
Ant Mortality with Food Competition in Forests along a Temperature Gradient



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
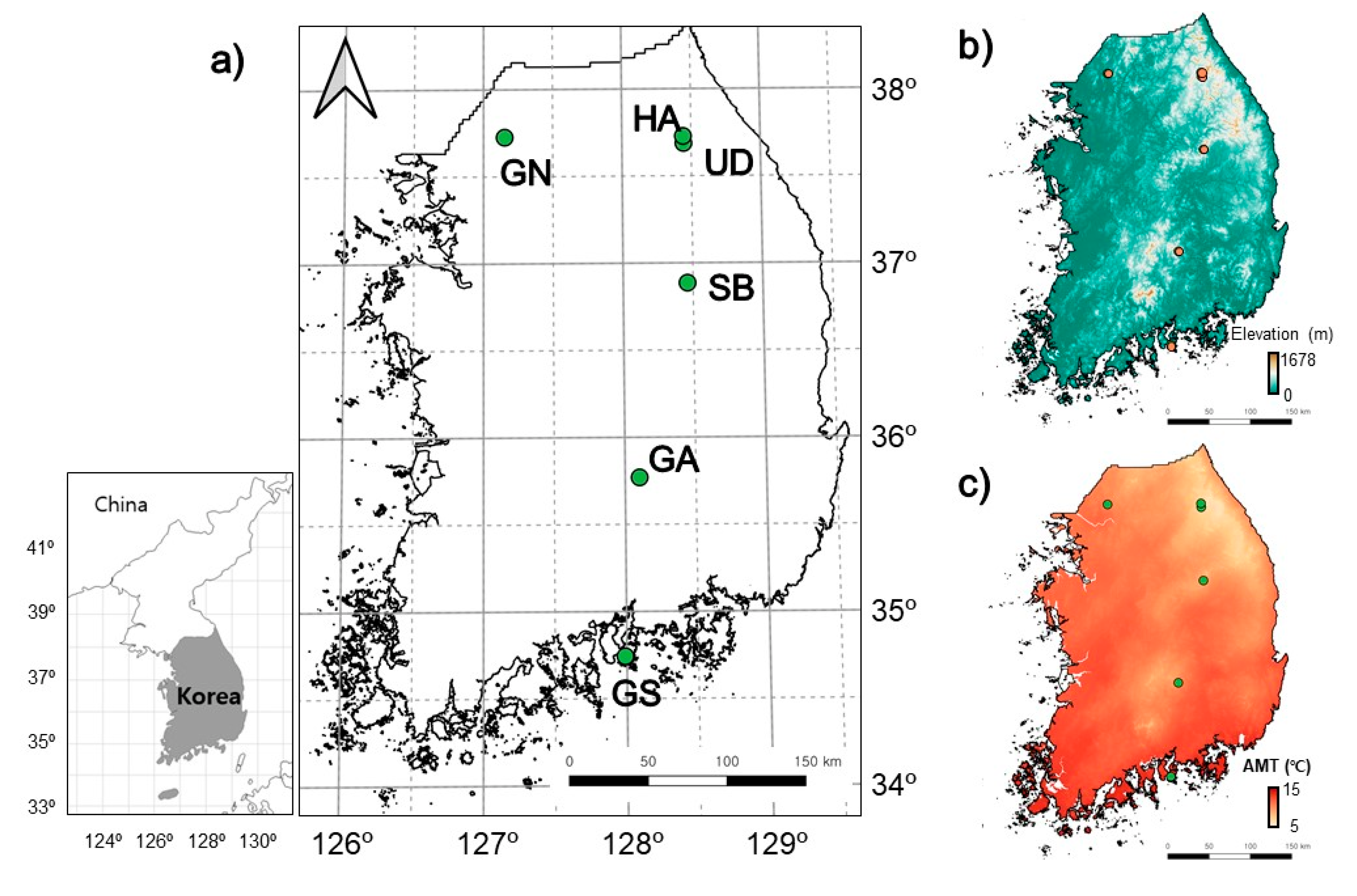
2.1. Study Sites
2.2. Field Experiment
2.3. Data Analyses
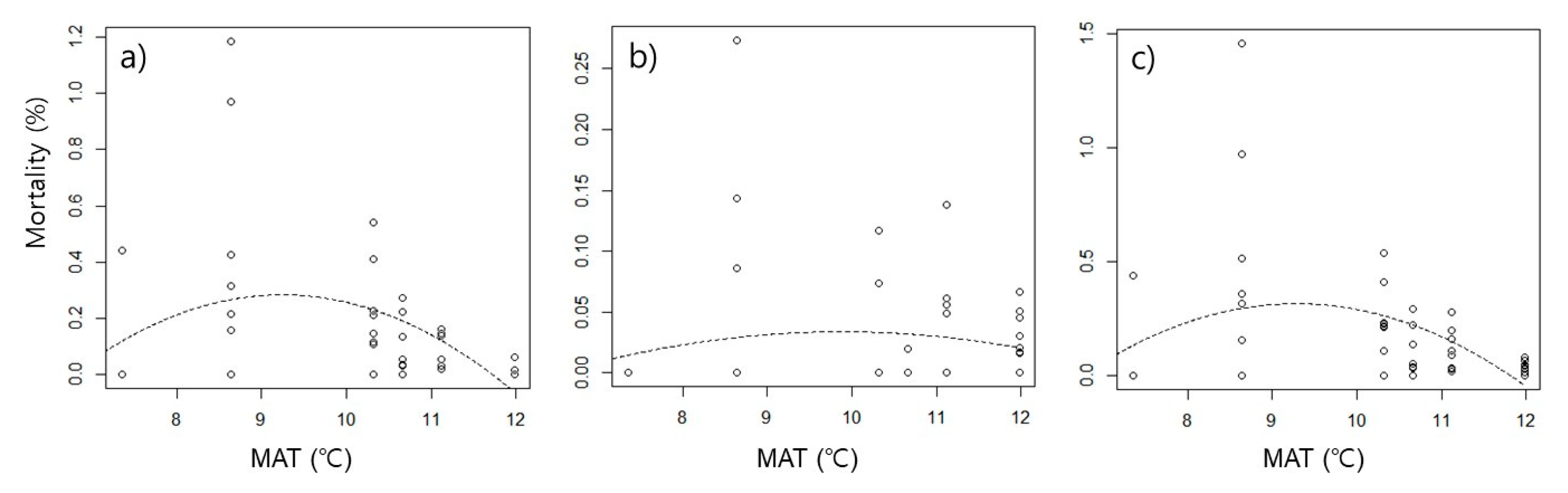
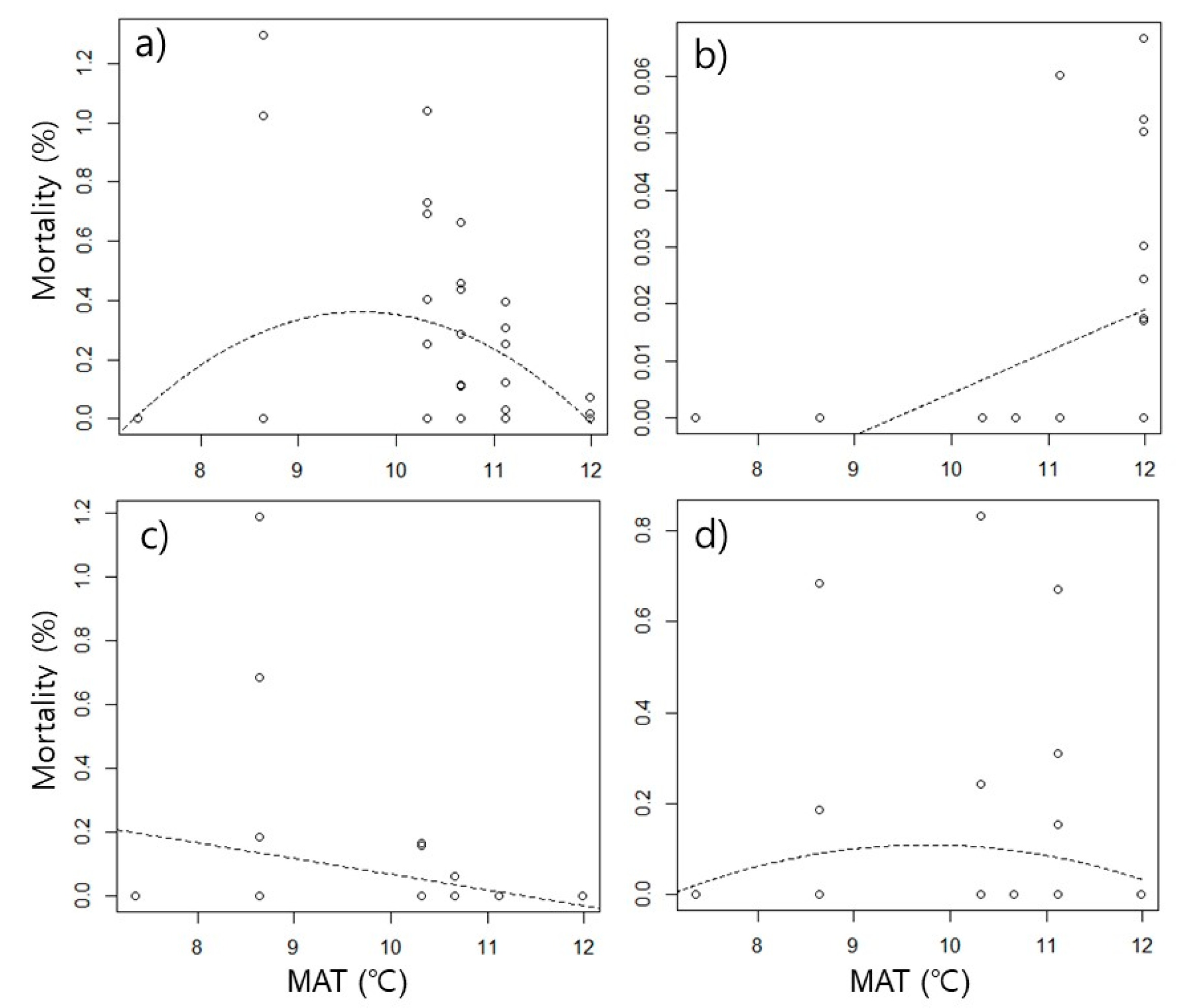
3. Results
4. Discussion
4.1. High Mortality at Intermediate Temperatures
4.2. Different Mortality among Species
4.3. Effects of Climate Change on Species Distribution
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hölldobler, B.; Wilson, E.O. The Ants; Belknap Press: Cambridge, UK, 1990. [Google Scholar]
- Suiter, D.R.; Gochnour, B.M.; Holloway, J.B.; Vail, K.M. Alternative methods of ant (Hymenoptera: Formicidae) control with emphasis on the Argentine ant, Linepithema humile. Insects 2021, 12, 487. [Google Scholar] [CrossRef] [PubMed]
- Dieng, H.; Zawawi, R.B.M.; Yusof, N.I.S.B.M.; Ahmad, A.H.; Abang, F.; Ghani, I.A.; Satho, T.; Ahmad, H.; Zuharah, W.F.; Majid, A.H.A.; et al. Green tea and its waste attract workers of formicine ants and kill their workers—implications for pest management. Ind. Crops Prod. 2016, 89, 157–166. [Google Scholar] [CrossRef]
- Carroll, C.R.; Janzen, D.H. Ecology of foraging by ants. Annu. Rev. Ecol. Syst. 1973, 4, 231–257. [Google Scholar] [CrossRef]
- Cerdá, X.; Arnan, X.; Retana, J. Is competition a significant hallmark of ant (Hymenoptera: Formicidae) ecology? Myrmecol. News 2013, 18, 131–147. [Google Scholar]
- Feener, D.H., Jr.; Orr, M.R.; Wackford, K.M.; Longo, J.M.; Benson, W.W.; Gilbert, L.E. Geographic variation in resource dominance-discovery in Brazilian ant communities. Ecology 2008, 89, 1824–1836. [Google Scholar] [CrossRef] [Green Version]
- Tizón, R.; Wulff, J.P.; Peláez, D.V. The effect of increase in the temperature on the foraging of Acromyrmex lobicornis (Hymenoptera: Formicidae). Zool. Stud. 2014, 53, 40. [Google Scholar] [CrossRef] [Green Version]
- Seifert, B. The ecology of Central European non-arboreal ants—37 years of a broad-spectrum analysis under permanent taxonomic control. Soil Org. 2017, 89, 1–67. [Google Scholar]
- Frizzi, F.; Bartalesi, V.; Santini, G. Combined effects of temperature and interspecific competition on the mortality of the invasive garden ant, Lasius neglectus: A laboratory study. J. Therm. Biol. 2017, 65, 76–81. [Google Scholar] [CrossRef]
- Cerdá, X.; Retana, J.; Manzaneda, A. The role of competition by dominants and temperature in the foraging of subordinate species in Mediterranean ant communities. Oecologia 1998, 117, 404–412. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.S. High competition between ant species at intermediate temperatures. J. Therm. Biol. 2018, 72, 59–66. [Google Scholar] [CrossRef]
- Rosumek, F.B. Natural history of ants: What we (do not) know about trophic and temporal niches of neotropical species. Sociobiology 2017, 64, 244–255. [Google Scholar] [CrossRef] [Green Version]
- Rosumek, F.B.; Blüthgen, N.; Brückner, A.; Menzel, F.; Gebauer, G.; Heethoff, M. Unveiling community patterns and trophic niches of tropical and temperate ants using an integrative framework of field data, stable isotopes and fatty acids. PeerJ 2018, 6, e5467. [Google Scholar] [CrossRef]
- Briese, D.; Macauley, B. Temporal structure of an ant community in semi-arid Australia. Aust. J. Ecol. 1980, 5, 121–134. [Google Scholar] [CrossRef]
- Cros, S.; Cerdá, X.; Retana, J. Spatial and temporal variations in the activity patterns of Mediterranean ant communities. Ecoscience 1997, 4, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Bestelmeyer, B.T. The trade-off between thermal tolerance and behavioural dominance in a subtropical South American ant community. J. Anim. Ecol. 2000, 69, 998–1009. [Google Scholar] [CrossRef]
- McGlynn, T.P.; Eben Kirksey, S. The effects of food presentation and microhabitat upon resource monopoly in a ground-foraging ant (Hymenoptera: Formicidae) community. Rev. Biol. Trop. 2000, 48, 629–642. [Google Scholar]
- LeBrun, E.G. Who is the top dog in ant communities? Resources, parasitoids, and multiple competitive hierarchies. Oecologia 2005, 142, 643–652. [Google Scholar] [CrossRef]
- Passera, L.; Roncin, E.; Kaufmann, B.; Keller, L. Increased soldier production in ant colonies exposed to intraspecific competition. Nature 1996, 379, 630–631. [Google Scholar] [CrossRef]
- Thomas, M.L.; Tsutsui, N.D.; Holway, D.A. Intraspecific competition influences the symmetry and intensity of aggression in the Argentine ant. Behav. Ecol. 2005, 16, 472–481. [Google Scholar] [CrossRef]
- Kwon, T.-S.; Kim, S.-S.; Lee, C.; Jung, S.; Sung, J. Korean Ant Atlas (2006–2009); Korea Forest Research Institute: Seoul, Korea, 2012. [Google Scholar]
- Roback, P.; Legler, J. Beyond Multiple Linear Regression: Applied Generalized Linear Models and Multilevel Models in R; Chapman and Hall/CRC: Boca Raton, FL, USA, 2020. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 23 June 2018).
- Parr, C.L.; Gibb, H. Competition and the role of dominant ants. In Ant Ecology; Lach, L., Parr, C., Abbott, K.I., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 77–96. [Google Scholar]
- Retana, J.; Cerdá, X. Agonistic relationships among sympatric mediterranean ant species (Hymenoptera: Formicidae). J. Insect Behav. 1994, 8, 365–380. [Google Scholar] [CrossRef]
- Cerdá, X.; Retana, J.; Cros, S. Prey size reverses the outcome of interference interactions of scavenger ants. Oikos 1998, 82, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Nonacs, P.; Dill, L.M. Mortality risk vs. food quality trade-offs in a common currency: Ant patch preferences. Ecology 1990, 71, 1886–1892. [Google Scholar] [CrossRef]
- Kwon, T.-S.; Kim, S.-S.; Chun, J.H. Pattern of ant diversity in Korea: An empirical test of Rapoport’s altitudinal rule. J. Asia-Pac. Entomol. 2014, 17, 161–167. [Google Scholar] [CrossRef]
- Kwon, T.-S.; Lee, C.M. Prediction of abundance of ants according to climate change scenarios RCP 4.5 and 8.5 in South Korea. J. Asia-Pac. Biodivers. 2015, 8, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Fellers, J.H. Interference and exploitation in a guild of woodland ants. Ecology 1987, 68, 1466–1478. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | UD (7.35 °C) | HA (8.64 °C) | GN (10.32 °C) | SB (10.60 °C) | GA (11.12 °C) | GS (11.98 °C) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Abund | Occur | Abund | Occur | Abund | Occur | Abund | Occur | Abund | Occur | Abund | Occur | |
| Aphaenogaster japonica | 9 | 33.75 | 1692 | 40.63 | 5430 | 63.75 | 9153 | 79.38 | 6248 | 79.38 | 2 | 0.63 |
| Camponotus atrox | 242 | 59.38 | 246 | 48.75 | 26 | 8.75 | 0 | 0 | 0 | 0 | 0 | 0 |
| Camponotus nipponensis | 0 | 0 | 0 | 0 | 21 | 1.25 | 1 | 0.63 | 0 | 0 | 0 | 0 |
| Camponotus sp. | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.63 | 0 | 0 |
| Crematogaster osakensis | 0 | 0 | 0 | 0 | 0 | 0 | 604 | 1.25 | 0 | 0 | 0 | 0 |
| Crematogaster vagula | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 1.25 | 597 | 3.75 |
| Ectomomyrmex javanus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 3.75 | 117 | 21.25 |
| Formica japonica | 2 | 1.25 | 0 | 0 | 254 | 21.88 | 22 | 5 | 334 | 20.63 | 0 | 0 |
| Lasius japonicus | 28 | 6.88 | 1 | 0.63 | 1460 | 21.88 | 618 | 15.63 | 466 | 1.88 | 138 | 3.75 |
| Lasius spathepus | 214 | 0.63 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Myrmica spp. | 536 | 83.13 | 2460 | 78.75 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Nylanderia flavipes | 36 | 0.63 | 1074 | 25.00 | 2655 | 68.13 | 6927 | 95.63 | 12,882 | 94.38 | 42,289 | 100.00 |
| Pheidole fervida | 229 | 11.25 | 84 | 2.50 | 15,201 | 44.38 | 12,077 | 38.75 | 8484 | 23.13 | 946 | 5.63 |
| Themnothorax nassonovi | 199 | 35.00 | 0 | 0 | 2 | 0.63 | 0 | 0 | 0 | 0 | 0 | 0 |
| Vollenhovia emeryi | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0.63 | 0 | 0 | 1 | 0.63 |
| MAT (°C) | Killer Species | Victim Species | ||||
|---|---|---|---|---|---|---|
| Lasius japonicus | Pheidole fervida | Myrmica spp. | Nylanderia flavipes | Aphaenogaster japonica | ||
| 7.35 | Camponotus atrox | 1 | ||||
| 8.64 | A. japonica | 2 | 3 | 1 | 3 | |
| C. atrox | 9 | 6 | 4 | |||
| Myrmica spp. | 3 | |||||
| 10.32 | A. japonica | 34 | 9 | 3 | ||
| C. atrox | 6 | 1 | ||||
| F. japonica | 1 | |||||
| P. fervida | 1 | 1 | ||||
| 10.60 | A. japonica | 1 | 12 | 16 | ||
| F. japonica | 1 | |||||
| L. japonicus | 2 | |||||
| P. fervida | 1 | 1 | ||||
| 11.12 | A. japonica | 3 | 12 | 6 | ||
| F. japonica | 1 | |||||
| N. flavipes | 2 | |||||
| P. fervida | 2 | 5 | ||||
| 11.98 | L. japonicus | 2 | ||||
| N. flavipes | 17 | |||||
| Ectomomyrmex javanus | 1 | |||||
| SUM | 1 | 61 | 16 | 76 | 19 | |
| Species | Weight (mg) | N | Dominance |
|---|---|---|---|
| Camponotus atrox | 7.942 (3.594) | 22 | 87.2 |
| Ectomomyrmex javanus | 5.761 (0.694) | 20 | |
| Formica japonica | 1.770 (0.642) | 20 | 30.2 |
| Myrmica kotokui | 1.048 (0.124) | 19 | 30.9 |
| Myrmica kurokii | 0.747 (0.172) | 22 | |
| Aphaenogaster japonica | 0.679 (0.108) | 19 | 84 |
| Pheidole fervida (major) | 0.375 (0.063) | 30 | 79.7 |
| Pheidole fervida (minor) | 0.107 (0.018) | 25 | |
| Lasius japonicus | 0.211 (0.075) | 24 | 50.3 |
| Crematogaster vagula | 0.185 (0.045) | 2 | |
| Crematogaster matsumurai | 0.165 (0.035) | 27 | |
| Vollenhovia emeryi | 0.108 (0.018) | 5 | |
| Themnothorax nassonovi | 0.085 (0.026) | 40 | |
| Nylanderia flavipes | 0.068 (0.017) | 26 | 3.6 |
| Species | Intraspecific Competition | Interspecific Competition | Total |
|---|---|---|---|
| Aphaenogaster japonica | 12 (0.05) | 7 (0.03) | 19 (0.08) |
| Lasius japonicus | 1 (0.04) | 1 (0.04) | |
| Myrmica spp. | 3 (0.10) | 13 (0.43) | 16 (0.53) |
| Nylanderia flavipes | 19 (0.03) | 57 (0.09) | 76 (0.12) |
| Pheidole fervida | 3 (0.01) | 58 (0.16) | 61 (0.16) |
| Killer Species | Victim Species | ||||
|---|---|---|---|---|---|
| Lasius japonicus | Pheidole fervida | Myrmica spp. | Nylanderia flavipes | Aphaenogaster japonica | |
| L. japonicus | 2 | ||||
| P. fervida | 2 | 4 | |||
| Myrmica spp. | 2 | ||||
| N. flavipes | 11 | ||||
| A. japonica | 1 | 2 | 2 | 5 | |
| Victim Species | Killer | Variable | Full Model | Minimum Adequate Model | ||
|---|---|---|---|---|---|---|
| Coef. | AIC | Coef. | AIC | |||
| Nylanderia flavipes | Other species | AT | 0.13 * | 126.4 | 0.13 * | 126.4 |
| MAT | 9.977 *** | 9.977 *** | ||||
| MAT2 | −0.548 *** | −0.548 *** | ||||
| Same species | AT | −0.04 ns | 36.6 | 33.5 | ||
| MAT | 0.063 ns | 2.213 * | ||||
| MAT2 | −0.274 ns | |||||
| Total | AT | 0.067 ns | 136.1 | 137.2 | ||
| MAT | 4.44 * | 4.879 * | ||||
| MAT2 | −0.257 ** | −0.275 ** | ||||
| Pheidole fervida | Other species | AT | 0.2 *** | 87 | 0.2 *** | 87 |
| MAT | 14.758 ** | 14.758 ** | ||||
| MAT2 | −0.838 *** | −0.838 *** | ||||
| Same species | AT | 0.114 ns | 24 | |||
| MAT | 0.001 ns | |||||
| MAT2 | −0.633 ns | |||||
| Total | AT | 0.195 *** | 24 | 0.195 *** | 24 | |
| MAT | 11.463 ** | 11.463 ** | ||||
| MAT2 | −0.654 ** | −0.654 ** | ||||
| Aphaenogaster japonica | Other species | AT | 0.247 ns | 35.4 | 34.5 | |
| MAT | 21.265 ns | −0.075 *** | ||||
| MAT2 | −1.21 ns | |||||
| Same species | AT | 0.267 * | 34.5 | 0.267 * | 34.5 | |
| MAT | −13.532 * | −13.532 * | ||||
| MAT2 | 0.65 * | 0.65 * | ||||
| Total | AT | 0.249 * | 78 | 0.245 * | 79.5 | |
| MAT | −7.922 ns | −1.343 *** | ||||
| MAT2 | 0.342 ns | |||||
| Total | Other species | AT | 0.182 *** | NE | 0.182 *** | NE |
| MAT | 7.732 *** | 7.732 *** | ||||
| MAT2 | −0.444 *** | −0.444 *** | ||||
| Same species | AT | 0.007 ns | 110.4 | 108.4 | ||
| MAT | −4.457 * | −4.466 * | ||||
| MAT2 | 0.217 * | 0.218 * | ||||
| Total | AT | 0.128 ** | NE | 0.128 ** | NE | |
| MAT | 3.337 * | 3.337 * | ||||
| MAT2 | −0.205 * | −0.205 * | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, T.-S.; Lee, D.-S.; Park, Y.-S. Ant Mortality with Food Competition in Forests along a Temperature Gradient. Insects 2022, 13, 392. https://doi.org/10.3390/insects13040392
Kwon T-S, Lee D-S, Park Y-S. Ant Mortality with Food Competition in Forests along a Temperature Gradient. Insects. 2022; 13(4):392. https://doi.org/10.3390/insects13040392
Chicago/Turabian StyleKwon, Tae-Sung, Dae-Seong Lee, and Young-Seuk Park. 2022. "Ant Mortality with Food Competition in Forests along a Temperature Gradient" Insects 13, no. 4: 392. https://doi.org/10.3390/insects13040392
APA StyleKwon, T. -S., Lee, D. -S., & Park, Y. -S. (2022). Ant Mortality with Food Competition in Forests along a Temperature Gradient. Insects, 13(4), 392. https://doi.org/10.3390/insects13040392