
Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley

 , , ,
, , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Barley Samples
2.3. Fumigation Procedures with Ozone
2.4. Assessment of Insect Mortality
2.5. Germination Test
2.6. Grain Quality Measurement
2.7. Statistical Analysis
3. Results
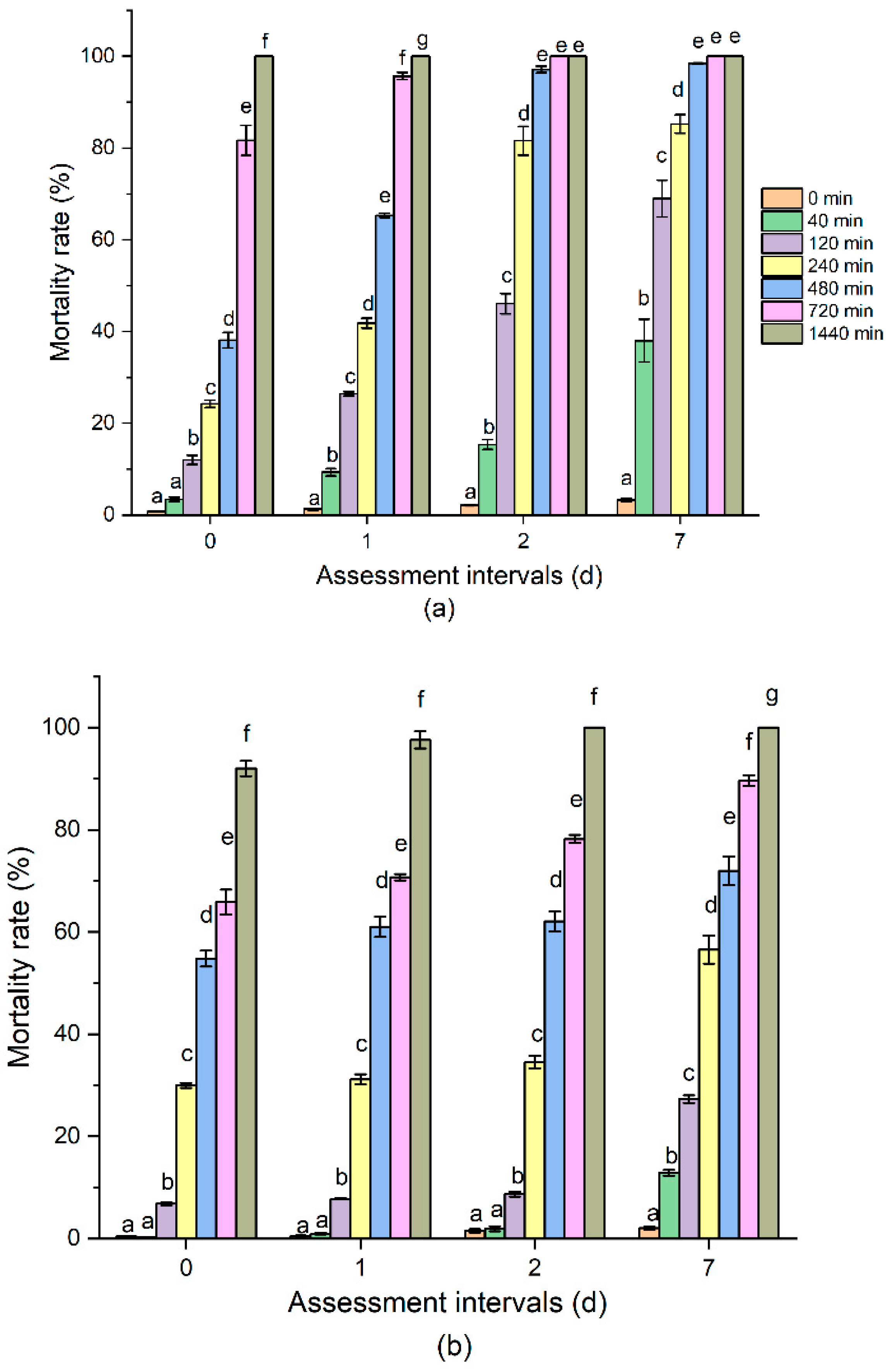
3.1. Toxicity of O3 on R. dominica and T. castaneum Adults at Different Assessment Intervals
3.2. Toxicity of O3 Duration on All Life Stages of R. dominica and T. castaneum
3.3. Effect of O3 Treatment Duration on Germination of Barley
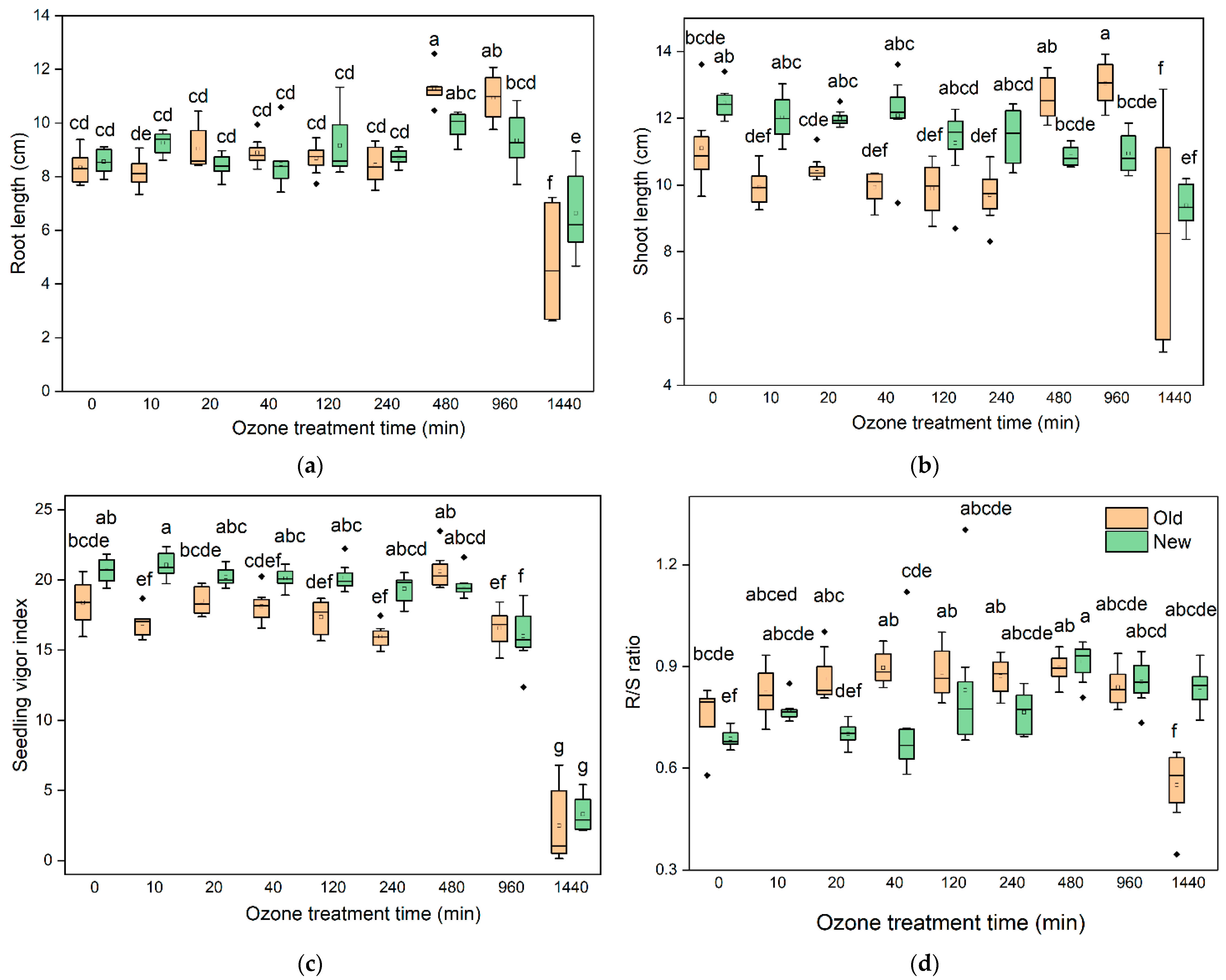
3.4. Effect of O3 Treatment Duration on Seedling Growth
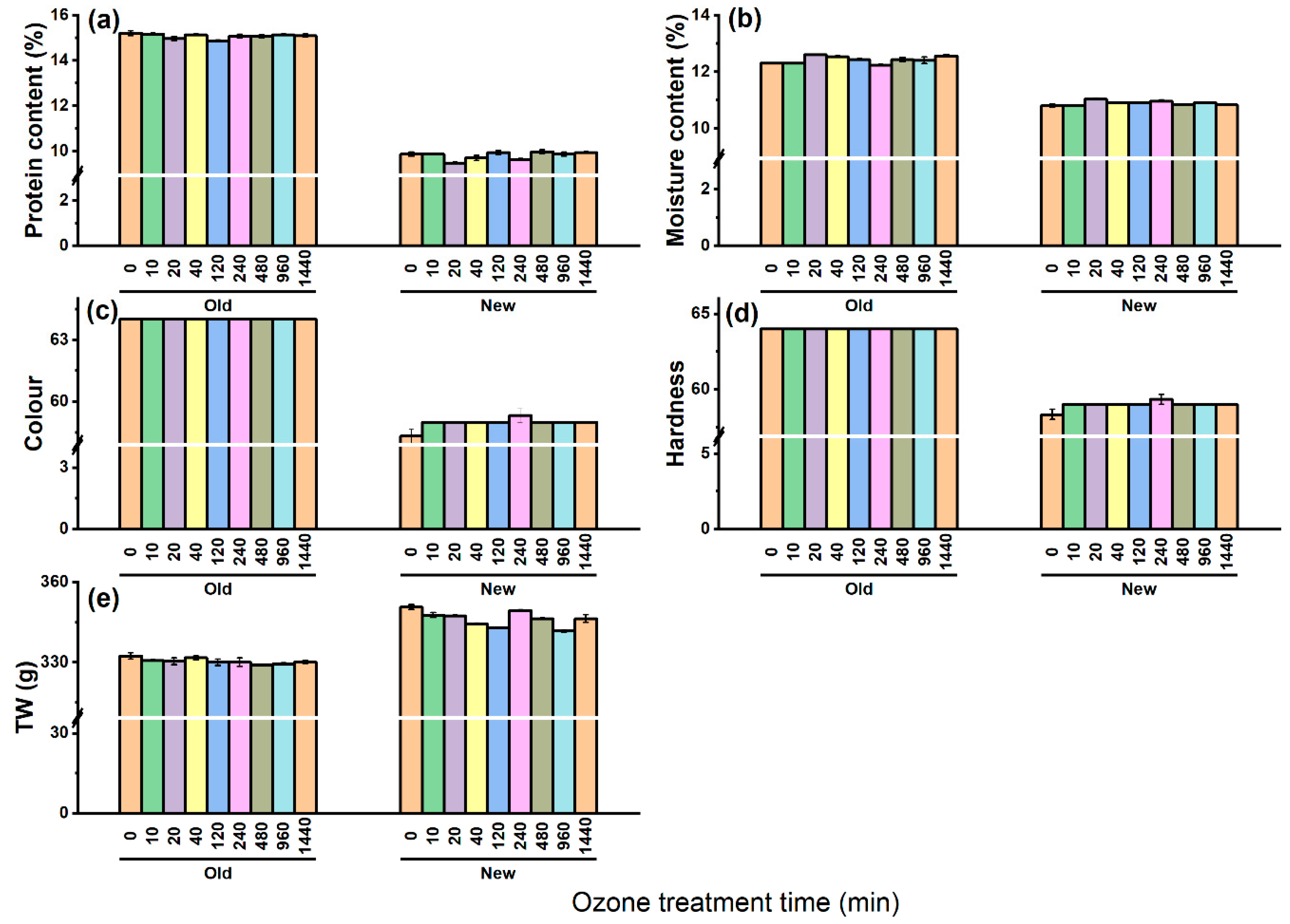
3.5. Effect of O3 on Barley Grain Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mesterházy, Á.; Oláh, J.; Popp, J. Losses in the grain supply chain: Causes and solutions. Sustainability 2020, 12, 2342. [Google Scholar] [CrossRef] [Green Version]
- Shankar, U.; Abrol, D.P. Integrated Pest Management in Stored Grains. In Integrated Pest Management: Principles and Practice; CABI: Wallingford, UK, 2012; pp. 386–407. [Google Scholar]
- Banks, H. In Behaviour of gases in grain storages, Fumigation and Controlled Atmosphere Storage of Grain. In Proceedings of the International Conference, Singapore, 4–18 February 1989; pp. 96–107. [Google Scholar]
- Bond, E.J.; Monro, H.A.U. Manual of Fumigation for Insect Control; FAO: Rome, Italy, 1984; Volume 54. [Google Scholar]
- Nayak, M.K.; Daglish, G.J.; Phillips, T.W.; Ebert, P.R. Resistance to the fumigant phosphine and its management in insect pests of stored products: A global perspective. Annu. Rev. Entomol. 2020, 65, 333–350. [Google Scholar] [CrossRef] [Green Version]
- Collins, P.J. Resistance to grain protectants and fumigants in insect pests of stored products in Australia. In Proceedings of the Australian Postharvest Technical Conference, Canberra, Australia, 25–27 June 2003; pp. 55–57. [Google Scholar]
- Cato, A.; Elliott, B.; Nayak, M.K.; Phillips, T.W. Geographic variation in phosphine resistance among North American populations of the red flour beetle (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2017, 110, 1359–1365. [Google Scholar] [CrossRef]
- Gautam, S.; Opit, G.; Hosoda, E. Phosphine resistance in adult and immature life stages of Tribolium castaneum (Coleoptera: Tenebrionidae) and Plodia interpunctella (Lepidoptera: Pyralidae) populations in California. J. Econ. Entomol. 2016, 109, 2525–2533. [Google Scholar] [CrossRef]
- Opit, G.; Phillips, T.W.; Aikins, M.J.; Hasan, M. Phosphine resistance in Tribolium castaneum and Rhyzopertha dominica from stored wheat in Oklahoma. J. Econ. Entomol. 2012, 105, 1107–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Li, F.; Liu, M.; Wang, Y.; Shen, F.; Tang, P. Susceptibility of Tribolium castaneum to phosphine in China and functions of cytochrome P450s in phosphine resistance. J. Pest Sci. 2019, 92, 1239–1248. [Google Scholar] [CrossRef]
- Song, X.; Wang, P.; Zhang, H. Phosphine resistance in Rhyzopertha dominica (Fabricius) (Coleoptera: Bostrichidae) from different geographical populations in China. Afr. J. Biotechnol. 2011, 10, 16367–16373. [Google Scholar]
- Wakil, W.; Kavallieratos, N.G.; Usman, M.; Gulzar, S.; El-Shafie, H.A. Detection of phosphine resistance in field populations of four key stored-grain insect pests in Pakistan. Insects 2021, 12, 288. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, S.; Nayak, K.R.; Anjum, S.S. The action of phosphine against the eggs of phosphine-resistant and-susceptible strains of Rhyzopertha dominica F. Pest Manag. Sci. Former. Pestic. Sci. 2001, 57, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, S. Phosphine resistance in stored grain insect pests in India. In Proceedings of the 7th International Working Conference on Stored-Product Protection, Berlin, Germany, 7–11 October 2018; pp. 14–19. [Google Scholar]
- Finkelman, S.; Lendler, E.; Navarro, S.; Navarro, H.; Ashbell, G. New prospects for ethyl formate as a fumigant for the date industry. In Proceedings of the 10th International Working Conference on Stored Product Protection, Estoril, Portugal, 27 June–2 July 2010; pp. 359–364. [Google Scholar]
- Athanassiou, C.; Korunic, Z.; Kavallieratos, N.; Peteinatos, G.; Boukouvala, M.; Mikeli, N. New trends in the use of diatomaceous earth against stored-grain insects. In Proceedings of the 9th International Working Conference on Stored Product Protection, Campinas, Brazil, 15 October 2006; pp. 730–740. [Google Scholar]
- Korunic, Z.; Cenkowski, S.; Fields, P. Grain bulk density as affected by diatomaceous earth and application method. Postharvest Biol. Technol. 1998, 13, 81–89. [Google Scholar] [CrossRef]
- Ziaee, M.; Ebadollahi, A.; Wakil, W. Integrating inert dusts with other technologies in stored products protection. Toxin Rev. 2021, 40, 404–419. [Google Scholar] [CrossRef]
- Kumar, H.; Vijay, V.K.; Subbarao, P.M.; Chandra, R. Studies on the application of bio-carbon dioxide as controlled atmosphere on pest management in wheat grain storage. J. Stored Prod. Res. 2022, 95, 101911. [Google Scholar] [CrossRef]
- Batta, Y.A.; Kavallieratos, N.G. The use of entomopathogenic fungi for the control of stored-grain insects. Int. J. Pest Manag. 2018, 64, 77–87. [Google Scholar] [CrossRef]
- Singh, D.; Raina, T.K.; Singh, J. Entomopathogenic fungi: An effective biocontrol agent for management of insect populations naturally. J. Pharm. Sci. Res. 2017, 9, 833. [Google Scholar]
- Rumbos, C.I.; Athanassiou, C.G. Use of entomopathogenic fungi for the control of stored-product insects: Can fungi protect durable commodities? J. Pest Sci. 2017, 90, 839–854. [Google Scholar] [CrossRef]
- Sutar, S.A.; Thirumdas, R.; Chaudhari, B.B.; Deshmukh, R.R.; Annapure, U.S. Effect of cold plasma on insect infestation and keeping quality of stored wheat flour. J. Stored Prod. Res. 2021, 92, 101774. [Google Scholar] [CrossRef]
- Carocho, M.; Antonio, A.L.; Barreira, J.C.; Rafalski, A.; Bento, A.; Ferreira, I.C. Validation of gamma and electron beam irradiation as alternative conservation technology for European chestnuts. Food Bioprocess Technol. 2014, 7, 1917–1927. [Google Scholar] [CrossRef] [Green Version]
- Navarro, S. The use of modified and controlled atmospheres for the disinfestation of stored products. J. Pest Sci. 2012, 85, 301–322. [Google Scholar] [CrossRef]
- Dufresne, S.; Hewitt, A.; Robitaille, S. Ozone sterilization: Another option for healthcare in the 21st century. Am. J. Infect. Control. 2004, 32, E26–E27. [Google Scholar] [CrossRef]
- Isikber, A.A.; Athanassiou, C.G. The use of ozone gas for the control of insects and micro-organisms in stored products. J. Stored Prod. Res. 2015, 64, 139–145. [Google Scholar] [CrossRef]
- Rice, R.G.; Graham, D.M.; Lowe, M.T. Recent ozone applications in food processing and sanitation. Food Saf. Mag. 2002, 8, 10–17. [Google Scholar]
- Carletti, L.; Botondi, R.; Moscetti, R.; Stella, E.; Monarca, D.; Cecchini, M.; Massantini, R. Use of ozone in sanitation and storage of fresh fruits and vegetables. J. Food Agric. Environ. 2013, 11, 585–589. [Google Scholar]
- FDA, United States Food and Drug Administration. Rules and Regulations, Part 173-Secondary Direct Food Additives Permitted in Food for Human Consumption (21 CFR Part 173 Authority: 21 USC. 321, 342, 348). Fed. Regist. 2001, 66, 123. [Google Scholar]
- Sadeghi, R.; Mirabi Moghaddam, R.; Taghizadeh, M. Application of ozone to control dried fig pests—Oryzaephilus surinamensis (Coleoptera: Silvanidae) and Ephestia kuehniella (Lepidoptera: Pyralidae)—And its organoleptic properties. J. Econ. Entomol. 2017, 110, 2052–2055. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.; Wu, J.; Doan, H. Inactivation of fungi associated with barley grain by gaseous ozone. J. Environ. Sci. Health Part B 2003, 38, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Bonjour, E.; Opit, G.; Hardin, J.; Jones, C.; Payton, M.; Beeby, R. Efficacy of ozone fumigation against the major grain pests in stored wheat. J. Econ. Entomol. 2011, 104, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Savi, G.D.; Piacentini, K.C.; Bittencourt, K.O.; Scussel, V.M. Ozone treatment efficiency on Fusarium graminearum and deoxynivalenol degradation and its effects on whole wheat grains (Triticum aestivum L.) quality and germination. J. Stored Prod. Res. 2014, 59, 245–253. [Google Scholar] [CrossRef]
- Sousa, A.D.; Faroni, L.D.A.; Guedes, R.; Tótola, M.; Urruchi, W. Ozone as a management alternative against phosphine-resistant insect pests of stored products. J. Stored Prod. Res. 2008, 44, 379–385. [Google Scholar] [CrossRef]
- McDonough, M.X.; Mason, L.J.; Woloshuk, C.P. Susceptibility of stored product insects to high concentrations of ozone at different exposure intervals. J. Stored Prod. Res. 2011, 47, 306–310. [Google Scholar] [CrossRef]
- Mishra, G.; Palle, A.A.; Srivastava, S.; Mishra, H.N. Disinfestation of stored wheat grain infested with Rhyzopertha dominica by ozone treatment: Process optimization and impact on grain properties. J. Sci. Food Agric. 2019, 99, 5008–5018. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Faroni, L.; Sousa, A.; Prates, L.; Abreu, A. Kinetics of the ozone gas reaction in popcorn kernels. J. Stored Prod. Res. 2019, 83, 168–175. [Google Scholar] [CrossRef]
- Pandiselvam, R.; Thirupathi, V.; Anandakumar, S. Reaction kinetics of ozone gas in paddy grains. J. Food Process Eng. 2015, 38, 594–600. [Google Scholar] [CrossRef]
- Subramanyam, B.; Xinyi, E.; Savoldelli, S.; Sehgal, B. Efficacy of ozone against Rhyzopertha dominica adults in wheat. J. Stored Prod. Res. 2017, 70, 53–59. [Google Scholar] [CrossRef]
- Zhu, F. Effect of ozone treatment on the quality of grain products. Food Chem. 2018, 264, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Dodd, J.G.; Vegi, A.; Vashisht, A.; Tobias, D.; Schwarz, P.; Wolf-Hall, C.E. Effect of ozone treatment on the safety and quality of malting barley. J. Food Prot. 2011, 74, 2134–2141. [Google Scholar] [CrossRef]
- Tiwari, B.; Brennan, C.S.; Curran, T.; Gallagher, E.; Cullen, P.; O’Donnell, C. Application of ozone in grain processing. J. Cereal Sci. 2010, 51, 248–255. [Google Scholar] [CrossRef]
- Jian, F.; Jayas, D.S.; White, N.D. Can ozone be a new control strategy for pests of stored grain? Agric. Res. 2013, 2, 1–8. [Google Scholar] [CrossRef]
- Matsue, Y.; Uchimura, Y.; Sato, H. Confirmation of the productivity of the stored seeds of wheat and two-rowed barley. Plant Prod. Sci. 2002, 5, 187–193. [Google Scholar] [CrossRef] [Green Version]
- ISTA. International Rules for Seed Testing; International Seed Testing Association: Basserdorf, Switzerland, 2006. [Google Scholar]
- Islam, A.; Anuar, N.; Yaakob, Z. Effect of genotypes and pre-sowing treatments on seed germination behavior of Jatropha. Asian J. Plant Sci. 2009, 8, 433. [Google Scholar] [CrossRef]
- Pasynkov, A.; Pasynkova, E. Comparative evaluation of different methods to determine the crude gluten content in wheat grain. Russ. Agric. Sci. 2021, 47, 238–243. [Google Scholar] [CrossRef]
- Pojić, M.; Mastilović, J.; Majcen, N. The application of near infrared spectroscopy in wheat quality control. In Infrared Spectroscopy–Life and Biomedical Sciences; Theophanides, T., Ed.; Intech Publisher: Rijeka, Croatia, 2012; pp. 167–184. [Google Scholar]
- Subramanyam, B.; Li, B. Efficacy of ozone against phosphine susceptible and resistant strains of four stored-product insect species. Insects 2017, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Holmstrup, M.; Sørensen, J.G.; Heckmann, L.-H.; Slotsbo, S.; Hansen, P.; Hansen, L.S. Effects of ozone on gene expression and lipid peroxidation in adults and larvae of the red flour beetle (Tribolium castaneum). J. Stored Prod. Res. 2011, 47, 378–384. [Google Scholar] [CrossRef]
- Boopathy, B.; Rajan, A.; Radhakrishnan, M. Ozone: An Alternative Fumigant in Controlling the Stored Product Insects and Pests: A Status Report. Ozone Sci. Eng. 2022, 44, 79–95. [Google Scholar] [CrossRef]
- Ballinger, C.A.; Cueto, R.; Squadrito, G.; Coffin, J.F.; Velsor, L.W.; Pryor, W.A.; Postlethwait, E.M. Antioxidant-mediated augmentation of ozone-induced membrane oxidation. Free Radic. Biol. Med. 2005, 38, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Xinyi, E.; Li, B.; Subramanyam, B. Efficacy of ozone against adults and immature stages of phosphine susceptible and resistant strains of Rhyzopertha dominica. J. Stored Prod. Res. 2019, 83, 110–116. [Google Scholar]
- Pimentel, M.A.G.; Faroni, L.R.D.A.; Tótola, M.R.; Guedes, R.N.C. Phosphine resistance, respiration rate and fitness consequences in stored-product insects. Pest Manag. Sci. Former. Pestic. Sci. 2007, 63, 876–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emekci, M.; Navarro, S.; Donahaye, E.; Rindner, M.; Azrieli, A. Respiration of Rhyzopertha dominica (F.) at reduced oxygen concentrations. J. Stored Prod. Res. 2004, 40, 27–38. [Google Scholar] [CrossRef]
- Wu, J.; Doan, H.; Cuenca, M.A. Investigation of gaseous ozone as an anti-fungal fumigant for stored wheat. J. Chem. Technol. Biotechnol. Int. Res. Process Environ. Clean Technol. 2006, 81, 1288–1293. [Google Scholar] [CrossRef]
- Shrestha, R.K.; Lindsey, L.E. Agronomic management of malting barley and research needs to meet demand by the craft brew industry. Agron. J. 2019, 111, 1570–1580. [Google Scholar] [CrossRef] [Green Version]
- Monroy Vazquez, M.E.; Peña-Valdivia, C.B.; García, J.R.; Solano, E.; Campos, H.; García, E. Chemical scarification and ozone in seed dormancy alleviation of wild and domesticated Opuntia, Cactaceae. Ozone Sci. Eng. 2017, 39, 104–114. [Google Scholar] [CrossRef]
- Wang, H.; Qi, Q.; Schorr, P.; Cutler, A.J.; Crosby, W.L.; Fowke, L.C. ICK1, a cyclin-dependent protein kinase inhibitor from Arabidopsis thaliana interacts with both Cdc2a and CycD3, and its expression is induced by abscisic acid. Plant J. 1998, 15, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Normov, D.; Chesniuk, E.; Shevchenko, A.; Normova, T.; Goldman, R.; Pozhidaev, D.; Bohinc, T.; Trdan, S. Does ozone treatment of maize seeds influence their germination and growth energy? Acta Agric. Slov. 2019, 114, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Gapper, C.; Dolan, L. Control of plant development by reactive oxygen species. Plant Physiol. 2006, 141, 341–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bláha, L. Possible Experimental Mistakes in Agricultural Research. Agric. Sci. 2019, 10, 1423–1437. [Google Scholar] [CrossRef] [Green Version]
- Alexander, L.; Yuri, S.; Mikhail, P.; Olga, S.; Sergey, K.; Irina, L. Treatment of spring wheat seeds by ozone generated from humid air and dry oxygen. Res. Agric. Eng. 2018, 64, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Bonifas, K.D.; Walters, D.T.; Cassman, K.G.; Lindquist, J.L. Nitrogen supply affects root: Shoot ratio in corn and velvetleaf (Abutilon theophrasti). Weed Sci. 2005, 53, 670–675. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Shao, H.; Luo, X.; Wang, R.; Li, Y.; Li, Y.; Luo, Y.; Chen, Z. Effect of ozone treatment on deoxynivalenol and wheat quality. PLoS ONE 2016, 11, e0147613. [Google Scholar] [CrossRef] [PubMed]
- Sui, Z.; Yao, T.; Zhong, J.; Li, Y.; Kong, X.; Ai, L. Ozonation treatment improves properties of wheat flour and the baking quality of cake. Philipp. Agric. Sci. 2016, 99, 50–57. [Google Scholar]
- Lozano-Isla, F.; Campos, M.L.; Endres, L.; Bezerra-Neto, E.; Pompelli, M.F. Effects of seed storage time and salt stress on the germination of Jatropha curcas L. Ind. Crops Prod. 2018, 118, 214–224. [Google Scholar] [CrossRef]
- Azevedo Neto, A.D.; Prisco, J.T.; Gomes-Filho, E. Changes in soluble amino-N, soluble proteins and free amino acids in leaves and roots of salt-stressed maize genotypes. J. Plant Interact. 2009, 4, 137–144. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Insects | DAT † | Slope ‡ ± SE | Intercept ± SE | Relative Median Potency * | LT50 (95% CI §) (min) | LT95 (95% CI) (min) | χ2 ¶ | df |
|---|---|---|---|---|---|---|---|---|
| R. dominica | 0 | 2.32 ± 0.05 | −6.21 ± 0.15 | 1.00 | 472.91 (343.04–653.55) | 2419.93 (1465.95–6336.68) | 118.02 | 6 |
| 1 | 2.48 ± 0.06 | −6.48 ± 0.15 | 1.15 | 410.12 (278.81–587.64) | 1887.41 (1130.49–5594.71) | 162.50 | 6 | |
| 2 | 2.74 ± 0.06 | −7.01 ± 0.17 | 1.25 | 378.37 (265.87–515.29) | 1510.96 (976.73–3617.63) | 138.20 | 6 | |
| 7 | 2.06 ± 0.05 | −4.80 ± 0.12 | 2.22 | 213.50 (146.17–291.40) | 1342.06 (843.67–3033.99) | 95.58 | 6 | |
| T. castaneum | 0 | 2.72 ± 0.07 | −7.23 ± 0.20 | 1.00 | 453.07 (228.14–769.72) | 1823.12 (972.31–27304.4) | 322.13 | 6 |
| 1 | 2.31 ± 0.06 | −5.54 ± 0.14 | 1.80 | 252.04 (143.69–378.75) | 1300.80 (747.48–4884.78) | 198.01 | 6 | |
| 2 | 2.85 ± 0.07 | −5.88 ± 0.16 | 3.91 | 115.79 (92.92–139.38) | 437.12 (341.45–620.62) | 40.70 | 6 | |
| 7 | 2.30 ± 0.07 | −4.15± 0.14 | 7.12 | 63.60 (49.06–78.09) | 329.40 (256.77–463.99) | 27.22 | 6 |
| Life Stage | O3 Treatment Time (min) | Rhyzopertha dominica | Tribolium castaneum |
|---|---|---|---|
| Mortality Rate (%) † | Mortality Rate (%) | ||
| Egg | 0 | 0.00 ± 0.00 g | 1.67 ± 1.67 g |
| 40 | 18.33 ± 3.33 fg | 17.92 ± 1.5 fg | |
| 120 | 38.33 ± 3.33 ef | 33.33 ± 3.85 ef | |
| 240 | 68.33 ± 4.41 bcd | 49.01 ± 5.21 de | |
| 480 | 70.63 ± 5.23 bc | 61.52 ± 9.34 cd | |
| 960 | 83.33 ± 6.01 ab | 81.11 ± 2.22 abc | |
| 1440 | 100.00 ± 0.00 a | 100.00 ± 0.00 a | |
| Larvae | 0 | 3.33 ± 1.92 g | 3.70 ± 0.93 g |
| 40 | 26.67 ± 3.33 f | 25.56 ± 2.94 f | |
| 120 | 53.33 ± 6.67 e | 51.38 ± 2.01 e | |
| 240 | 60.42 ± 5.51 de | 82.88 ± 1.93 bc | |
| 480 | 70.83 ± 2.08 cd | 99.67 ± 0.33 a | |
| 960 | 91.67 ± 2.08 ab | 100.00 ± 0.00 a | |
| 1440 | 100.00 ± 0.00 a | 100.00 ± 0.00 a | |
| Pupae | 0 | 0.00± 0.00 g | 2.78 ± 1.60 g |
| 40 | 12.95 ± 2.50 fg | 24.39 ± 3.43 ef | |
| 120 | 28.62 ± 3.10 de | 51.82 ± 4.30 cd | |
| 240 | 44.21 ± 4.89 c | 64.55 ± 2.92 b | |
| 480 | 63.82 ± 4.20 b | 74.24 ± 2.98 b | |
| 960 | 74.30 ± 1.86 b | 93.33 ± 3.33 a | |
| 1440 | 100.00 ± 0.00 a | 100.00 ± 0.00 a | |
| Adults | 0 | 2.06 ± 0.32 h | 3.21 ± 0.61 gh |
| 40 | 12.85 ± 0.58 g | 38.00 ± 8.09 e | |
| 120 | 27.29 ± 0.77 f | 68.97 ± 6.90 c | |
| 240 | 56.57 ± 2.74 d | 85.24 ± 3.51 b | |
| 480 | 71.95 ± 2.81 c | 98.45 ± 0.29 a | |
| 960 | 89.62 ± 1.06 ab | 100.00 ± 0.00 a | |
| 1440 | 100 ± 0.00 a | 100 ± 0.00 a |
| O3 Treatment Duration (min) | Germination Rate (%) | |
|---|---|---|
| Old (2013/2014) † | New (2019/2020) | |
| 0 | 94.34 ± 1.67 ab | 98.35 ± 0.56 a |
| 10 | 94.11 ± 0.97 ab | 99.06 ± 0.36 a |
| 20 | 95.28 ± 0.87 ab | 99.06 ± 0.36 a |
| 40 | 97.17 ± 1.01 a | 98.59 ± 0.59 a |
| 120 | 95.28 ± 1.01 ab | 98.82 ± 0.79 a |
| 240 | 84.21 ± 1.12 bc | 96.23 ± 0.71 a |
| 480 | 86.32 ± 0.93 c | 94.34 ± 0.94 a |
| 960 | 68.16 ± 1.61 d | 78.54 ± 2.71 b |
| 1440 | 14.39 ± 4.55 e | 20.28 ± 1.84 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, X.; Agarwal, M.; Xiao, Y.; Ren, Y.; Maker, G.; Yu, X. Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley. Insects 2022, 13, 318. https://doi.org/10.3390/insects13040318
Dong X, Agarwal M, Xiao Y, Ren Y, Maker G, Yu X. Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley. Insects. 2022; 13(4):318. https://doi.org/10.3390/insects13040318
Chicago/Turabian StyleDong, Xue, Manjree Agarwal, Yu Xiao, Yonglin Ren, Garth Maker, and Xiangyang Yu. 2022. "Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley" Insects 13, no. 4: 318. https://doi.org/10.3390/insects13040318
APA StyleDong, X., Agarwal, M., Xiao, Y., Ren, Y., Maker, G., & Yu, X. (2022). Ozone Efficiency on Two Coleopteran Insect Pests and Its Effect on Quality and Germination of Barley. Insects, 13(4), 318. https://doi.org/10.3390/insects13040318