
The Insecticidal Efficacy and Physiological Action Mechanism of a Novel Agent GC16 against Tetranychus pueraricola (Acari: Tetranychidae)

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Pesticide
2.2. Bioassays
2.2.1. Bioassay of GC16 against Different Stages of T. pueraricola
Egg Bioassay
Larva and Nymph Bioassay
Adult Bioassay
2.2.2. Bioassays of GC16 by Different Methods
Slide-Dip Assay
Leaf-Dip Assay
Spraying Assay
2.2.3. Bioassays for the Different Components of GC16
2.3. Observation of Poisoning Symptoms for T. pueraricola
2.4. Effects of GC16 on the Morphology of Female Adult T. pueraricola
2.5. Effects of GC16 on the Egg Hatching Rate and Developmental Duration of T. pueraricola
2.6. Cryo-SEM (Scanning Electron Microscopy)
2.7. TEM (Transmission Electron Microscopy)
2.8. The Effects of GC16 on Non-Target Organisms
2.9. Statistical Analysis
3. Results
3.1. Bioassays
3.1.1. Bioassays of GC16 against Different Stages of T. pueraricola
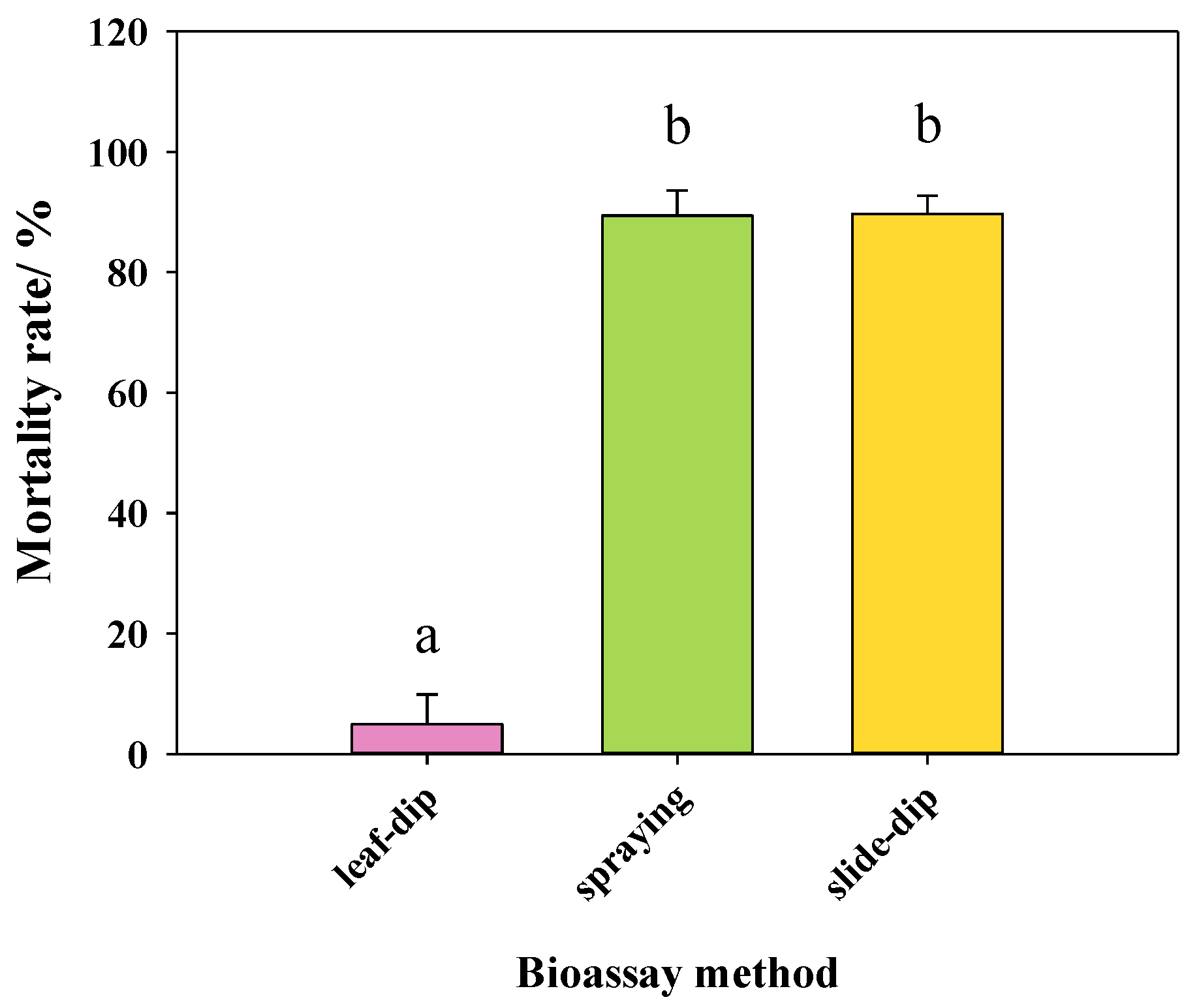
3.1.2. Bioassays of GC16 by Different Bioassay Method
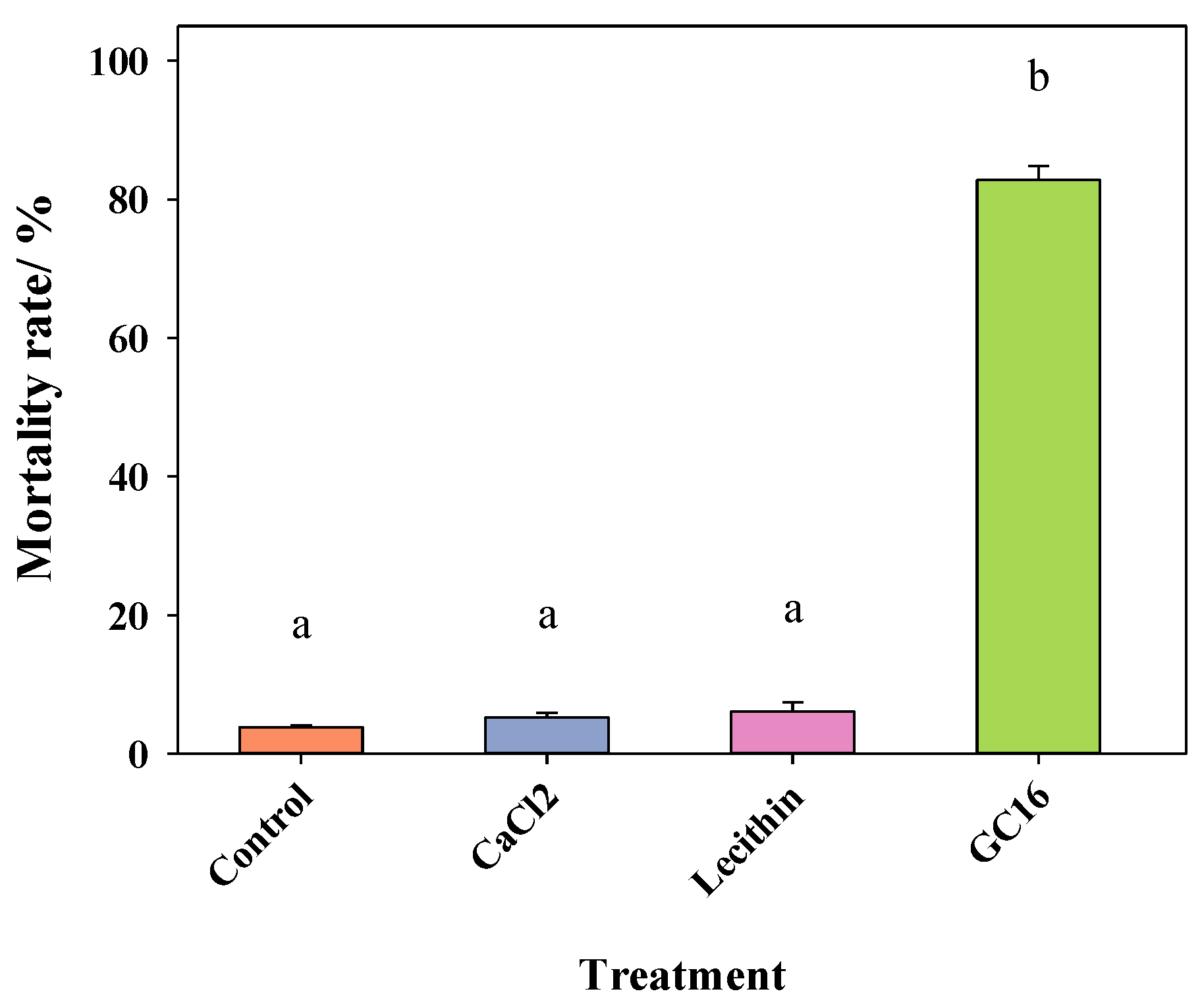
3.1.3. Bioassays for the Different Components of GC16
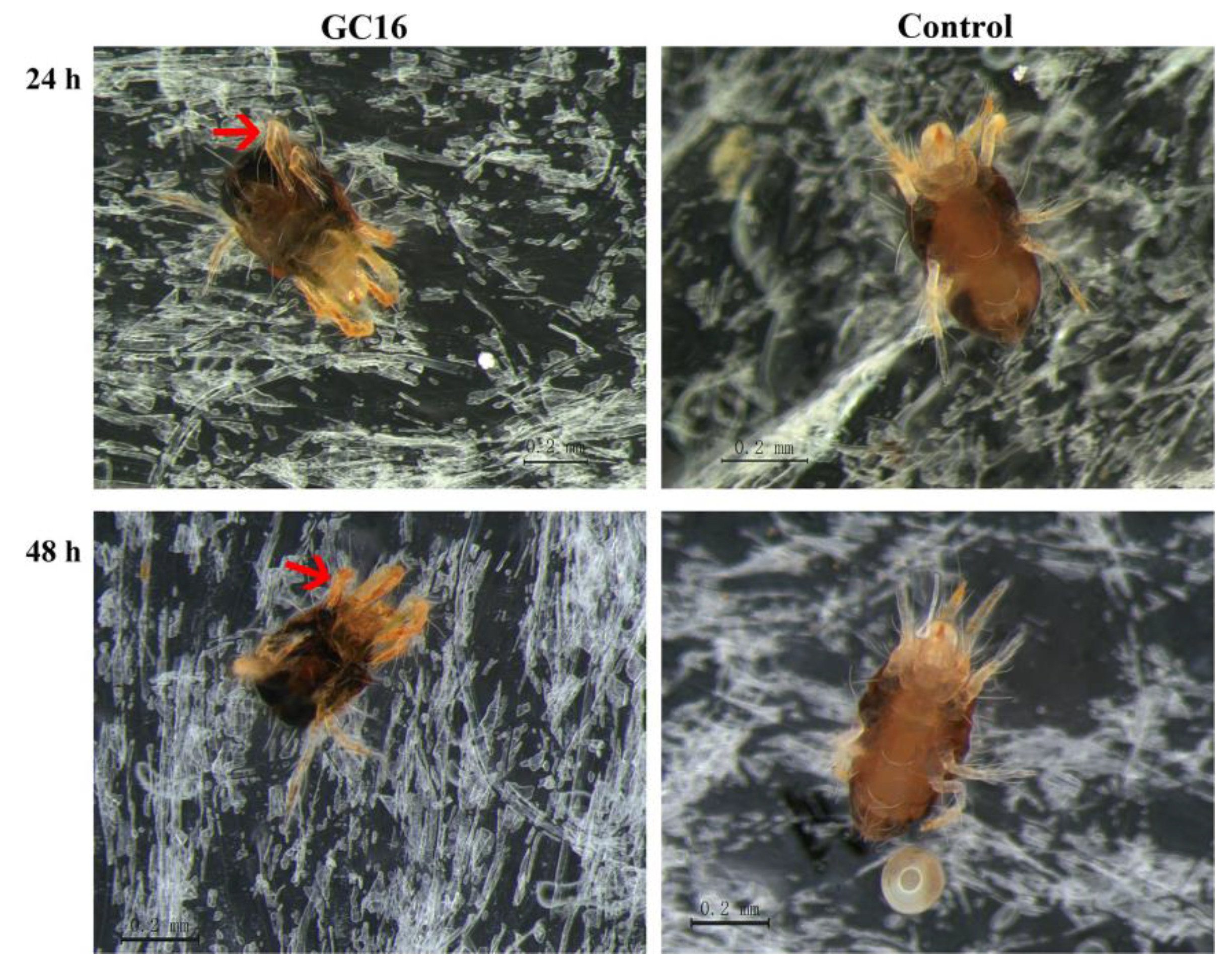
3.2. Observation of Poisoning Symptoms
3.3. Effects of GC16 on the Morphology of Female Adult T. pueraricola
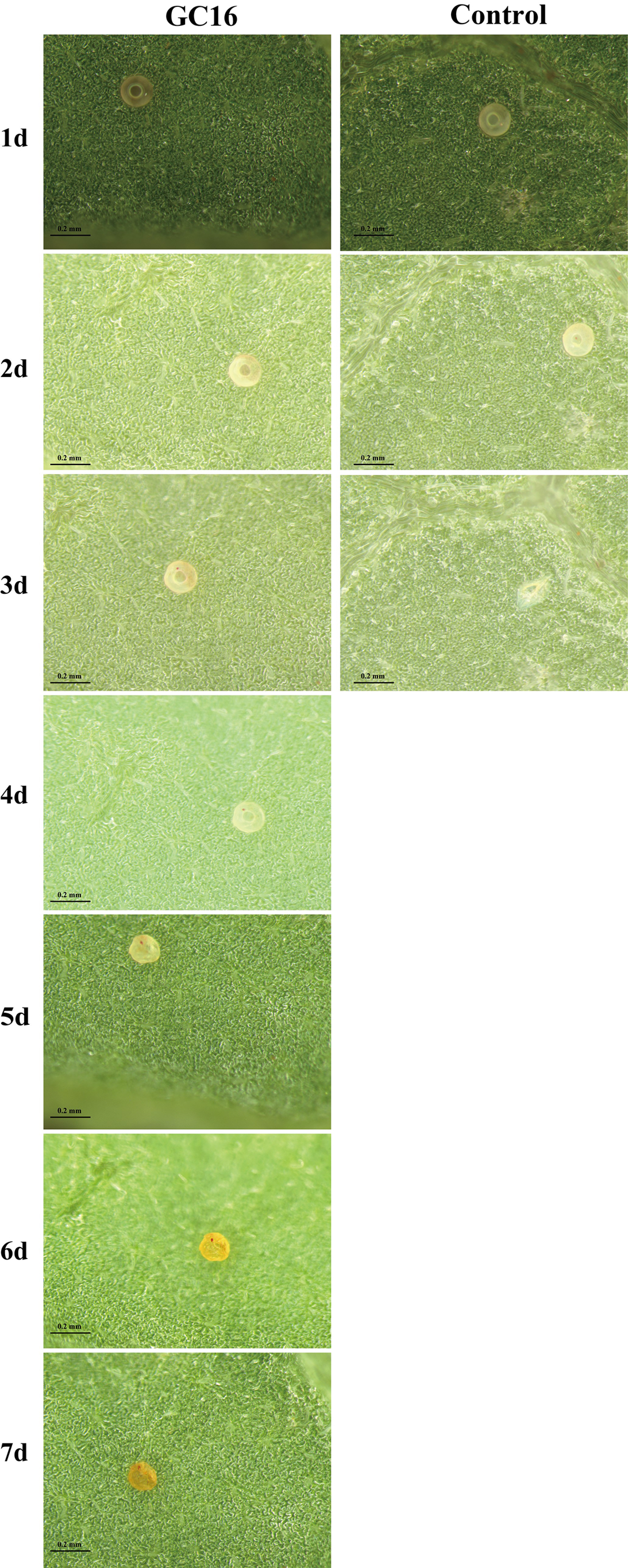
3.4. Effects of GC16 on the Hatching Rate and Developmental Duration of T. pueraricola Egg
3.5. Cryo-SEM Analysis
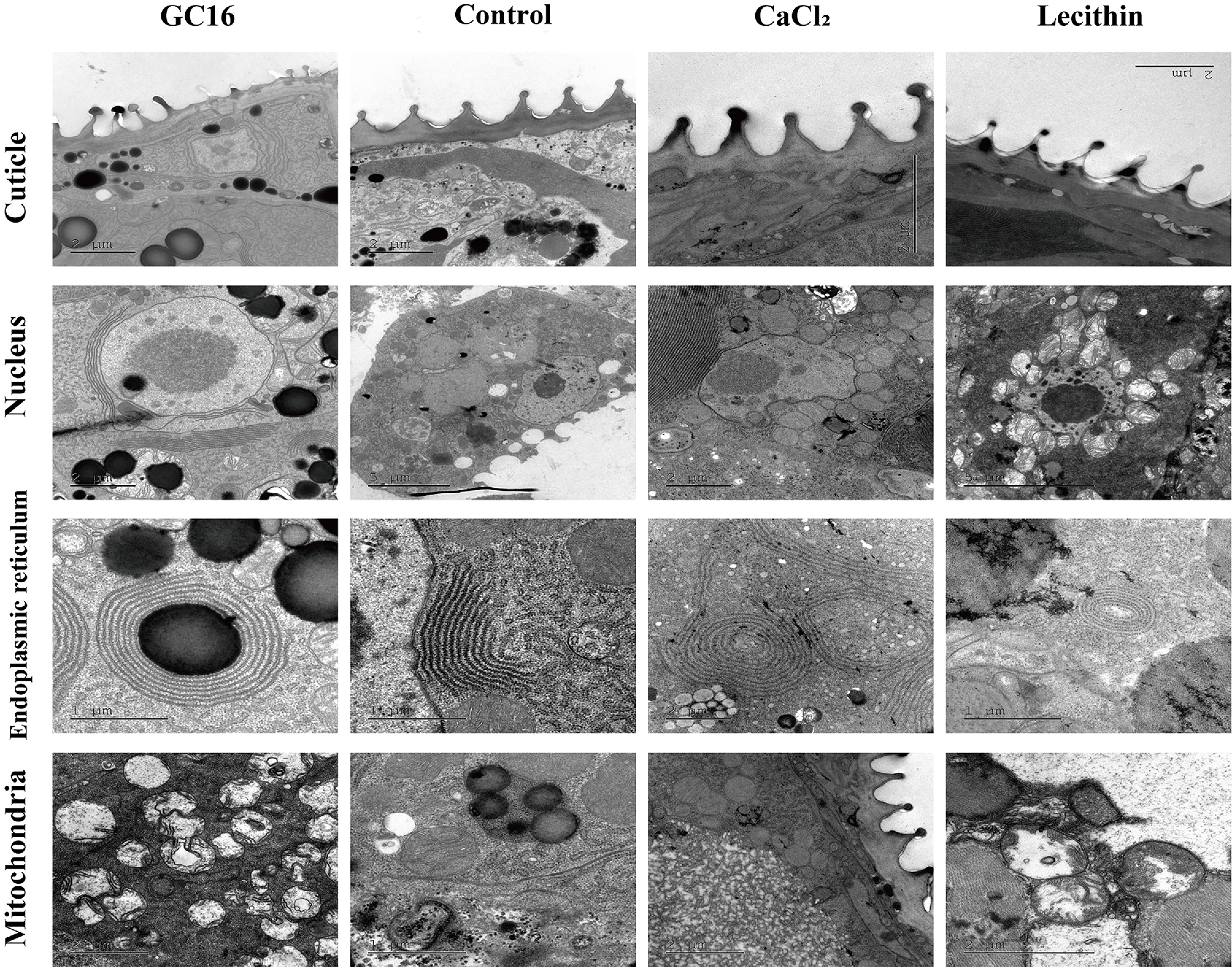
3.6. TEM Analysis
3.7. The Effects of GC16 on the Non-Target Organisms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumawat, S.; Asiwal, R.; Nitharwal, M. Agricultural sustainability: Various projects and programmes. Agric. Food E-Newsl. 2021, 3, 84–86. [Google Scholar]
- Yan, S.; Hu, Q.; Jiang, Q.H.; Chen, H.T.; Wei, J.; Yin, M.Z.; Du, X.G.; Shen, J. Simple osthole/nanocarrier pesticide efficiently controls both pests and diseases fulfilling the need of green production of strawberry. ACS Appl. Mater. Interfaces 2021, 13, 36350–36360. [Google Scholar] [CrossRef] [PubMed]
- Reganold, J.P.; Wachter, J.M. Organic agriculture in the twenty-first century. Nat. Plants 2016, 2, 15221. [Google Scholar] [CrossRef]
- Pauline, L. Sustainable agriculture for biodiversity, biodiversity for sustainable agriculture. Biodiversity 2017, 18, 124–125. [Google Scholar] [CrossRef]
- Kumar, J.K.; Monica, S.S.; Bojan, V.; Suganthi, A.; Paramasivam, M. Impact of pesticide exposure on environment and biodiversity: A review. Agric. Rev. 2021, in press. [Google Scholar] [CrossRef]
- Gotoh, T.; Suwa, A.; Kitashima, Y.; Rezk, H. Developmental and reproductive performance of Tetranychus pueraricola Ehara and Gotoh (Acari: Tetranychidae) at four constant temperatures. Appl. Entomol. Zool. 2004, 39, 675–682. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, T.; Suwa, A.; Kitashima, Y. Development and oviposition of Tetranychus pueraricola ehara and gotoh (acari: Tetranychidae) on various plants. J. Acarol. Soc. Jpn. 2004, 13, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Ilias, A.; Vontas, J.; Tsagkarakou, A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef]
- Ilias, A.; Vassiliou, V.A.; Vontas, J.; Tsagkarakou, A. Molecular diagnostics for detecting pyrethroid and abamectin resistance mutations in Tetranychus urticae. Pestic. Biochem. Physiol. 2017, 135, 9–14. [Google Scholar] [CrossRef]
- Jin, P.Y.; Tian, L.; Chen, L.; Hong, X.Y. Spider mites of agricultural importance in China, with focus on species composition during the last decade (2008–2017). Syst. Appl. Acarol. 2018, 23, 2087–2098. [Google Scholar] [CrossRef]
- Van, N.P.; Van, L.T.; Khajehali, J.; Vanholme, B.; Tirry, L. Mutations in the mitochondrial cytochrome b of Tetranychus urticae Koch (Acari:Tetranychidae) confer cross-resistance between bifenazate and acequinocyl. Pest Manag. Sci. 2009, 65, 404–412. [Google Scholar] [CrossRef]
- Vassiliou, V.A.; Kitsis, P. Acaricide resistance in Tetranychus urticae (Acari:Tetranychidae) populations from Cyprus. J. Econ. Entomol. 2013, 106, 1848–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, Y.J.; Xie, W.; Wu, Q.J.; Wang, S.L. Sublethal effects of spinetoram on the two-spotted spider mite, Tetranychus urticae (Acari: Tetranychidae). Pestic. Biochem. Physiol. 2016, 132, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.D.; He, Y.Y.; Zhang, Y.J.; Xie, W.; Wu, Q.J.; Wang, S.L. Status of pesticide resistance and associated mutations in the two-spotted spider mite, Tetranychus urticae, in China. Pestic. Biochem. Physiol. 2018, 150, 89–96. [Google Scholar] [CrossRef]
- Ionel, I.L.; Mara, G.; Stefania, R.; Margarita, G.O.; Corina, P. A hazard to human health-pesticide residues in some vegetal and animal foodstuff. J. Biotechnol. 2019, 305, S22–S23. [Google Scholar] [CrossRef]
- Shahid, E.; Khan, D.J.; Qaisrani, M.; Noman, M.; Rani, A.; Ali, S. Effect of pesticide residues on agriculture crops. J. Toxicol. Pharmaceut. Sci. 2021, 5, 18–23. [Google Scholar]
- (USTCA) Ulanqab Science and Technology Commissioners Association. The scientific and technological achievements of GC16 ecological preparations have taken root in Hainan. China Rural Sci. Technol. 2021, 5, 60–61. [Google Scholar]
- Hou, Q.L.; Zhang, Y.Q.; Li, C.X.; Ding, W.; Liu, X.J.; Luo, J.X. Acaricidal toxicity of scopoletin combined with Ca2+ and its influence on Ca2+-ATPase activity in Tetranychus cinnabarinus (Boisduval). Chin. J. Pestic. Sci. 2015, 17, 475–479. [Google Scholar] [CrossRef]
- Chen, J.; Ma, H.W.; Zhang, P.; Liu, M.T. Preparation of potassium chloride from K-feldspar with calcium chloride as additive agent: A review. Chem. Ind. Eng. Prog. 2016, 35, 3954–3963. [Google Scholar] [CrossRef]
- Ding, X.T.; Jiang, J.G.; Li, D.A.; Li, T.R.; Wang, J.M. Immobilizing effects of calcium-based agents on soil contaminated by vanadium ore. J. Agro-Environ. Sci. 2016, 35, 274–280. [Google Scholar] [CrossRef]
- Alhajj, M.; Montero, N.; Yarce, C.; Salamanca, C. Lecithins from vegetable, land, and marine animal sources and their potential applications for cosmetic, food, and pharmaceutical sectors. Cosmetics 2020, 7, 87. [Google Scholar] [CrossRef]
- Szuhaj, B.; Yeo, J.D.; Shahidi, F. Lecithins. In Bailey’s Industrial Oil and Fat Products; Shahidi, F., Ed.; Wiley: Hoboken, NJ, USA, 2020; pp. 1–86. [Google Scholar] [CrossRef]
- Younes, M.; Aquilina, G.; Castle, L.; Engel, K.H.; Fowler, P.; Frutos, M.J.; Furst, P.; Rainer, G.; Gundert-Remy, U.; Trine, H.; et al. Safety of use of oat lecithin as a food additive. EFSA J. 2020, 18, 5969. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.Z.; Yan, W.Q.; Zhou, Y.L.; Fan, L.P.; Liu, Y.F.; Li, J.W. Progress in the application of lecithins in water-in-oil emulsions. Trends Food Sci. Technol. 2021, 118, 388–398. [Google Scholar] [CrossRef]
- Velasco, L.; Rascón, M.; Calvo, M.V.; Montalvo, R.; Fontecha, J.; Garcia, H.K. Krill lecithin as surfactant for preparation of oil/water nanoemulsions as curcumin carriers. Eur. J. Lipid Sci. Technol. 2021, 123, 2000238. [Google Scholar] [CrossRef]
- Kumar, N.; Minhas, P.S.; Ambasankar, K.; Krishnani, K.K.; Rana, R.S. Dietary lecithin potentiates thermal tolerance and cellular stress protection of milk fish (Chanos Chanos) reared under low dose endosulfan-induced stress. J. Therm. Biol. 2014, 46, 40–46. [Google Scholar] [CrossRef]
- Djekic, L.; Krajisnik, D.; Martinovic, M.; Djordjevic, D.; Primorac, M. Characterization of gelation process and drug release profile of thermosensitive liquid lecithin/poloxamer 407 based gels as carriers for percutaneous delivery of ibuprofen. Int. J. Pharm. 2015, 490, 180–189. [Google Scholar] [CrossRef]
- Schirra, M.; D’Aquino, S.; Migheli, Q.; Pirisi, F.M.; Angioni, A. Influence of post-harvest treatments with fludioxonil and soy lecithin co-application in controlling blue and grey mould and fludioxonil residues in coscia pears. Food Addit. Contam. Part A 2009, 26, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Ozgen, M.; Palta, J.P. A natural lipid, lysophosphatidylethanolamine (lpe), can mitigate adverse effect of fungicide, chlorothalonil, on fruit set and yield in cranberries. Acta Hortic. 2003, 628, 747–752. [Google Scholar] [CrossRef]
- Jin, P.Y.; Tian, L.; Chen, L.; Hong, X.Y. High genetic diversity in a ‘recent outbreak’ spider mite, Tetranychus pueraricola, in mainland China. Exp. Appl. Acarol. 2019, 78, 15–27. [Google Scholar] [CrossRef]
- Ehara, S.; Gotoh, T. Two new species of spider mites occurring in Japan (Acari: Tetranychidae). J. Acarol. Soc. Jpn. 1996, 5, 17–25. [Google Scholar] [CrossRef]
- Suwa, A.; Gotoh, T. Geographic variation in diapause induction and mode of diapause inheritance in Tetranychus pueraricola. J. Appl. Entomol. 2006, 130, 329–335. [Google Scholar] [CrossRef]
- Kabir, K.H.; Chapman, R.B. Operational and biological factors influencing responses of spider mites (Acari: Tetranychidae) to propargite by using the Petri dish-Potter tower method. J. Econ. Entomol. 1997, 90, 272–277. [Google Scholar] [CrossRef]
- Ay, R. Determination of susceptibility and resistance of some greenhouse populations of Tetranychus urticae Koch to chlorpyrifos (Dursban 4) by the petri dish-Potter tower method. J. Pest Sci. 2005, 78, 139–143. [Google Scholar] [CrossRef]
- Tang, X.F.; Zhang, Y.J.; Wu, Q.J.; Xie, W.; Wang, S.L. Stage-specific expression of resistance to different acaricides in four field populations of Tetranychus urticae (Acari: Tetranychidae). J. Econ. Entomol. 2014, 107, 1900–1907. [Google Scholar] [CrossRef] [PubMed]
- Amer, M.; Mekky, H.; Fedawy, H. Effect of combined plant essential oils on Dermanyssus gallinae: In vitro and in vivo study. J. World’s Poult. Res. 2020, 10, 199–206. [Google Scholar] [CrossRef]
- Cheng, S.; Lin, R.; You, Y.; Lin, T.; Zeng, Z.; Yu, C. Comparative sensitivity of Neoseiulus cucumeris and its prey Tetranychus cinnabarinus, after exposed to nineteen pesticides. Ecotoxicol. Environ. Saf. 2021, 217, 112234. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, V.; Cranham, J.E.; Jepson, L.R.; Helle, W. Revised method for spider mites and their eggs (e.g., Tetranychus spp. and Panonychus ulmi Koch), FAO method No. 10a. FAO Plant Prod. Prot. Pap. 1980, 21, 49–53. [Google Scholar]
- Wang, L.; Zhang, Y.J.; Xie, W.; Wu, Q.J.; Wang, S.L. A bioassay for evaluation of the resistance of Tetranychus urticae (Acari: Tetranychidae) to selected acaricides. Syst. Appl. Acarol. 2015, 20, 579–590. [Google Scholar] [CrossRef]
- Tarelli, G.; Zerba, E.; Alzogaray, R. Toxicity to vapor exposure and topical application of essential oils and monoterpenes on Musca domestica (Diptera: Muscidae). J. Econ. Entomol. 2009, 102, 1383–1388. [Google Scholar] [CrossRef]
- Qiu, Y.; Song, X.H.; Huang, Y.Z. Behavior response and insecticidal mechanism of β-Asarone against Sitophilus Zeamais Adult. J. Chin. Cereals Oils Assoc. 2014, 29, 80–85. [Google Scholar]
- Pashte, V.; Patil, S.C. Toxicity and poisoning symptoms of selected insecticides to honey bees (Apis mellifera mellifera L.). Arch. Biol. Sci. 2017, 70, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Li, C.X.; Zhang, Y.Q.; Wang, D.; Zhang, B.C.; Luo, J.X.; Ding, W. Effects of curcumin and scopoletinon morphological change and water loss of Tetranychus cinnabarinus Boisduval (Acarina: Tetranychidae). J. Southwest Univ. 2017, 39, 10–15. [Google Scholar] [CrossRef]
- Walther, P. Cryo-SEM and TEM of high pressure frozen Cells-some technical contributions. Microsc. Microanal. 2001, 7, 728–729. [Google Scholar] [CrossRef]
- Yu, Z.M.; Kang, B.; He, X.W.; Lv, S.L.; Bai, Y.H.; Ding, W.N.; Chen, M.; Cho, H.T.; Wu, P. Root hair-specific expansins modulate root hair elongation in rice. Plant J. 2011, 66, 725–734. [Google Scholar] [CrossRef]
- Yan, H.T.; Wang, Y.; Zhang, J.R.; Cui, X.R.; Wu, J.S.; Zhou, J.; Lu, J.; Guo, R.Y.; Ou, M.; Lai, H.X.; et al. Rice root hair phenotypes imaged by Cryo-SEM. Bio-Protocol 2021, 11, e4037. [Google Scholar] [CrossRef]
- Parastou, T.; Cecilia, C.; Negar, K. Sample Preparation for Transmission Electron Microscopy. Methods Mol. Biol. 2019, 1897, 417–424. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Li, M.J.; Yang, C.L.; Wang, X.C. M05B5.4 (Lysosomal phospholipase A2) promotes disintegration of autophagic vesicles to maintain C. elegans development. Autophagy 2021, 18, 595–607. [Google Scholar] [CrossRef]
- Ma, S.C.; Jiang, W.Q.; Li, Q.; Li, T.; Wu, W.J.; Bai, H.Y.; Shi, B.J. Design, synthesis, and study of the insecticidal activity of novel steroidal 1,3,4-Oxadiazoles. J. Agric. Food Chem. 2021, 69, 11572–11581. [Google Scholar] [CrossRef]
- Ma, T.F.; Zhao, L.Y.; Zhang, J.; Tang, R.F.; Wang, X.; Liu, N.; Zhang, Q.; Wang, F.Y.; Li, M.J.; Shan, Q.; et al. A pair of transporters controls mitochondrial Zn2+ levels to maintain mitochondrial homeostasis. Protein Cell 2021, 13, 180–202. [Google Scholar] [CrossRef]
- Zhang, Q.; Ding, L.J.; Li, M.; Cui, W.W.; Ding, W.; Luo, J.X.; Zhang, Y.Q. Action modes of Aloe vera L. extracts against Tetranychus cinnabarinus Boisduval (Acarina: Tetranychidae). Agric. Sci. 2013, 4, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Liu, S.; Wan, F.; Jian, Y.; Guo, F.; Chen, J.; Ning, Y.S.; Ding, W. Graphene oxide-acaricide nanocomposites advance acaricidal activity of acaricides against Tetranychus cinnabarinus by directly inhibiting the transcription of a cuticle. Environ. Sci. Nano 2021, 8, 3122–3137. [Google Scholar] [CrossRef]
- Zhou, H.; Wan, F.; Guo, F.; Liu, J.; Ding, W. High value-added application of a renewable bioresource as acaricide: Investigation the mechanism of action of scoparone against Tetranychus cinnabarinus. J. Adv. Res. 2021, in press. [CrossRef]
- Zhang, J.; Feng, G.; Ma, Z.Q.; Feng, J.T.; Zhang, X. Toxicity of sarisan against Mythimna separata Walker and its effects on AChE and ATPases. Acta Entomol. Sin. 2007, 50, 574–577. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Ding, W.; Zhao, Z.M.; Wu, J.; Fan, Y.H. Studies on acaricidal bioactivities of Artemisia annua L. extracts against Tetranychus cinnabarinus Bois. (Acari: Tetranychidae). Agric. Sci. China 2008, 7, 577–584. [Google Scholar] [CrossRef]
- Subhomay, S.; Suresh, W.; Jitendra, K.; Panwar, V.P.S.; Parmar, B.S. Ovicidal and egg hatching inhibiting activity of azadirachtin A, B, and H against the maize stem borer Chillo partellus (Swinhoe). Pestic. Res. J. 2005, 17, 6–9. [Google Scholar]
- Zhou, H.; Liu, J.L.; Wan, F.L.; Guo, H.; Ning, Y.S.; Liu, S.S.; Ding, W. Insight into the mechanism of action of scoparone inhibiting egg development of Tetranychus cinnabarinus Boisduval. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 246, 109055. [Google Scholar] [CrossRef]
- Chen, N.; Pei, X.J.; Li, S.; Fan, Y.L.; Liu, T.X. Involvement of integument-rich CYP4G19 in hydrocarbon biosynthesis and cuticular penetration resistance in Blattella germanica (L.). Pest Manag. Sci. 2020, 76, 215–226. [Google Scholar] [CrossRef]
- Su, C.; Xia, X. Sublethal effects of methylthio-diafenthiuron on the life table parameters and enzymatic properties of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae). Pestic. Biochem. Physiol. 2020, 162, 43–51. [Google Scholar] [CrossRef]
- Echlin, P. The application of scanning electron microscopy to biological research. Philo. T R. Soc. B 1971, 261, 51–59. [Google Scholar] [CrossRef]
- Noble-Nesbitt, J. Structural aspects of penetration through insect cuticles. Pestic. Sci. 1970, 1, 204–208. [Google Scholar] [CrossRef]
- Johnson, R.A.; Kaiser, A.; Quinlan, M.; Sharp, W. Effect of cuticular abrasion and recovery on water loss rates in queens of the desert harvester ant Messor pergandei. J. Exp. Biol. 2011, 214, 3495–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Shi, B.; Zhao, Y.; Hu, Z.N.; Wu, W.J. Histopathological effects and immunolocalization of periplocoside NW from Periploca sepium Bunge on the midgut epithelium of Mythimna separata Walker larvae. Pestic. Biochem. Physiol. 2014, 115, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Jiang, H.; Zhao, Y.; He, J.; Cheng, H.G.; Martyniuket, C.J. A comprehensive review of 1,2,4-triazole fungicide toxicity in zebrafish (Danio rerio): A mitochondrial and metabolic perspective. Sci. Total Environ. 2021, 809, 151177. [Google Scholar] [CrossRef]
- Nobuyoshi, T.; Hirofumi, N.; Yasuhiro, S.; Naoki, I. Development of a new acaricide, cyflumetofen. J. Pestic. Sci. 2012, 37, 263–264. [Google Scholar] [CrossRef] [Green Version]
- Del Valle, I.; Gómez-Durán, A.; Holt, W.V.; Muiño-Blanco, T.; Cebrián-Pérez, J.A. Soy lecithin Interferes with mitochondrial function in frozen-thawed ram spermatozoa. J. Androl. 2012, 33, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Guo, F.; Luo, J.; Zhang, Y.; Liu, J.; Zhang, Y.C.; Zheng, X.Y.; Wan, F.L.; Ding, W. Functional analysis of an upregulated calmodulin gene related to the acaricidal activity of curcumin against Tetranychus cinnabarinus (Boisduval). Pest Manag. Sci. 2021, 77, 719–730. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Y.; Lai, T.; Liu, X.; Guo, F.; Guo, T.; Ding, W. Acaricidal mechanism of scopoletin against Tetranychus cinnabarinus. Front. Physiol. 2019, 10, 164. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.F.; Zhang, Y.Y.; Zhou, H.; Liu, J.L.; Guo, H.; Luo, J.X.; Ding, W.; Zhang, Y.Q. Silencing T-type voltage-gated calcium channel gene reduces the sensitivity of Tetranychus cinnabarinus (Boisduval) to scopoletin. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 227, 108644. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, Y.; Lai, T.; Wang, D.; Liu, J.L.; Guo, H.; Ding, W. Silencing chitinase genes increases susceptibility of Tetranychus cinnabarinus (Boisduval) to Scopoletin. BioMed Res. Int. 2017, 7, 9579736. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | N a | Slope ± SE | LC50 (95% FL b(g/L)) | LC90 (95% FL b(g/L)) | χ2(df) c |
|---|---|---|---|---|---|
| egg | 425 | 1.187 ± 0.182 | 2.239 (1.349 ~ 3.131) | 26.888 (16.980 ~ 58.918) | 2.615 (10) |
| larva | 885 | 2.652 ± 0.202 | 1.963 (1.655 ~ 2.300) | 5.973 (4.759 ~ 8.245) | 21.988 (13) |
| nymph | 686 | 1.907 ± 0.199 | 1.266 (0.782 ~ 1.705) | 5.951 (4.044 ~ 12.833) | 39.524 (13) |
| adult | 625 | 2.107 ± 0.193 | 1.996 (1.624 ~ 2.396) | 8.099 (6.143 ~ 12.155) | 17.138 (13) |
| Treatment (g/L) | 20 min | 40 min | 2 h | 4 h | 6 h | 8–18 h | 18–24 h |
|---|---|---|---|---|---|---|---|
| Control (0) | feed and oviposit actively | feed and oviposit actively | feed and oviposit actively | feed and oviposit actively | feed and oviposit actively | feed and oviposit actively | feed and oviposit actively |
| GC16 (2.00) | stationary | stationary | moved slightly | moved slightly | 1. crawled 2. moved slightly 3. stationary/died | 1. crawled 2. moved slightly 3. died | 1. crawled 2. wiggled when tapped 3. died |
| GC16 (6.67) | stationary | stationary | stationary | stationary/died | stationary/died | died, started shrinking | shriveled |
| Treatment | Body Length (mm) | Rst for Body Length | Body Width (mm) | Rst for Body Width |
|---|---|---|---|---|
| Control-24 h | 0.57 ± 0.01 a | 24 h Rst = 14.36% | 0.28 ± 0.01 a,b | 24 h Rst = −0.16% |
| GC16-24 h | 0.49 ± 0.01 b | 0.28 ± 0.01 a,b | ||
| Control-48 h | 0.56 ± 0.01 a | 48 h Rst = 24.69% | 0.30 ± 0.01 a | 48 h Rst = 14.53% |
| GC16-48 h | 0.42 ± 0.01 c | 0.26 ± 0.01 b |
| Organism | Mortality Rate (%) | t, p | |
|---|---|---|---|
| GC16 | Control | ||
| Tetranychus pueraricola | 86.21 ± 2.02 A,a | 2.38 ± 1.19 A,b | t = −35.693, df = 4, p < 0.001 |
| Picromerus lewisi | 4.15 ± 0.23 B,a | 4.35 ± 0.37 A,a | t = 0.447, df = 4, p = 0.678 |
| Harmonia axyridis | 8.33 ± 1.67 B,a | 5.83 ± 0.83 A,a | t = −1.342, df = 4, p = 0.251 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Du, G.; Xie, S.; Long, X.; Sun, G.; Zhu, S.; He, X.; Liu, Y.; Zhu, Y.; Chen, B. The Insecticidal Efficacy and Physiological Action Mechanism of a Novel Agent GC16 against Tetranychus pueraricola (Acari: Tetranychidae). Insects 2022, 13, 433. https://doi.org/10.3390/insects13050433
He Y, Du G, Xie S, Long X, Sun G, Zhu S, He X, Liu Y, Zhu Y, Chen B. The Insecticidal Efficacy and Physiological Action Mechanism of a Novel Agent GC16 against Tetranychus pueraricola (Acari: Tetranychidae). Insects. 2022; 13(5):433. https://doi.org/10.3390/insects13050433
Chicago/Turabian StyleHe, Yanyan, Guangzu Du, Shunxia Xie, Xiaoming Long, Ganlin Sun, Shusheng Zhu, Xiahong He, Yixiang Liu, Youyong Zhu, and Bin Chen. 2022. "The Insecticidal Efficacy and Physiological Action Mechanism of a Novel Agent GC16 against Tetranychus pueraricola (Acari: Tetranychidae)" Insects 13, no. 5: 433. https://doi.org/10.3390/insects13050433
APA StyleHe, Y., Du, G., Xie, S., Long, X., Sun, G., Zhu, S., He, X., Liu, Y., Zhu, Y., & Chen, B. (2022). The Insecticidal Efficacy and Physiological Action Mechanism of a Novel Agent GC16 against Tetranychus pueraricola (Acari: Tetranychidae). Insects, 13(5), 433. https://doi.org/10.3390/insects13050433