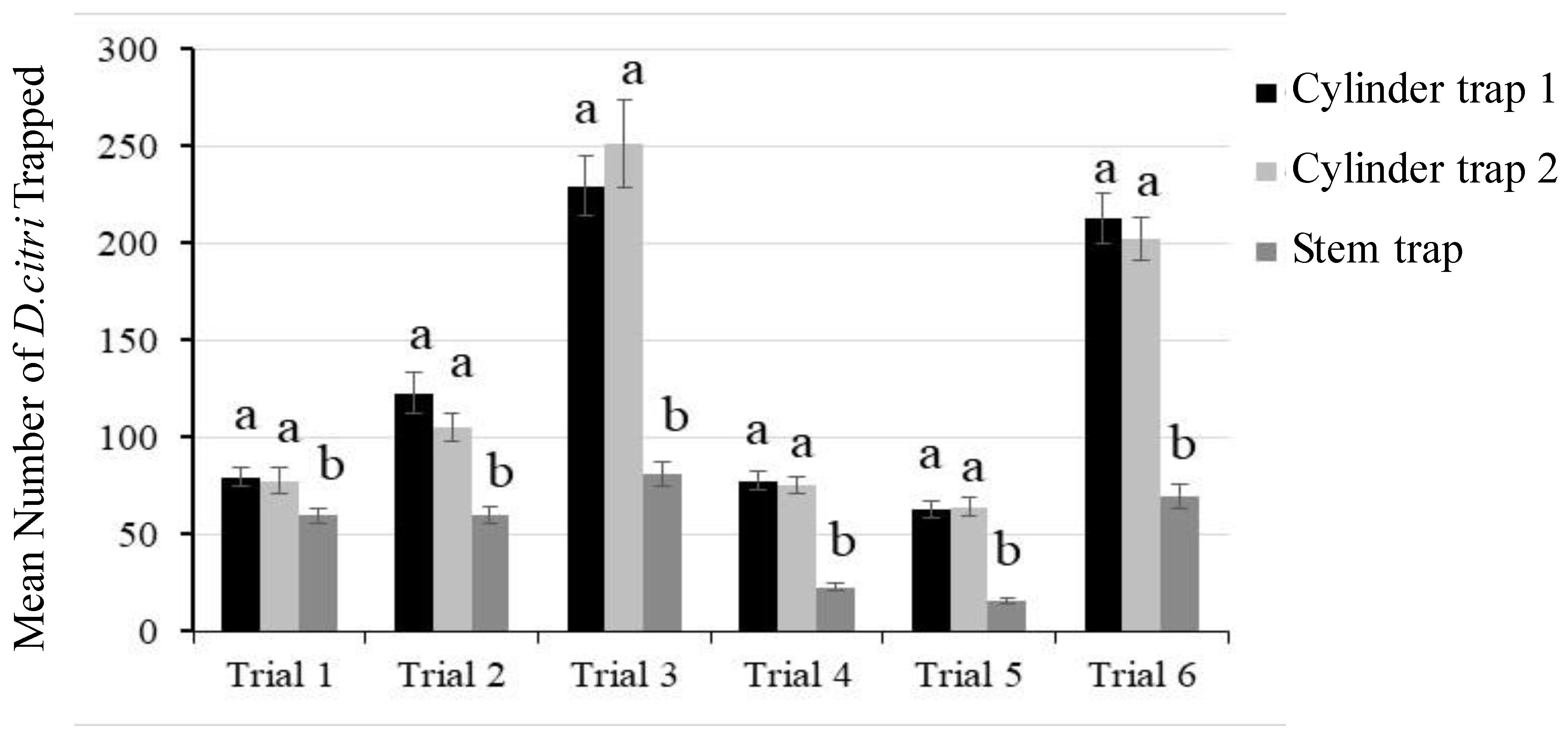
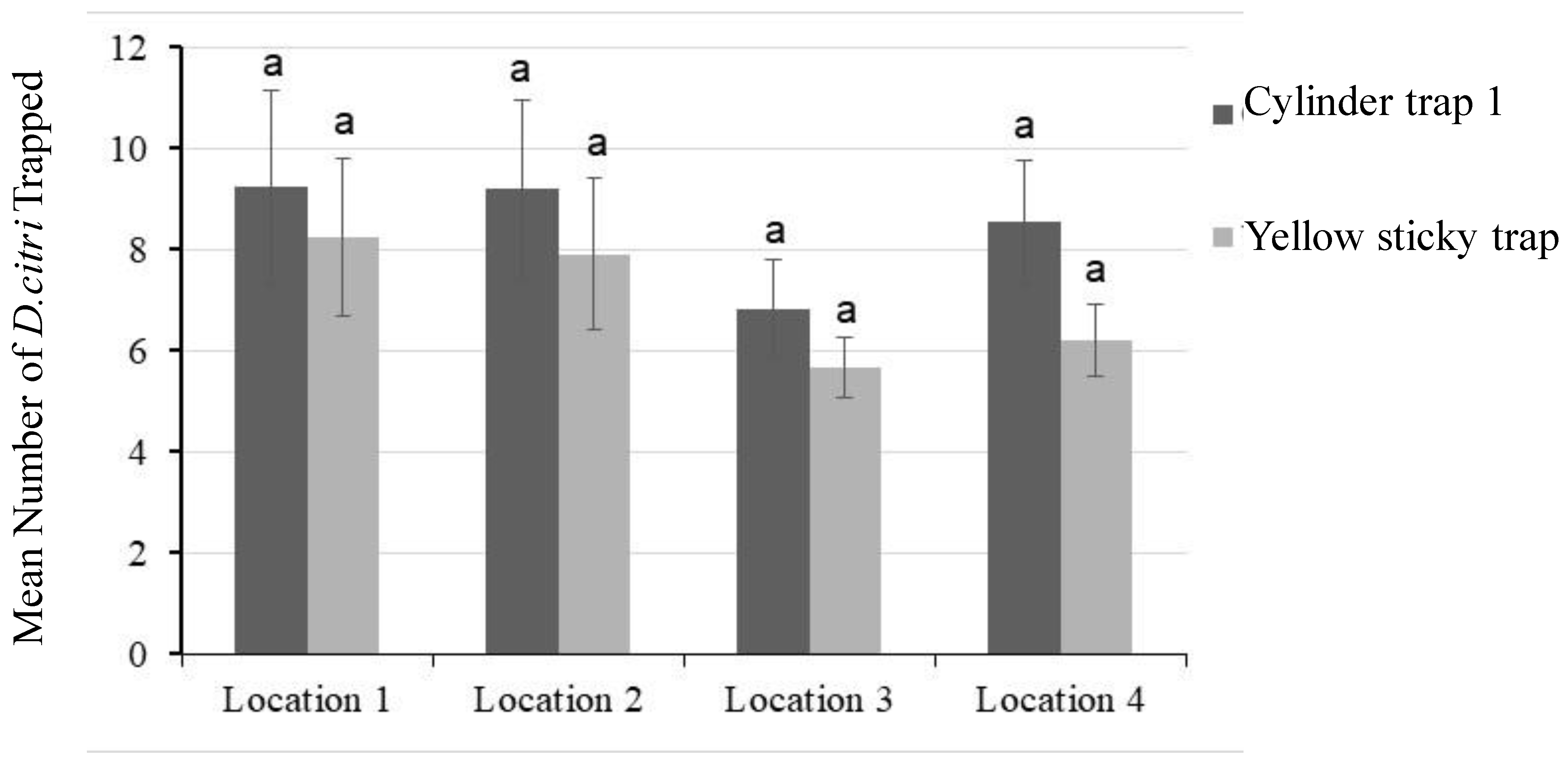
3.3. Evaluation of Cylinder Trap 1 in Citrus Groves in Temecula, CA
3.4. By-Catch
3.5. Cost of 3D-Printed Traps
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Regional Management of Huanglongbing (HLB) in Latin America and the Caribbean. Available online: http://www.fao.org/americas/perspectivas/hlb/en/ (accessed on 1 January 2022).
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar] [CrossRef]
- Coletta-Filho, H.D.; Targon, M.L.P.N.; Takita, M.A.; De Negri, J.D.; Pompeu, J.; Machado, M.A.; do Amaral, A.M.; Muller, G.W. First Report of the Causal Agent of Huanglongbing (“Candidatus Liberibacter asiaticus”) in Brazil. Plant Dis. 2004, 88, 1382. [Google Scholar] [CrossRef] [PubMed]
- Halbert, S.E. The discovery of Huanglongbing in Florida. In Proceedings of the International Citrus Canker Huanglongbing Research Workshop, Orlando, FL, USA, 7–11 November 2005; p. 50. [Google Scholar]
- USDA; NASS. Florida Citrus Statistics, 2018–2019; Florida Department of Agriculture and Consumer Services: Tallahassee, FL, USA, 2020; pp. 1–114.
- Singerman, A.; Burani-Arouca, M.; Futch, S.H. The Profitability of New Citrus Plantings in Florida in the Era of Huanglongbing. Hortscience 2018, 53, 1655–1663. [Google Scholar] [CrossRef] [Green Version]
- Singerman, A.; Rogers, M.E. The Economic Challenges of Dealing with Citrus Greening: The Case of Florida. J. Integr. Pest Manag. 2020, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- USDA; NASS. Florida Citrus Statistics, 2019–2020; Florida Department of Agriculture and Coosnumer Services: Tallahassee, FL, USA, 2021; pp. 1–114.
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumagai, L.B.; LeVesque, C.S.; Blomquist, C.L.; Madishetty, K.; Guo, Y.; Woods, P.W.; Rooney-Latham, S.; Rascoe, J.; Gallindo, T.; Schnabel, D.; et al. First Report of Candidatus Liberibacter asiaticus Associated with Citrus Huanglongbing in California. Plant Dis. 2013, 97, 283. [Google Scholar] [CrossRef] [PubMed]
- Burckhardt, D.; Ouvrard, D.; Percy, D.M. An updated classification of the jumping plant-lice (Hemiptera: Psylloidea) integrating molecular and morphological evidence. Eur. J. Entomol. 2021, 736, 1137–1182. [Google Scholar] [CrossRef]
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar] [CrossRef]
- da Graça, J.V.; Korsten, L. Citrus huanglongbing: Review, present status and future strategies. In Diseases of Fruits and Vegetables; Naqui, S.A.M.H., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; Volume I, pp. 229–245. [Google Scholar]
- Graham, J.; Gottwald, T.; Sétamou, M. Status of Huanglongbing (HLB) outbreaks in Florida, California and Texas. Trop. Plant Pathol. 2020, 45, 265–278. [Google Scholar] [CrossRef]
- McRoberts, N.; Figuera, S.G.; Olkowski, S.; McGuire, B.; Luo, W.; Posny, D.; Gottwald, T. Using models to provide rapid programme support for California’s efforts to suppress Huanglongbing disease of citrus. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180281. [Google Scholar] [CrossRef] [Green Version]
- Halbert, S.E.; Singer, B. Epidemiology of Huanglongbing: Implications of infective colonization events. In Asian Citrus Psyllid: Biology, Ecology and Management of the Huanglongbing Vector; Qureshi, J.A., Stansly, P.A., Eds.; CABI: London, UK, 2020; pp. 140–155. [Google Scholar]
- Lee, J.A.; Halbert, S.E.; Dawson, W.O.; Robertson, C.J.; Keesling, J.E.; Singer, B.H. Asymptomatic spread of huanglongbing and implications for disease control. Proc. Natl. Acad. Sci. USA 2015, 112, 7605–7610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelz-Stelinski, K.S.; Brlansky, R.H.; Ebert, T.A.; Rogers, M.E. Transmission parameters for Candidatus liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae). J. Econ. Entomol. 2010, 103, 1531–1541. [Google Scholar] [CrossRef] [PubMed]
- Halbert, S.E.; Manjunath, K.; Ramadugu, C.; Lee, R.F. Incidence of Huanglongbing-Associated ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri (Hemiptera: Psyllidae) Collected from Plants for Sale in Florida. Fla. Entomol. 2012, 95, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Manjunath, K.L.; Halbert, S.E.; Ramadugu, C.; Webb, S.; Lee, R.F. Detection of ‘Candidatus Liberibacter asiaticus’ in Diaphorina citri and Its Importance in the Management of Citrus Huanglongbing in Florida. Phytopathology 2008, 98, 387–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irey, M.S.; Gadea, P.; Hall, D.G. Detection of Candidatus Liberibacter asiaticus (Las) on yellow sticky traps by real-time PCR. In Proceedings of the 2nd International Research Conference on Huanglongbing, Orlando, FL, USA, 10–14 January 2011. [Google Scholar]
- Horton, D.R.; Miliczky, E.R.; Lewis, T.M.; Wohleb, C.H.; Waters, T.D.; Dickens, A.A.; Halbert, S.E.; Ramadugu, C.; Jensen, A.S. Building a better Psylloidea (Hemiptera) trap? A field-look at a prototype trap constructed using three-dimensional printer technology. Can. Entomol. 2019, 151, 115–129. [Google Scholar] [CrossRef]
- Sétamou, M.; Saldaña, R.R.; Hearn, J.M.; Dale, J.; Feria Arroyo, T.P.; Czokajlo, D. Screening Sticky Cards as a Simple Method for Improving Efficiency of Diaphorina citri (Hemiptera: Liviidae) Monitoring and Reducing Nontarget Organisms. J. Econ. Entomol. 2019, 112, 1167–1174. [Google Scholar] [CrossRef]
- Wentz, K.M.; Cooper, W.R.; Horton, D.R.; Wohleb, C.H.; Waters, T.D.; Halbert, S.E.; Ramadugu, C.; Snyder, J.; Kao, R.M. Prototype 3D-Printed Traps Capture Bactericera cockerelli (Šulc) (Hemiptera: Triozidae) Directly into Preservative for Improved Detection of “Candidatus Liberibacter solanacearum”. J. Entomol. Sci. 2020, 55, 147–155. [Google Scholar] [CrossRef]
- Action Plan for Asian Citrus Psyllid and Huanglongbing (Citrus Greening) in California. Available online: https://www.cdfa.ca.gov/citruscommittee/docs/ActionPlan.pdf (accessed on 1 March 2022).
- Hall, D.G.; Ramadugu, C.; Halbert, S.E. Efficacy of propylene glycol and salt for preserving ‘Ca. Liberibacter asiaticus’ in infected adult Asian Citrus Psyllids captured in outdoor traps. In Proceedings of the 92nd Annual Meeting of the South Eastern Branch of Entomological Society of America, Orlando, FL, USA, 4–7 March 2018; p. 47. [Google Scholar]
- Hall, D.G.; Sétamou, M.; Mizell, R.F. A comparison of sticky traps for monitoring Asian citrus psyllid (Diaphorina citri Kuwayama). Crop Prot. 2010, 29, 1341–1346. [Google Scholar] [CrossRef]
- Sétamou, M.; Sanchez, A.; Saldaña, R.R.; Patt, J.M.; Summy, R. Visual Responses of Adult Asian Citrus Psyllid (Hemiptera: Liviidae) to Colored Sticky Traps on Citrus Trees. J. Insect Behav. 2014, 27, 540–553. [Google Scholar] [CrossRef]
- Przyborowski, J.; Wilenski, H. Homogeneity of results in testing samples from Poisson series: With an application to testing clover seed for dodder. Biometrika 1940, 31, 313–323. [Google Scholar] [CrossRef]
- R-Core-Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 30 July 2018).
- Mann, H.B.; Whitney, D.R. On a Test of Whether one of Two Random Variables is Stochastically Larger than the Other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Sétamou, M.; Flores, D.; French, J.V.; Hall, D.G. Dispersion patterns and sampling plans for Diaphorina citri (Hemiptera: Psyllidae) in citrus. J. Econ. Entomol. 2008, 101, 1478–1487. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, J.A.; Stansly, P.A. Integrated approaches for managing the Asian citrus psyllid Diaphorina citri (Homoptera: Psyllidae) in Florida. Proc. Fla. State Hort. Soc. 2007, 120, 110–115. [Google Scholar]
- Aubert, B.; Hua, X.Y. Monitoring flight activity of Diaphorina citri on citrus and Murraya canopies. In Proceedings of the 4th International Asia Pacific Conference on Citrus Rehabilitation, Chiang Mai, Thailand, 4–10 February 1990. [Google Scholar]
- Miranda, M.P.; dos Santos, F.L.; Bassanezi, R.B.; Montesino, L.H.; Barbosa, J.C.; Sétamou, M. Monitoring methods for Diaphorina citri Kuwayama (Hemiptera: Liviidae) on citrus groves with different insecticide application programmes. J. Appl. Entomol. 2018, 142, 89–96. [Google Scholar] [CrossRef]
- Monzo, C.; Arevalo, H.A.; Jones, M.M.; Vanaclocha, P.; Croxton, S.D.; Qureshi, J.A.; Stansly, P.A. Sampling Methods for Detection and Monitoring of the Asian Citrus Psyllid (Hemiptera: Psyllidae). Environ. Entomol. 2015, 44, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.B. Comparison of Insect Vacuums for Sampling Asian Citrus Psyllid (Homoptera: Psyllidae) on Citrus Trees. Southwest. Entomol. 2012, 37, 55–60. [Google Scholar] [CrossRef]
- Stansly, P.A.; Qureshi, J.; Averalo, A. Why, When and how to monitor and manage Asian citrus psyllid. Citrus Ind. 2009, 90, 24–25. [Google Scholar]
- Flores, D.; Hall, D.G.; Jenkins, D.A.; Sétamou, M. Abundance of Asian Citrus Psyllid on Yellow Sticky Traps in Florida, Puerto Rico, and Texas Citrus Groves. Southwest. Entomol. 2009, 34, 1–11. [Google Scholar] [CrossRef]
- Hall, D.G.; Hentz, M.G.; Ciomperlik, M.A. A comparison of traps and stem tap sampling for monitoring adult Asian citrus psyllid (Hemiptera: Psyllidae) in citrus. Fla. Entomol. 2007, 90, 327–334. [Google Scholar] [CrossRef]
- Razi, M.F.; Keremane, M.L.; Ramadugu, C.; Roose, M.; Khan, I.A.; Lee, R.F. Detection of citrus huanglongbing-associated ‘Candidatus Liberibacter asiaticus’ in citrus and Diaphorina citri in Pakistan, seasonal variability, and implications for disease management. Phytopathology 2014, 104, 257–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, A.; Czokajlo, D.; Patt, J.M.; Sétamou, M. Development and Field Validation of a Beta-cyfluthrin-Based ‘Attract-and-Kill’ Device for Suppression of Asian Citrus Psyllid (Hemiptera: Liviidae) on Residential Citrus. J. Econ. Entomol. 2019, 112, 2824–2832. [Google Scholar] [CrossRef] [PubMed]
- Amelia, H. Wohlers Report 2021: Weakened Growth for Additive Manufacturing. Available online: https://www.3dnatives.com/en/wohlers-report-2021-180320214/#! (accessed on 2 January 2021).
- Berry, D.; Selby, R.D.; Horvath, J.C.; Cameron, R.H.; Porqueras, D.; Stouthamer, R. A Modular System of 3D Printed Emergence Traps for Studying the Biology of Shot Hole Borers and Other Scolytinae. J. Econ. Entomol. 2016, 109, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Domingue, M.J.; Pulsifer, D.P.; Lakhtakia, A.; Berkebile, J.; Steiner, K.C.; Lelito, J.P.; Hall, L.P.; Baker, T.C. Detecting emerald ash borers (Agrilus planipennis) using branch traps baited with 3D-printed beetle decoys. J. Pest Sci. 2015, 88, 267–279. [Google Scholar] [CrossRef]
- Qing, X.; Sánchez-Monge, A.; Bert, W. Three-dimensional modelling and printing as tools to enhance education and research in Nematology. Nematology 2015, 17, 1245–1248. [Google Scholar] [CrossRef] [Green Version]
- Will, K.; Steplowski, I. A 3D printed Malaise trap head. Pan-Pac. Entomol. 2016, 92, 86–91. [Google Scholar] [CrossRef]
- Garlotta, D. A Literature Review of Poly(Lactic Acid). J. Polym. Environ. 2001, 9, 63–84. [Google Scholar] [CrossRef]
- Vargas, R.I.; Stark, J.D.; Kido, M.H.; Ketter, H.M.; Whitehand, L.C. Methyl eugenol and cue-lure traps for suppression of male oriental fruit flies and melon flies (Diptera: Tephritidae) in Hawaii: Effects of lure mixtures and weathering. J. Econ. Entomol. 2000, 93, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Patt, J.M.; Sétamou, M. Responses of the Asian citrus psyllid to volatiles emitted by the flushing shoots of its rutaceous host plants. Environ. Entomol. 2010, 39, 618–624. [Google Scholar] [CrossRef] [Green Version]
- Bian, L.; Cai, X.-M.; Luo, Z.-X.; Li, Z.-Q.; Chen, Z.-M. Decreased capture of natural enemies of pests in light traps with light-emitting diode technology. Ann. Appl. Biol. 2018, 173, 251–260. [Google Scholar] [CrossRef]
- Paris, T.M.; Allan, S.A.; Udell, B.J.; Stansly, P.A. Wavelength and Polarization Affect Phototaxis of the Asian Citrus Psyllid. Insects 2017, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Sétamou, M.; Sanchez, A.; Patt, J.M.; Nelson, S.D.; Jifon, J.; Louzada, E.S. Diurnal Patterns of Flight Activity and Effects of Light on Host Finding Behavior of the Asian Citrus Psyllid. J. Insect Behav. 2012, 25, 264–276. [Google Scholar] [CrossRef]
- Shimoda, M.; Honda, K. Insect reactions to light and its applications to pest management. Appl. Entomol. Zool. 2013, 48, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, A.; Anfora, G.; Lucchi, A.; Lanzo, F.; Virant-Doberlet, M.; Mazzoni, V. Exploitation of insect vibrational signals reveals a new method of pest management. PLoS ONE 2012, 7, e32954. [Google Scholar] [CrossRef]
- Hartman, E.; Rohde, B.; Lujo, S.; Dixon, M.; McNeill, S.; Mankin, R.W. Behavioral Responses of Male Diaphorina citri (Hemiptera: Liviidae) to Mating Communication Signals from Vibration Traps in Citrus (Sapindales: Rutaceae) Trees. Fla. Entomol. 2017, 100, 767–771. [Google Scholar] [CrossRef] [Green Version]
- Lujo, S.; Hartman, E.; Norton, K.; Pregmon, E.A.; Rohde, B.B.; Mankin, R.W. Disrupting Mating Behavior of Diaphorina citri (Liviidae). J. Econ. Entomol. 2016, 109, 2373–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mankin, R.W.; Rohde, B.; McNeill, S. Vibrational duetting mimics to trap and disrupt mating of the devastating Asian citrus psyllid insect pest. Proc. Meet. Acoust. 2015, 25, 010006. [Google Scholar] [CrossRef] [Green Version]
- Rohde, B.; Paris, T.M.; Heatherington, E.M.; Hall, D.G.; Mankin, R.W. Responses of Diaphorina citri (Hemiptera: Psyllidae) to Conspecific Vibrational Signals and Synthetic Mimics. Ann. Entomol. Soc. Am. 2013, 106, 392–399. [Google Scholar] [CrossRef] [Green Version]
- Aksenov, A.A.; Martini, X.; Zhao, W.; Stelinski, L.L.; Davis, C.E. Synthetic blends of volatile, phytopathogen-induced odorants can be used to manipulate vector behavior. Front. Ecol. Evol. 2014, 2, 78. [Google Scholar] [CrossRef] [Green Version]
- Coutinho-Abreu, I.V.; Forster, L.; Guda, T.; Ray, A. Odorants for surveillance and control of the Asian Citrus Psyllid (Diaphorina citri). PLoS ONE 2014, 9, e109236. [Google Scholar] [CrossRef]
- Zanardi, O.Z.; Volpe, H.X.L.; Luvizotto, R.A.G.; Magnani, R.F.; Gonzalez, F.; Calvo, C.; Oehlschlager, C.A.; Lehan, B.J.; Esperanca, V.; Delfino, J.Y.; et al. Laboratory and field evaluation of acetic acid-based lures for male Asian citrus psyllid, Diaphorina citri. Sci. Rep. 2019, 9, 12920. [Google Scholar] [CrossRef] [Green Version]
- Onagbola, E.O.; Meyer, W.L.; Boina, D.R.; Stelinski, L.L. Morphological characterization of the antennal sensilla of the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), with reference to their probable functions. Micron 2008, 39, 1184–1191. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arthropod Groups Identified | Mean Number Captured per Cylinder Trap |
|---|---|
| Parasitic Hymenoptera | 13.324 |
| Diptera | 13.169 |
| Formicidae | 9.634 |
| Diaphorina citri | 9.084 |
| Hemiptera (except D. citri) | 6.676 |
| Thysanoptera | 5.648 |
| Coleoptera | 4.972 |
| Acari | 0.958 |
| Araneae | 0.535 |
| Lepidoptera | 0.479 |
| Neuroptera | 0.155 |
| Psocoptera | 0.042 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snyder, J.; Dickens, K.L.; Halbert, S.E.; Dowling, S.; Russell, D.; Henderson, R.; Rohrig, E.; Ramadugu, C. The Development and Evaluation of Insect Traps for the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae), Vector of Citrus Huanglongbing. Insects 2022, 13, 295. https://doi.org/10.3390/insects13030295
Snyder J, Dickens KL, Halbert SE, Dowling S, Russell D, Henderson R, Rohrig E, Ramadugu C. The Development and Evaluation of Insect Traps for the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae), Vector of Citrus Huanglongbing. Insects. 2022; 13(3):295. https://doi.org/10.3390/insects13030295
Chicago/Turabian StyleSnyder, James, Katrina L. Dickens, Susan E. Halbert, Stefanie Dowling, Dyrana Russell, Ruth Henderson, Eric Rohrig, and Chandrika Ramadugu. 2022. "The Development and Evaluation of Insect Traps for the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae), Vector of Citrus Huanglongbing" Insects 13, no. 3: 295. https://doi.org/10.3390/insects13030295
APA StyleSnyder, J., Dickens, K. L., Halbert, S. E., Dowling, S., Russell, D., Henderson, R., Rohrig, E., & Ramadugu, C. (2022). The Development and Evaluation of Insect Traps for the Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae), Vector of Citrus Huanglongbing. Insects, 13(3), 295. https://doi.org/10.3390/insects13030295