
Effects of Elevated CO2 on the Fitness of Three Successive Generations of Lipaphis erysimi


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Closed-Dynamics CO2 Chamber
2.2. Host Plants and L. erysimi Stock
2.3. L. erysimi Feeding
2.4. Life Table Analysis and Population Projection of L. erysimi
3. Results
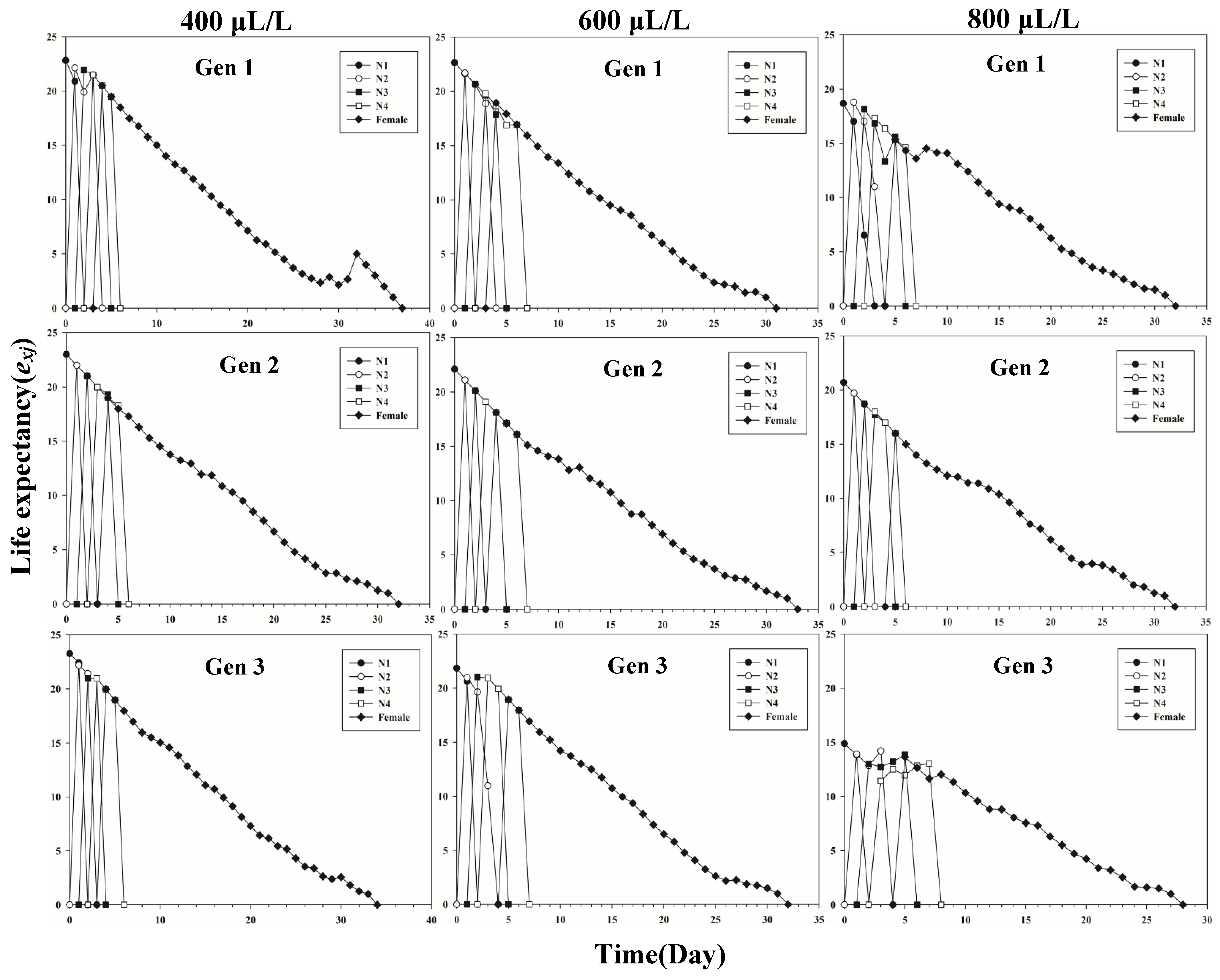
3.1. Life History Parameters of L. erysimi
3.2. Population Parameters of L. erysimi
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fukayama, H.; Sugino, M.; Fukude, T.; Masumoto, C.; Taniguchi, Y.; Okada, M.; Sameshima, R.; Hatanaka, T.; Misso, S.; Hasegawa, T.; et al. Gene expression profiling of roce grown in free air CO2 enrichment. Oecologia 2011, 104, 454–466. [Google Scholar]
- Seneweera, S.; Makino, A.; Hirotu, N.; Norton, R.; Suzuki, Y. New insight into photosynthetic acclimation to elevated CO2: The role of leaf nitrogen and ribulose-1, 5-bisphosphate carboxylase/oxygenase content in rice leaves. Environ. Exp. Bot. 2011, 71, 128–136. [Google Scholar] [CrossRef]
- Integovernmental Panel on Climate Change. The Physical Science Basis; IPCC Fifth Assessment Report; IPCC, 2013; Available online: http://www.ipcc.ch/report/ar5/wg1/ (accessed on 20 March 2022).
- Woodward, F.I. Global change: Translating plant ecophysiological responses to ecosystems. Trends Ecol. Evol. 1990, 5, 308–311. [Google Scholar] [CrossRef]
- Hartley, S.E.; Jones, C.G.; Couper, G.C.; Jones, T.H. Biosynthesis of plant phenolic compounds in elevated atmospheric CO2. Glob. Chang. Biol. 2000, 6, 497–506. [Google Scholar] [CrossRef]
- Kitao, M.S.; Komatsu, M.; Yazaki, K.C.; Kitaoka, S.S.; Tobita, H. Growth overcompensation against O3 exposure in two Japanese oak species, Quercus mongolica var. crispula and Quercus serrata, grown under elevated CO2. Environ. Pollut. 2015, 206, 133–141. [Google Scholar] [CrossRef]
- Novriyanti, E.; Watanabe, M.; Kitao, M.S.; Utsugi, H.; Uemura, A.; Koike, T.S. High nitrogen and elevated [CO2] effects on the growth, defense and photosynthetic performance of two eucalypt species. Environ. Pollut. 2012, 170, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Chen, F.J.; Ge, F. Impacts of early-season square abscission on the growth and yield of transgenic Bt cotton under elevated CO2. Field Crops Res. 2007, 102, 239–243. [Google Scholar] [CrossRef]
- Fitzgerald, G.J.; Tausz, M.; O’Leary, G.J.; Mollah, M.R.; Tausz-Posch, S.; Seneweera, S.; Mock, I.; Löw, M.; Partington, D.L.; Mcneil, D.; et al. Elevated atmospheric [CO2] can dramatically increase wheat yields in semi-arid environments and buffer against heat waves. Glob. Chang. Biol. 2016, 22, 2269–2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.H.; Zheng, X.X.; Liu, J.P.; Zeng, Y.Y.; Yang, F.L.; Wu, G. Time-dependent stress evidence in dynamic allocation of physiological metabolism of Nilaparvata lugens in response to elevated CO2. Environ. Pollut. 2020, 265, 114767. [Google Scholar] [CrossRef]
- Yan, H.Y.; Guo, H.G.; Sun, Y.C.; Ge, F. Plant phenolics mediated bottom-up effects of elevated CO2 on Acyrthosiphon pisum and its parasitoid Aphidius avenae. Insect Sci. 2020, 27, 170–184. [Google Scholar] [CrossRef]
- Robinson, E.A.; Ryan, G.D.; Newman, J.A. A meta-analytical review of the effects of elevated CO2 on plant–arthropod interactions highlights the importance of interacting environmental and biological variables. New Phytol. 2012, 194, 321–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.X.; Wu, G. How does the arthropod-plant system respond to abrupt and gradual increases in atmospheric CO2? J. Integr. Agric. 2020, 4, 1156–1160. [Google Scholar] [CrossRef]
- Liu, J.P.; Huang, W.K.; Chi, H.; Wang, C.H.; Hua, H.X.; Wu, G. Effects of elevated CO2 on the fitness and potential population damage of Helicoverpa armigera based on two-sex life table. Sci. Rep. 2017, 7, 1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.P.; Zhuang, J.; Huang, W.K.; Chi, H.; Wang, C.H.; Hua, H.X.; Wu, G. Different adaptability of the brown planthopper Nilaparvata lugens (Stål), to gradual and abrupt increases in atmospheric CO2. J. Pest Sci. 2020, 93, 979–991. [Google Scholar] [CrossRef]
- Sun, Y.C.; Guo, H.J.; Yuan, E.Y.; Ge, F. Elevated CO2 increases R gene-dependent resistance of Medicago truncatula against the pea aphid by up-regulating a heat shock gene. New Phytol. 2018, 217, 1697–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.J.; Wu, G.; Ge, F. Impacts of elevated CO2 on the population abundance and reproductive activity of aphid Sitobion avenae Fabricius feeding on spring wheat. J. Appl. Entomol. 2004, 128, 723–730. [Google Scholar] [CrossRef]
- Wen, D.; Liu, J.; Fan, S.; Zhang, Z.Y.; Wu, G. Evaluation on the fitness and population projection of Nilaparvata lugens in response to elevated CO2 using two-sex life table. Int. J. Pest Manag. 2019, 66, 368–377. [Google Scholar] [CrossRef]
- Whittaker, J.B. Insects and plants in a changing atmosphere. J. Ecol. 2001, 89, 507–518. [Google Scholar] [CrossRef]
- Sun, Y.C.; Ge, F. How do aphids respond to elevated CO2? J. Asia-Pac. Entomol. 2011, 14, 217–220. [Google Scholar] [CrossRef]
- Newman, J.A.; Gibsons, A.J.; Thornley, J.H.M. How predictable are aphid population responsesto elevated CO2? J. Anim. Ecol. 2003, 72, 556–566. [Google Scholar] [CrossRef]
- Liu, T.X.; Yue, B.S. Host selection, development, survival, and reproduction of turnip Aphid (Homoptera: Aphididae) on green and red cabbage varieties. J. Econ. Entomol. 2000, 93, 1308–1314. [Google Scholar]
- Qayyum, A.; Aziz, M.A.; Iftikhar, A.; Hafeez, F.; Atlihan, R. Demographic Parameters of Lipaphis erysimi (Hemiptera: Aphididae) on Different Cultivars of Brassica Vegetables. J. Econ. Entomol. 2018, 111, 1885–1894. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.S.; Zou, F.D.; Sun, Q.Z. The occurrence of alate turnip aphid Lipaphis erysimi (Homoptera: Aphididae) and its development and reproduction. J. Southwest Agric. Univ. 2002, 24, 17–20. (In Chinese) [Google Scholar]
- Chen, F.J.; Ge, F.; Su, J.W. An improved open-top chamber for research on the effects of elevated CO2 on agricultural pests in field. Chin. J. Ecol. 2005, 24, 585–590. (In Chinese) [Google Scholar]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; Nation Chung Hsing University: Taichung, Taiwan, 2017; Available online: http://140.120.197.173/Ecology/prod02.htm (accessed on 20 March 2022).
- Chi, H.; Liu, H. Two new methods for study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; Getz, W.M. Mass rearing and harvesting based on an age-stage, two-sex life table: A potato tuber worm (Lepidoptera: Gelechiidae) case study. Environ. Entomol. 1988, 17, 18–25. [Google Scholar] [CrossRef]
- Yu, J.Z.; Chi, H.; Chen, B.H. Life table and predation of Lemnia biplagiata (Coleoptera: Coccinellidae) fed on Aphis gossypii (Homoptera: Aphididae) with a proof on relationship among gross reproduction rate, net reproduction rate, and preadult survivorship. Ann. Entomol. Soc. Am. 2005, 98, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Seo, B.Y.; Kim, E.Y.; Ahn, J.J.; Kim, Y.; Kang, S.; Jung, J.K. Development, Reproduction, and Life Table Parameters of the Foxglove Aphid, Aulacorthum solani Kaltenbach (Hemiptera: Aphididae), on Soybean at Constant Temperatures. Insects 2020, 11, 296. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Q.; Wang, Q.; Rui, C.; Cui, L. The feeding behavior and life history changes in imidacloprid-resistant Aphis gossypii glover (Homoptera: Aphididae). Pest Manag. Sci. 2020, 76, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Fallahpour, F.; Ghorbani, R.; Mahallati, M.N.; Hosseini, M. Demographic parameters of Lipaphis erysimi on canola cultivars under different nitrogen fertilization regimes. J. Agric. Sci. Technol. 2015, 17, 35–47. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life History Parameters | 400 μL/L | 600 μL/L | 800 μL/L | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Gen 1 | Gen 2 | Gen 3 | Gen 1 | Gen 2 | Gen 3 | Gen 1 | Gen 2 | Gen 3 | |
| 1st instar (d) | 1.26 ± 0.05 bA | 1.32 ± 0.06 aA | 1.33 ± 0.06 aA | 1.57 ± 0.06 aA | 1.27 ± 0.06 aB | 1.38 ± 0.06 aB | 1.64 ± 0.06 aA | 1.32 ± 0.06 aB | 1.48 ± 0.06 aAB |
| 2nd instar (d) | 1.14 ± 0.04 aB | 1.05 ± 0.03 cB | 1.32 ± 0.06 abA | 1.10 ± 0.04 aA | 1.22 ± 0.05 bA | 1.20 ± 0.05 bA | 1.14 ± 0.05 aB | 1.47 ± 0.06 aA | 1.45 ± 0.07 aA |
| 3rd instar (d) | 1.10 ± 0.04 aA | 1.15 ± 0.05 aA | 1.21 ± 0.05 bA | 1.22 ± 0.05 aB | 1.25 ± 0.06 aB | 1.44 ± 0.07 aA | 1.13 ± 0.05 aB | 1.29 ± 0.06 aA | 1.17 ± 0.05 bAB |
| 4th instar (d) | 1.70 ± 0.06 aA | 1.48 ± 0.06 aB | 1.33 ± 0.06 bB | 1.34 ± 0.06 bA | 1.38 ± 0.06 aA | 1.24 ± 0.06 bA | 1.54 ± 0.07 aA | 1.15 ± 0.05 bB | 1.57 ± 0.08 aA |
| APOP (d) | 0.20 ± 0.05 aB | 0.37 ± 0.06 aA | 0.43 ± 0.06 aA | 0.14 ± 0.04 aAB | 0.05 ± 0.03 bB | 0.28 ± 0.06 aA | 0.07 ± 0.03 bB | 0.42 ± 0.06 aA | 0.34 ± 0.07 aA |
| TPOP (d) | 5.38 ± 0.07 aB | 5.37 ± 0.08 bB | 5.62 ± 0.07 abA | 5.36 ± 0.09 aA | 5.14 ± 0.06 cB | 5.57 ± 0.08 bA | 5.52 ± 0.09 aB | 5.64 ± 0.06 aAB | 5.90 ± 0.13 aA |
| Fecundity | 100.6 ± 3.5 aA | 101.4 ± 3.6 aA | 100.6 ± 3.8 aA | 102.0 ± 2.6 aA | 96.5 ± 4.4 aA | 106.2 ± 4.0 aA | 83.2 ± 5.5 bAB | 94.3 ± 4.1 aA | 79.8 ± 5.6 bB |
| Adult longevity (d) | 19.3 ± 0.8 aA | 18.0 ± 0.9 aA | 18.8 ± 0.9 aA | 17.7 ± 0.7 aA | 17.0 ± 0.9 aA | 18.7 ± 0.8 aA | 14.9 ± 1.1 bAB | 15.8 ± 0.9 aA | 13.0 ± 1.0 bB |
| Survival rate (%) | 0.92 ± 0.03 abB | 1.00 ± 0.00 aA | 0.97 ± 0.02 aAB | 0.98 ± 0.02 aA | 1.00 ± 0.00 aA | 0.90 ± 0.04 aB | 0.90 ± 0.04 bB | 0.98 ± 0.02 aA | 0.73 ± 0.06 bC |
| Generations | Population Parameters | ||||
|---|---|---|---|---|---|
| Intrinsic Rate of Increase (r) (d−1) | Finite Rate of Increase (λ) | Net Reproductive Rate (R0) (Offspring) | Mean Generation Time (T) (d) | ||
| 400 μL/L | Gen 1 | 0.4744 ± 0.0076 aA | 1.6071 ± 0.0123 aA | 92.80 ± 4.63 aA | 9.55 ± 0.11 aB |
| Gen 2 | 0.4926 ± 0.0054 aA | 1.6365 ± 0.0088 aA | 101.42 ± 3.59 aA | 9.38 ± 0.10 aB | |
| Gen 3 | 0.4552 ± 0.0052 aB | 1.5766 ± 0.0081 aB | 97.28 ± 4.37 aA | 10.06 ± 0.10 aA | |
| 600 μL/L | Gen 1 | 0.4763 ± 0.0057 aA | 1.6102 ± 0.0092 aA | 100.27 ± 3.07 aA | 9.67 ± 0.11 aB |
| Gen 2 | 0.4832 ± 0.0047 abA | 1.6213 ± 0.0076 abA | 96.52 ± 4.43 aA | 9.46 ± 0.10 aB | |
| Gen 3 | 0.4573 ± 0.0067 aB | 1.5799 ± 0.0105 aB | 95.58 ± 5.44 aA | 9.97 ± 0.08 aA | |
| 800 μL/L | Gen 1 | 0.4395 ± 0.0085 bB | 1.5519 ± 0.0131 bB | 75.06 ± 5.91 bB | 9.81 ± 0.12 aAB |
| Gen 2 | 0.4742 ± 0.0039 bA | 1.6068 ± 0.0063 bA | 92.70 ± 4.30 aA | 9.55 ± 0.09 aB | |
| Gen 3 | 0.4055 ± 0.0124 bC | 1.500 ± 0.0185 bC | 58.53 ± 6.10 bB | 10.02 ± 0.14 aA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-M.; Zhao, M.-H.; Huang, F.; Shang, F.-G.; Zhang, Y.-H.; Liu, C.-M.; He, S.-J.; Wu, G. Effects of Elevated CO2 on the Fitness of Three Successive Generations of Lipaphis erysimi. Insects 2022, 13, 333. https://doi.org/10.3390/insects13040333
Li X-M, Zhao M-H, Huang F, Shang F-G, Zhang Y-H, Liu C-M, He S-J, Wu G. Effects of Elevated CO2 on the Fitness of Three Successive Generations of Lipaphis erysimi. Insects. 2022; 13(4):333. https://doi.org/10.3390/insects13040333
Chicago/Turabian StyleLi, Xue-Mei, Mu-Hua Zhao, Feng Huang, Fang-Ge Shang, Yun-Hui Zhang, Cheng-Min Liu, Shuai-Jie He, and Gang Wu. 2022. "Effects of Elevated CO2 on the Fitness of Three Successive Generations of Lipaphis erysimi" Insects 13, no. 4: 333. https://doi.org/10.3390/insects13040333
APA StyleLi, X. -M., Zhao, M. -H., Huang, F., Shang, F. -G., Zhang, Y. -H., Liu, C. -M., He, S. -J., & Wu, G. (2022). Effects of Elevated CO2 on the Fitness of Three Successive Generations of Lipaphis erysimi. Insects, 13(4), 333. https://doi.org/10.3390/insects13040333