
Temperature Dependence for Survival, Development, and Reproduction of the Cactus Cochineal Dactylopius opuntiae (Cockerell)

Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Establishment of D. opuntiae Colony
2.2. Biological Parameters
2.3. Reproduction Parameters
2.4. The Cochineal Thermal Requirements
2.5. Life Table Parameters
3. Results
3.1. Biological Parameters
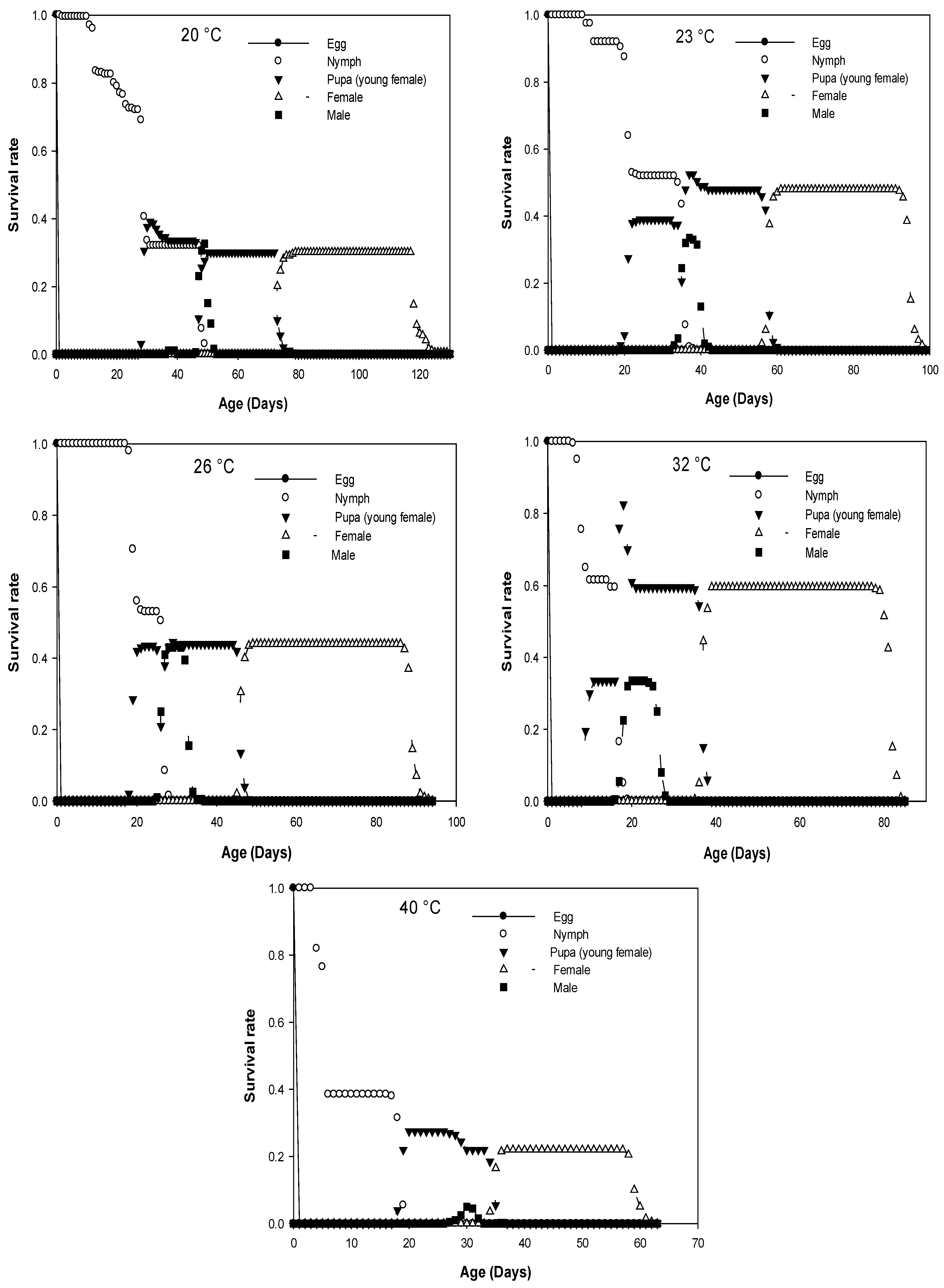
3.2. Survival Rate and Sex Ratio
3.3. Reproduction Parameters
3.4. Population Growth Parameters
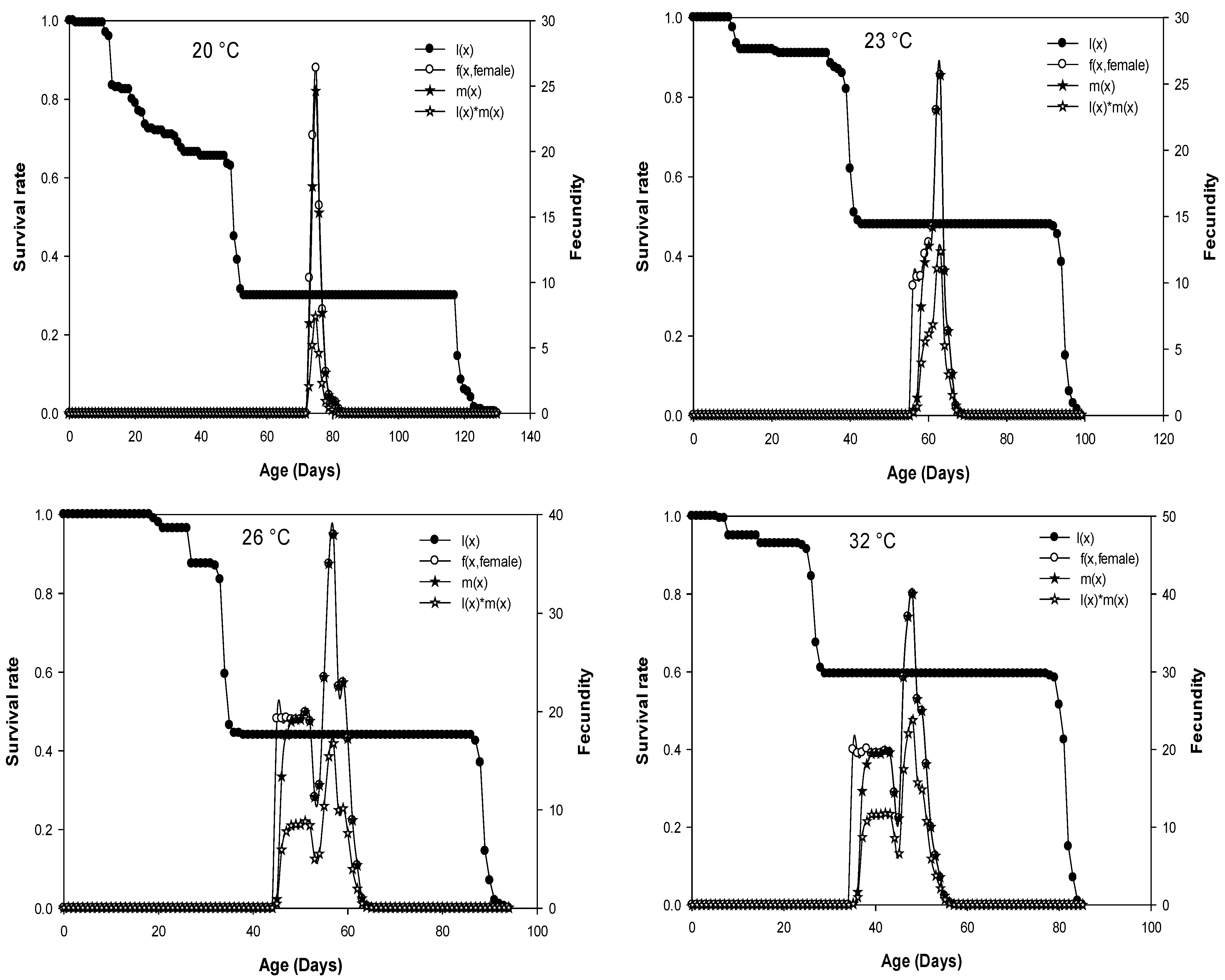
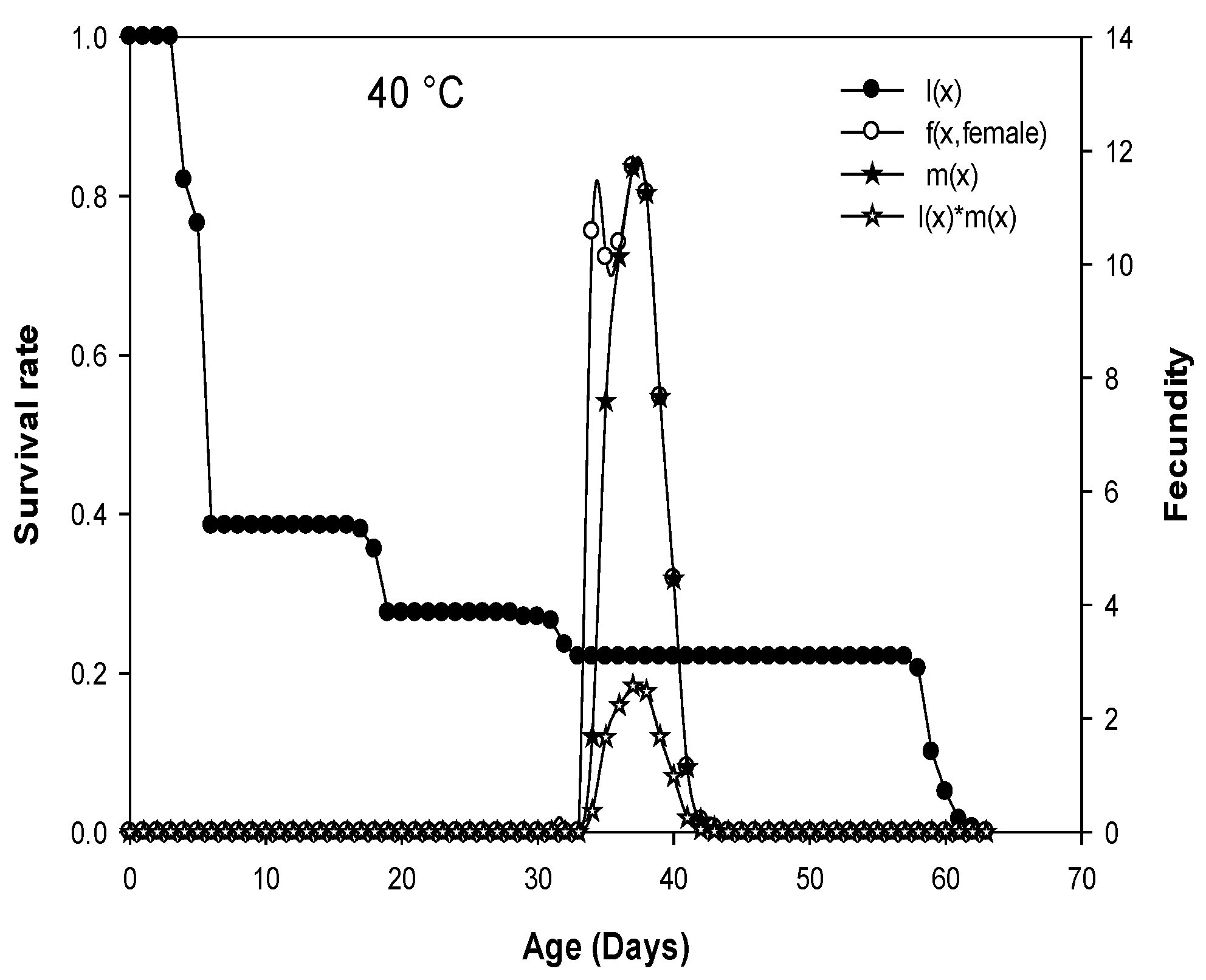
3.5. Life Table Parameters
3.6. Thermal Requirements
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paterson, I.D.; Hoffmann, J.H.; Klein, H.; Mathenge, C.W.; Neser, S.; Zimmermann, H.G. Biological control of Cactaceae in South Africa. Afr. Entomol. 2011, 19, 230–246. [Google Scholar] [CrossRef]
- Klein, H. Biological control of invasive cactus species (Family Cactaceae). 2.2. Cochineal insects (Dactylopius spp.). PPRI Leaflet Ser. Weeds Biocontrol. 2002, 2, 1–3. [Google Scholar]
- Spodek, M.; Ben-Dov, Y.; Protasov, A.; Carvalho, C.J.; Mendel, Z. First record of Dactylopius opuntiae (Cockerell) (Hemiptera: Coccoidea: Dactylopiidae) from Israel. Phytoparasitica 2014, 42, 377–379. [Google Scholar] [CrossRef]
- Bouharroud, R.; Amarraque, A.; Qessaoui, R. First report of the Opuntia cochineal scale Dactylopius opuntiae (Hemiptera: Dactylopiidae) in Morocco. EPPO Bull. 2016, 46, 308–310. [Google Scholar] [CrossRef]
- Mazzeo, G.; Nucifora, S.; Russo, A.; Suma, P. Dactylopius opuntiae, a new prickly pear cactus pest in the Mediterranean: An overview. Entomol. Exp. Appl. 2019, 167, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Iannetta, M. Interazioni tra cambiamenti climatici e desertificazione. Memorie di Scienze Fisiche e Naturali 125° 2007, 31, 277–295. [Google Scholar]
- Stange, E.E.; Ayres, M.P. Climate Change Impacts: Insects. Available online: http://www.els.net (accessed on 3 January 2019).
- MacGregor, L.R.; Sampedro, R.G. Catălogo de coccidos Mexicanos—Familia Dactylopiidae (Homoptera-Coccoidea). An. Inst. Cienc. Mar Limnol. Univ. Nac. Auton. Mex 1984, 54, 217–223. [Google Scholar]
- Portillo, M.; Vigueras, A. A Review on the cochineal species in Mexico, hosts and natural enemies. Acta Hortic. 2006, 728, 249–256. [Google Scholar] [CrossRef]
- Badii, M.H.; Flores, A.E. Prickly pear cacti pests and their control inMexico. Fla. Entomol. 2001, 84, 503–505. [Google Scholar] [CrossRef]
- Khan, H.A.A.; Sayyed, A.H.; Akram, W.; Raza, S.; Ali, M. The predatory potential of Chrysoperla carnea and Cryptolaemus montrouzieri larvae on different stages of the mealybug, Phenacoccus solenopsis: A threat to cotton in South Asia. J. Insect Sci. 2012, 12, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Aalaoui, M.; Bouharroud, R.; Sbaghi, M.; El Bouhssini, M.; Hilali, L. Seasonal biology of Dactylopius opuntiae (Hemiptera: Dactylopiidae) on Opuntia ficus-indica (Caryophyllales: Cactaceae) under field and semi-field conditions in Morocco. Ponte 2020, 1, 259–271. [Google Scholar] [CrossRef]
- Chávez-Moreno, C.K.; Tecante, A.; Casas, A.; Claps, L.E. Distribution and habitat in Mexico of Dactylopius costa Dactylopius opuntiae, alien pest in theMediterranean 67 (Hemiptera: Dactylopiidae) and their cacti hosts (Cactaceae: Opuntioideae). Neotrop. Entomol. 2011, 40, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Mann, J. Cactus-feeding insects and mites. Bull. US Natl. Mus. 1969, 256, 1–158. [Google Scholar] [CrossRef]
- Vanegas-Rico, J.M.; Rodríguez-Leyva, E.; Lomeli-Flores, J.R.; González-Hernández, H.; Pérez-Panduro, A.; MoraAguilera, G. Biology and life history of Hyperaspis trifurcate feeding on Dactylopius opuntiae. Bio. Control. 2016, 61, 691–701. [Google Scholar]
- Batista-Lopes, E.; De Brito, C.H.; Cavalcanti, I.; De Luna, B.J. Selecáo de genotipos de palma forrageira (Opuntia spp.) e (Nopalea spp.) resistentes a cochonilha-do-carmim (Dactylopius opuntiae Cockerell, 1929) na Paraíba. Brasil. Eng. Ambient. Pesquisa Tecnologia. 2010, 7, 204–215. (In Portuguese) [Google Scholar]
- Santos, D.C.; Lira, M.A.; Farias, I.; Dias, F.M.; Costa, A.F.; Pereira, V.L.A.; Silva, D.D. Selecăo de Clones de Palma Forrageira Resistentes ă Cochonilha do Carmim Dactylopius sp. em Condicŏes de Campo; Reuniăo Anual da Sociedade Brasileira de Zootecnia: Jŏao Pessoa, Paraȋba, Brazil, 2006. [Google Scholar]
- Lopes, E.B.; Batista, J.L.; Brito, C.H.; Santos, D.C. Pragas da palma. Palma Forrageira: Cultivo, Uso Atual e Perspectivas de Utilizacăo no Semiarido Nordestino; Lopes, E.B., Ed.; EMEPA/FAEPA: Joăo Pessoa, Paraíba, Brazil, 2007; pp. 34–40. [Google Scholar]
- Cavalcanti, V.A.L.B.; Sena, R.C.; Coutinho, J.L.B.; Arruda, G.P.; Rodrigues, F.B. Controle das cochonilhas da palma forrageira. BoletimIPA Responde 2001, 39, 1–2. [Google Scholar]
- Amarasekare, K.; Chong, J.H.; Epsky, N.D.; Mannion, C.M. Effect of temperature on the life history of the mealybug Paracoccus marginatus (Hemiptera: Pseudococcidae). J. Econ. Entomol. 2008, 101, 1798–1804. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.H.; Roda, A.L.; Mannion, C.M. Life history of the mealybug, Maconellicoccus hirsutus (Hemiptera: Psuedococcidae) at constant temperatures. Environ. Entomol. 2008, 37, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.C.; Song, J.H.; Kim, D.S. Effect of temperature on the development and fecundity of the cryptic mealybug Pseudococcus cryptus in the laboratory. J. Asia Pac. Entomol. 2008, 11, 149–153. [Google Scholar] [CrossRef]
- Huffaker, C.B.; Berryman, A.; Turchin, P. Dynamics and regulation of insect populations. Ecol. Entomol. 1999, 2, 269–305. [Google Scholar]
- Huang, Z.; Ren, S.; Musa, P.D. Effects of temperature on development, survival, longevity, and fecundity of the Bemisia tabaci Gennadius (Homoptera: Aleyrodidae) predator, Axinoscymnus cardilobus (Coleoptera: Coccinellidae). Biol. Control. 2008, 46, 209–215. [Google Scholar] [CrossRef]
- Prasad, Y.G.; Prabhakar, M.; Sreedevi, G.; Rao, G.R.; Venkateswarlu, B. Effect of temperature on development, survival and reproduction of the mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) on cotton. Crop. Prot. 2012, 39, 81–88. [Google Scholar] [CrossRef]
- Aysal, T.; Kivan, M. Development and population growth of Stephanitis pyri (F.) (Heteroptera: Tingidae) at five temperatures. J. Pest Sci. 2008, 81, 135–141. [Google Scholar] [CrossRef]
- Mota, J.A.; Soares, A.O.; Garcia, P.V. Temperature dependence for development of the whitefly predator Clitostethus arcuatus (Rossi). BioControl 2008, 53, 603–613. [Google Scholar] [CrossRef]
- Briere, J.F.; Pracros, P.; Le Roux, A.Y.; Pierre, J.S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Honěk, A.; Kocourek, F. Temperature and development time in insects: A general relationship between thermal constants. Zool. Syst. 1990, 117, 401–439. [Google Scholar]
- Sullivan, P.R. Population growth potential of Dactylopius ceylonicus Green (Hemiptera: Dactylopiidae) on Opuntia vulgaris Miller. J. Aust. Entomol. Soc. 1990, 29, 123–129. [Google Scholar] [CrossRef]
- Hosking, J.R. The effect of temperature on the population growth potential of Dactylopius austrinus De Lotto (Homoptera: Dactylopiidae), on Opuntia aurantiaca Lindley. J. Aust. entomol. Soc. 1984, 23, 133–139. [Google Scholar] [CrossRef]
- Aldama-Aguilera, C.; Llanderal-Cázares, C. Cochineal: Comparison of production methods in cut cladodes. Agrociencia 2003, 37, 11–19. [Google Scholar]
- Vinogradova, E.B.; Reznik, S.Y. Induction of larval diapause in the blowfly, Calliphora vicina R.-D.(Diptera, Calliphoridae) under field and laboratory conditions. Entomol. Rev. 2013, 93, 935–941. [Google Scholar] [CrossRef]
- Flores-Hernández, A.; Murillo-Amador, B.; Rueda-Puente, E.O.; Salazar-Torres, J.C.; García-Hernández, J.L.; Troyo Diéguez, E. Reproduction of wild cochineal Dactylopius opuntiae (Homoptera: Dactylopiidae). Rev. Mex. Biodivers. 2006, 77, 97–102. [Google Scholar]
- El Aalaoui, M.; Bouharroud, R.; Sbaghi, M.; El Bouhssini, M.; Hilali, L.; Dari, K. Comparative toxicity of different chemical and biological insecticides against the scale insect Dactylopius opuntiae and their side effects on the predator Cryptolaemus montrouzieri. Arch. Phytopathol. Plant Prot. 2019, 52, 155–169. [Google Scholar] [CrossRef]
- Abbas, N.; Shad, S.A.; Razaq, M. Fitness cost, cross resistance and realized heritability of resistance to imidacloprid in Spodoptera litura (Lepidoptera: Noctuidae). Pestic. Biochem. Phys. 2012, 103, 181–188. [Google Scholar] [CrossRef]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Gutierrez, A.P.; Mackauer, M. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar] [CrossRef]
- Honěk, A.; Kocourek, F. Thermal requirements for development of aphidophagous Coccinellidae (Coleoptera), Chrysopidae, Hemerobiidae (Neuroptera), and Syrphidae (Diptera): Some general trends. Oecologia 1988, 76, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.F. Predictive population equation based on key factors. Memoirs Entomol. Soc. Can. 1963, 32, 16–21. [Google Scholar] [CrossRef]
- Ju, R.T.; Wang, F.; Li, B. Effects of temperature on the development and population growth of the sycamore lace bug. Corythucha ciliata. J. Insect Sci. 2011, 11, 16. Available online: http://www.insectscience.org/11.16/i1536-2442-11-16.pdf (accessed on 1 January 2011).
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Programfor the Age-Stage, Two-Sex Life Table Analysis. 2017. Available online: http://140.120.197.173/Ecology/ (accessed on 5 May 2017).
- Carver, R.H.; Nash, J.G. Doing Data Analysis with SPSS: Version 18.0; Cengage Learning: Boston, MA, USA, 2011. [Google Scholar]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Moran, V.C.; Cobby, B.S. On the life history and fecundity of the cochineal insect, Dactylopius austrinus De Lotto (Homoptera: Dactylopiidae), a biocontrol agent for the cactus Opuntia aurantiaca. Bull. Entomol. Res. 1979, 69, 629–636. [Google Scholar] [CrossRef]
- Mathenge, C.W.; Holford, P.; Hoffmann, J.H.; Spooner-Hart, R.; Beattie, G.A.C.; Zimmermann, G. The biology of Dactylopius tomentosus (Hemiptera: Dactylopiidae). Bull. Entomol. Res. 2009, 99, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Guerra, G.P.; Kosztarab, M. Biosystematics of the Family Dactylopiidae (Homoptera: Coccineae) with Emphasis on the Life Cycle of Dactylopius coccus Costa: Studies on the Morphology and Systematics of Scale Insects No. 16; Bulletin No. 92–1; Virginia Agricultural Experiment Station, Virginia Polytechnic Institute and State University: Blacksburg, VA, USA, 1992. [Google Scholar]
- Dixon, A.F.G. Insect Predator-Prey Dynamics. Ladybird Beetles and Biological Control; Cambridge University Press: Cambridge, MA, USA, 2000; p. 257. [Google Scholar]
- Hodek, I.; Honěk, A. Ecology of Coccinellidae; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; p. 464. [Google Scholar]
- Waterworth, R.A.; Wright, I.M.; Millar, J.G. Reproductive biology of three cosmopolitan mealybug (Hemiptera: Psuedococcidae) species, Pseudococcus longispinus, Pseudococcus viburni and Planococcus ficus. Ann. Entomol. Soc. Am. 2011, 104, 249–260. [Google Scholar] [CrossRef]
- Luna, J.A.P.; Leyva, E.R.; Lomeli-Flores, J.R.; Guzmán, A.L.V.; Rico, J.M.V. Ciclo de vida y fecundidad de “Dactylopius opuntiae” (hemiptera: Dactylopiidae) en “Opuntia ficus-indica” (caryophyllales: Cactaceae). Agrociencia 2018, 52, 103–114. [Google Scholar]
- Yu, J.Z.; Chen, B.H.; Güncan, A.; Atlihan, R.; Gökçe, A.; Smith, C.L.; Gümüs, E.; Chi, H. Demography and mass-rearing Harmonia dimidiate (Coleoptera: Coccinellidae), using Aphis gossypii (Hemiptera: Aphididae) and eggs of Bactrocera dorsalis (Diptera: Tephritidae). J. Econ. Entomol. 2018, 111, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Seyfollahi, F.; Esfandiari, M.; Mossadegh, M.S.; Rasekh, A. Life table parameters of the coccinellid Hyperaspis polita, a native predator in Iran, feeding on the invasive mealybug Phenacoccus solenopsis. J. Asia-Pac. Entomol. 2016, 19, 835–840. [Google Scholar] [CrossRef]
- Cruz-Rodríguez, J.A.; Gonzălez-Machorro, E.; Villegas González, A.A.; Rodríguez Ramírez, M.L.; Mejía Lara, F. Autonomous biological control of Dactylopius opuntiae (Hemiptera: Dactyliiopidae) in a prickly pear plantation with ecological management. Environ. Entomol. 2016, 45, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Watson, G.W.; Zhang, R. The potential distribution of an invasive mealybug Phenacoccus solenopsis and its threat to cotton in Asia. Agric. For. Entomol. 2010, 12, 403–416. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Duration | Temperature ( °C) | |||||
|---|---|---|---|---|---|---|
| 20 | 23 | 26 | 32 | 40 | ||
| Egg incubation period (hours) | 23.82 ±0.46 A | 21.63 ±0.92 B | 17.39 ±0.94 C | 7.81 ±0.49 D | 2.31 ±0.47 E | |
| First-instar nymph (days) | Female | 12.14 ±0.43 A * b ** | 11.83 ±0.62 Ba | 11.03 ±0.45 Ca | 2.17 ±0.42 Eb | 4.82 ±0.39 Da |
| Male | 12.33 ±0.65 Aa | 11.91 ±0.68 Ba | 11.15 ±0.53 Ca | 2.36 ±0.57 Ea | 5.05 ±0.69 Da | |
| Second-instar nymph (days) | Female | 35.20 ±0.51 Aa | 23.13 ±0.37 Ba | 15.12 ±0.36 Ca | 14.20 ±0.50 Da | 13.14 ±0.35 Ea |
| Male | 16.03 ±0.35 Ab | 8.14 ±0.35 Cb | 7.20 ±0.43 Db | 6.18 ±0.42 Eb | 13.36 ±0.50 Ba | |
| Third-instar nymph (days) | Young female | 25.34 ±0.95 Aa | 22.11 ±0.35 Ba | 19.19 ±0.48 Da | 19.89 ±0.43 Ca | 16.16 ±0.37 Ea |
| Male Pupal duration (Pupa + Cocoon) | 18.21 ±0.56 Ab | 14.15 ±0.36 Bb | 7.14 ±0.41 Db | 9.67 ±0.56 Cb | 9.73 ±0.47 Cb | |
| Total pre-adult (days) | Female | 73.67 ±1.23 Aa | 58.13 ±0.85 Ba | 46.36 ±0.75 Ca | 37.26 ±0.78 Da | 35.11 ±0.69 Ea |
| Male | 47.10 ±1.90 Ab | 35.21 ±0.86 Bb | 26.48 ±0.73 Db | 19.19 ±0.80 Eb | 29.18 ±0.98 Cb | |
| Longevity (days) | Female | 45.85 ±1.40 Aa | 37.15 ±0.63 Da | 43.01 ±0.69 Ca | 44.68 ±0.85 Ba | 24.59 ±0.73 Ea |
| Male | 3.32 ±0.73 Cb | 5.16 ±0.37 Bb | 7.85 ±0.39 Ab | 7.78 ±0.42 Ab | 2.73 ±0.47 Db | |
| Life cycle (days) | Female | 94.23 ±1.75 Aa | 73.16 ±1.04 Ba | 70.18 ±0.88 Ca | 62.05 ±1.04 Da | 43.55 ±0.90 Ea |
| Male | 50.75 ±0.93 Ab | 40.35 ±0.86 Bb | 34.33 ±0.83 Cb | 26.97 ±0.92 Eb | 31.91 ±1.14 Db | |
| Temperature (°C) | n | Sex Ratio ** | The Pre-Adult Survival Rate * |
|---|---|---|---|
| 20 | 129 | 0.47 | 0.65 |
| 23 | 164 | 0.59 | 0.82 |
| 26 | 175 | 0.50 | 0.88 |
| 32 | 186 | 0.64 | 0.93 |
| 40 | 55 | 0.80 | 0.28 |
| Biological Parameters | Temperature (°C) | ||||
|---|---|---|---|---|---|
| 20 (n = 60) | 23 (n = 96) | 26 (n = 88) | 32 (n = 119) | 40 (n = 44) | |
| Total pre-oviposition period (days) | 73.67 ± 1.23 a * | 58.13 ± 0.85 b | 46.36 ± 0.75 c | 37.26 ± 0.78 d | 35.11 ± 0.69 e |
| Adult preoviposition period (days) | 25.34 ± 0.95 a | 22.11 ± 0.35 b | 19.19 ± 0.48 d | 19.89 ± 0.43 c | 16.16 ± 0.37 e |
| Oviposition period (days) | 4.72 ± 0.69 e | 8.55 ± 0.52 c | 16.82 ± 0.44 b | 17.97 ± 0.37 a | 6.20 ± 0.41 d |
| Post-oviposition period (days) | 15.30 ± 0.82 a | 5.90 ± 0.34 b | 5.93 ± 0.33 b | 5.82 ± 0.61 b | 3.18 ± 0.39 c |
| Fecundity | 78.37 ± 5.14 d | 118.20 ± 3.38 c | 325.02 ± 6.46 b | 355.29 ± 10.51 a | 55.89 ± 2.50 e |
| Daily reproduction (eggs/female) | 16.85 ± 1.83 c | 13.86 ± 0.65 d | 19.34 ± 0.59 b | 19.78 ± 0.64 a | 9.03 ± 0.48 e |
| Maximum daily fecundity | 20.5 | 15 | 21.31 | 21.65 | 10 |
| Hatchability (%) | 61.55 ± 2.59 e | 78.83 ± 0.60 d | 96.92 ± 0.06 b | 97.75 ± 0.07 a | 91.04 ± 0.41 c |
| Population Growth Parameters | Temperature (°C) | ||||
|---|---|---|---|---|---|
| 20 | 23 | 26 | 32 | 40 | |
| r | 0.041 ± 0.001 e * | 0.065 ± 0.000 d | 0.092 ± 0.000 b | 0.119 ± 0.000 a | 0.066 ± 0.000 c |
| λ | 1.042 ± 0.001 e | 1.067 ± 0.000 d | 1.096 ± 0.000 b | 1.126 ± 0.000 a | 1.068 ± 0.000 c |
| R0 | 23.47 ± 0.29 d | 56.73 ± 0.03 c | 143.00 ± 0.07 b | 211.36 ± 0.21 a | 12.30 ± 0.00 e |
| T | 76.47 ± 16.93 a | 62.36 ± 10.88 ab | 54.26 ± 6.93 ab | 44.43 ± 6.60 b | 38.75 ± 22.80 b |
| DT | 16.79 ± 0.34 a | 10.74 ± 0.00 b | 7.55 ± 0.00 d | 5.82 ± 0.00 e | 10.55 ± 0.00 c |
| Life Table Parameters | Temperature (°C) | ||||
|---|---|---|---|---|---|
| 20 | 23 | 26 | 32 | 40 | |
| Number of initial crawlers (N0) | 200 | 200 | 200 | 200 | 200 |
| Number of first-instar nymph female | 70 | 100 | 94 | 121 | 55 |
| Number of first-instar nymph male | 90 | 75 | 94 | 70 | 20 |
| Number of second-instar nymph female | 65 | 98 | 90 | 119 | 49 |
| Number of second-instar nymph male | 80 | 70 | 89 | 67 | 14 |
| Number developing into young female | 61 | 96 | 88 | 119 | 44 |
| Number developing into cocoon | 72 | 68 | 87 | 67 | 11 |
| Number developing into adults (A) | 133 | 164 | 175 | 186 | 55 |
| Number developing into mature female | 61 | 96 | 88 | 119 | 44 |
| Number developing into adult male | 72 | 68 | 87 | 67 | 11 |
| Predicted fecundity of next generation (G2 = A × P♀ × PF) | 4898.91 | 11,437.03 | 28,439.25 | 42,293.72 | 2459.16 |
| Population trend index I = G2/N0 | 24.49 | 57.18 | 142.20 | 211.47 | 12.30 |
| Life Stages | R2 | LDT (°C) | SET (°D) | ||
|---|---|---|---|---|---|
| a | b | ||||
| First-instar nymph female | 0.0447 | −0.9936 | 0.90 | 22.23 | 22.37 |
| First-instar nymph male | 0.0403 | −0.8893 | 0.90 | 22.07 | 24.81 |
| Second-instar nymph female | 0.0063 | −0.0987 | 0.98 | 15.67 | 158.73 |
| Second-instar nymph male | 0.0128 | −0.1852 | 0.90 | 14.47 | 78.13 |
| Young female | 0.0021 | −0.0029 | 0.99 | 1.38 | 476.19 |
| Cocoon | 0.0142 | −0.2378 | 0.90 | 16.75 | 70.42 |
| Total pre-adult female | 0.0013 | −0.0132 | 0.99 | 10.15 | 769.23 |
| Total pre-adult male | 0.0028 | −0.0342 | 0.99 | 12.21 | 357.14 |
| Adult female | 0.0013 | −0.0137 | 0.80 | 10.54 | 769.23 |
| Adult male | 0.0177 | −0.3724 | 0.82 | 21.04 | 56.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Aalaoui, M.; Sbaghi, M. Temperature Dependence for Survival, Development, and Reproduction of the Cactus Cochineal Dactylopius opuntiae (Cockerell). Insects 2022, 13, 426. https://doi.org/10.3390/insects13050426
El Aalaoui M, Sbaghi M. Temperature Dependence for Survival, Development, and Reproduction of the Cactus Cochineal Dactylopius opuntiae (Cockerell). Insects. 2022; 13(5):426. https://doi.org/10.3390/insects13050426
Chicago/Turabian StyleEl Aalaoui, Mohamed, and Mohamed Sbaghi. 2022. "Temperature Dependence for Survival, Development, and Reproduction of the Cactus Cochineal Dactylopius opuntiae (Cockerell)" Insects 13, no. 5: 426. https://doi.org/10.3390/insects13050426
APA StyleEl Aalaoui, M., & Sbaghi, M. (2022). Temperature Dependence for Survival, Development, and Reproduction of the Cactus Cochineal Dactylopius opuntiae (Cockerell). Insects, 13(5), 426. https://doi.org/10.3390/insects13050426