
Using Autumnal Trap Crops to Manage Tarnished Plant Bugs (Lygus lineolaris)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
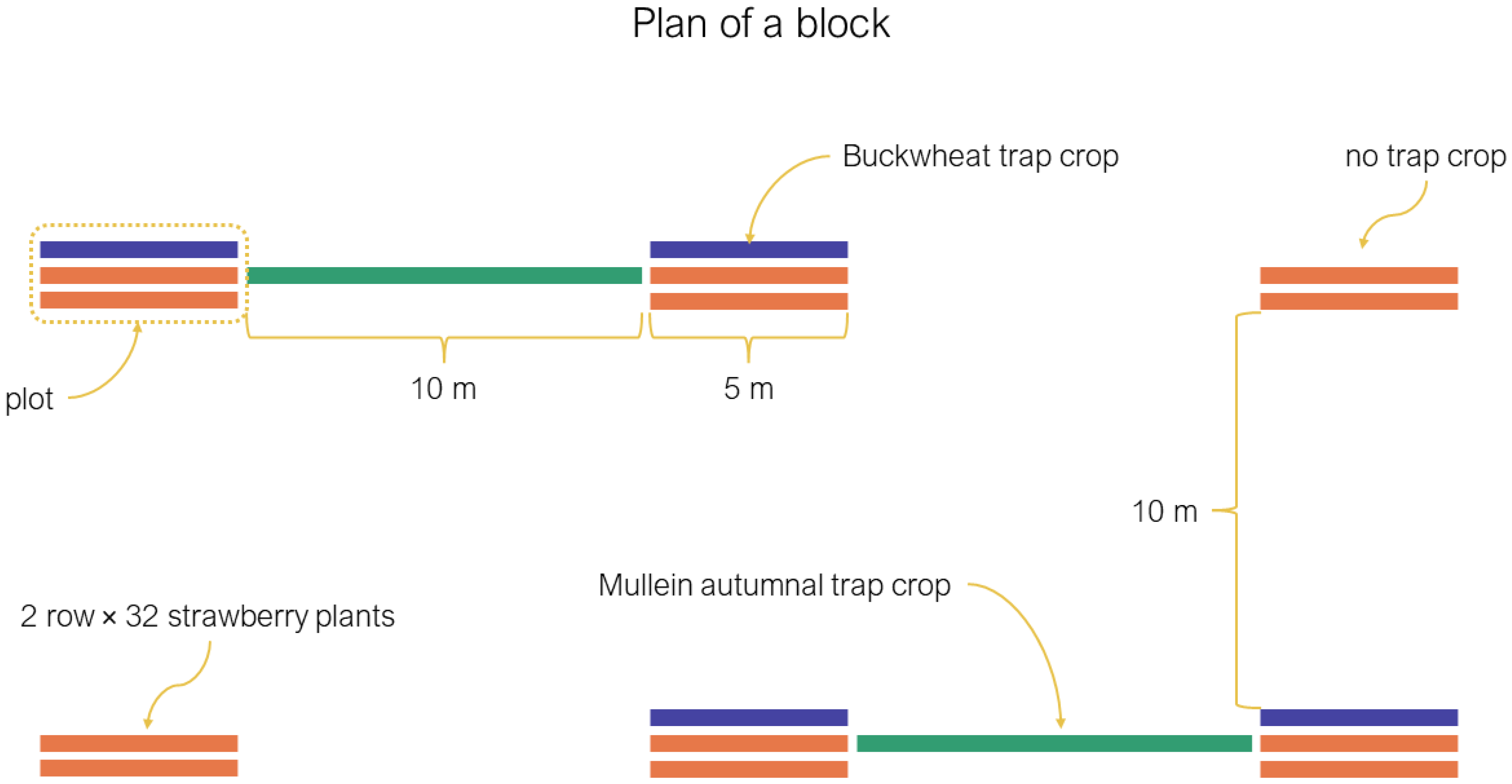
2.2. Experimental Design and Data Collection
2.2.1. Summer Trap Crop Plots
2.2.2. Autumn Trap Crop Plots
2.2.3. Winter Survivorship
2.3. Statistical Analysis
2.3.1. Population Density on Mullein Plant
2.3.2. The Peak of Population Density on Summer and Autumnal Hosts
2.3.3. Winter Survivorship
3. Results
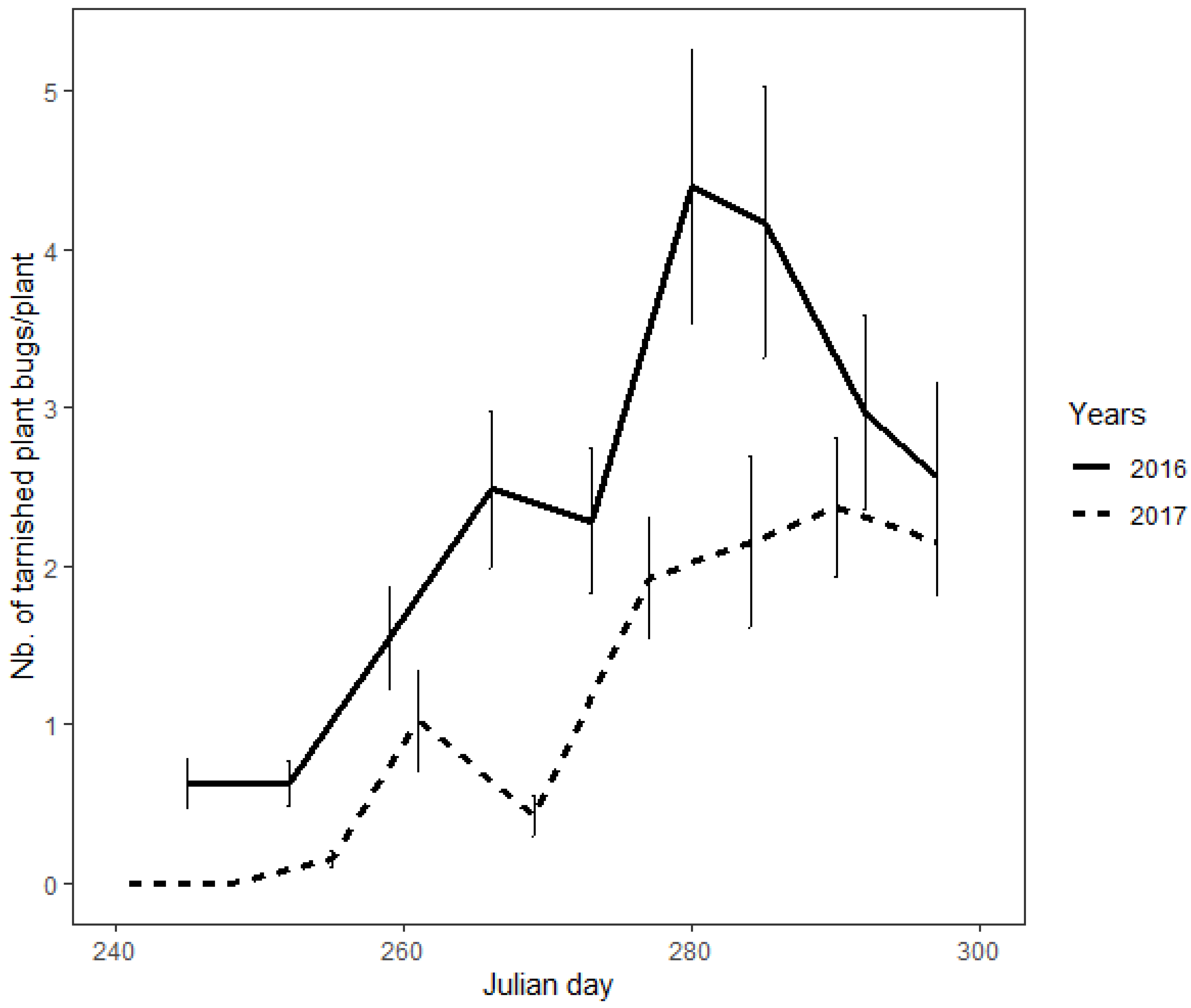
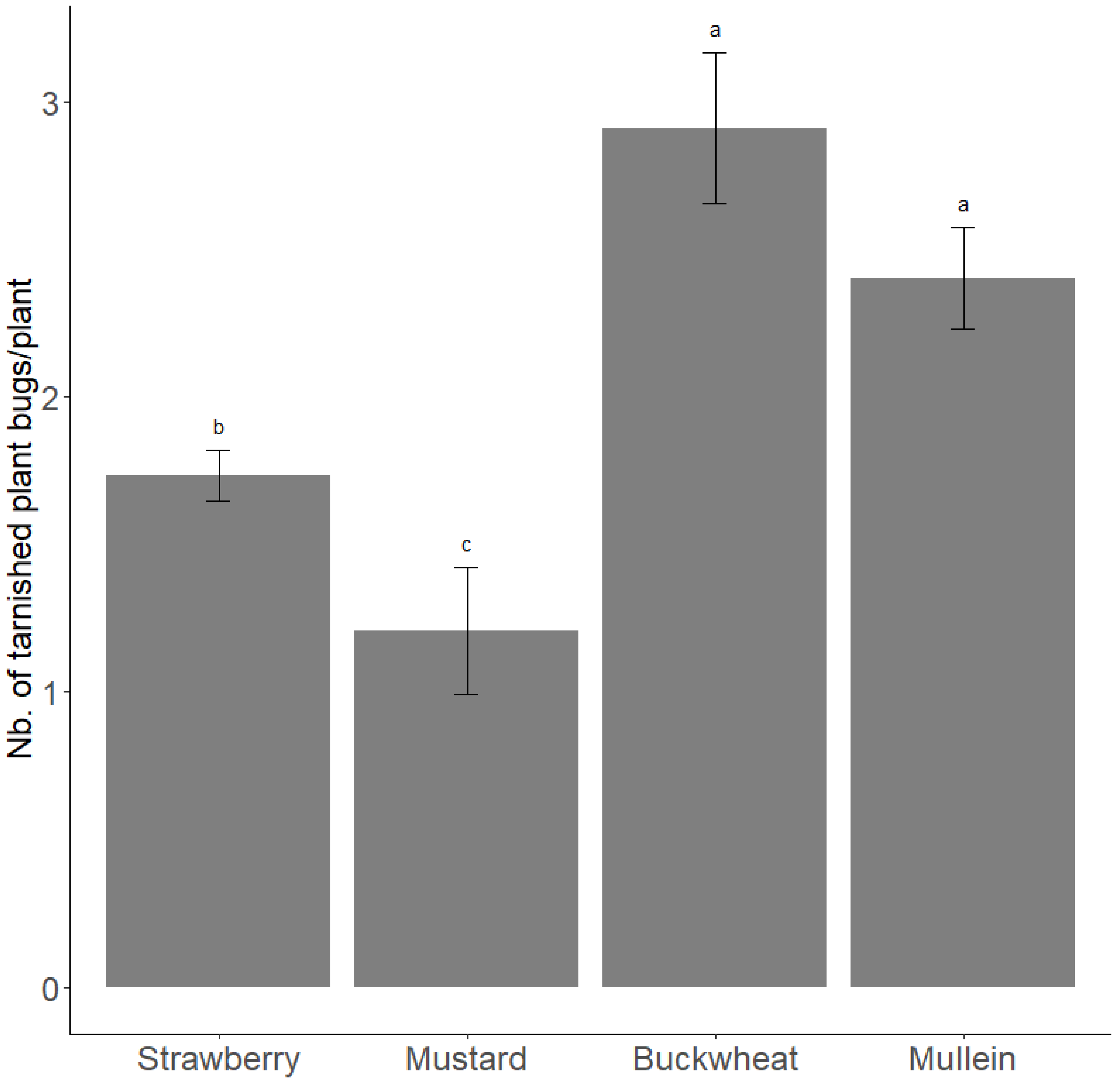
3.1. Population Density on Mullein Plant
3.2. The Peak of Population Density on Summer and Autumnal Hosts
3.3. Winter Survivorship
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shelton, A.M.; Badenes-Perez, F.R. Concepts and Applications of Trap Cropping in Pest Management. Annu. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swezey, S.L.; Nieto, D.J.; Bryer, J.A. Control of Western Tarnished Plant Bug Lygus hesperus Knight (Hemiptera: Miridae) in California Organic Strawberries Using Alfalfa Trap Crops and Tractor-Mounted Vacuums. Environ. Entomol. 2007, 36, 9. [Google Scholar] [CrossRef]
- Dumont, F.; Provost, C. Combining the Use of Trap Crops and Insecticide Sprays to Control the Tarnished Plant Bug in Strawberry Fields. Can. Entomol. 2019, 151, 1–9. [Google Scholar] [CrossRef]
- Hokkanen, H.M. Trap Cropping in Pest Management. Annu. Rev. Entomol. 1991, 36, 119–138. [Google Scholar] [CrossRef]
- Bale, J.S. Insects at Low Temperature: A Predictable Relationship? Funct. Ecol. 1991, 5, 291–298. [Google Scholar] [CrossRef]
- Danks, H.V. Modes of Seasonal Adaptation in the Insects: I. Winter Survival. Can. Entomol. 1978, 110, 1167–1205. [Google Scholar] [CrossRef]
- Tauber, C.A.; Tauber, M.J. Insect Seasonal Cycles: Genetics and Evolution. Annu. Rev. Ecol. Syst. 1981, 12, 281–308. [Google Scholar] [CrossRef]
- Turnock, W.J.; Fields, P.G. Winter Climates and Coldhardiness in Terrestrial Insects. Eur. J. Entomol. 2005, 102, 561. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Meeting the Energetic Demands of Insect Diapause: Nutrient Storage and Utilization. J. Insect Physiol. 2007, 53, 760–773. [Google Scholar] [CrossRef]
- Sinclair, B.J. Linking Energetics and Overwintering in Temperate Insects. J. Therm. Biol. 2015, 54, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Villavaso, E.J.; Snodgrass, G.L. Diapause in Tarnished Plant Bug (Heteroptera: Miridae) Reared in Dynamic Photoperiod Environmental Cabinets. J. Agric. Urban Entomol. 2004, 21, 87. [Google Scholar]
- Danks, H.V. Winter Habitats and Ecological Adaptations for Winter Survival. In Insects at Low Temperature; Springer: Berlin/Heidelberg, Germany, 1991; pp. 231–259. [Google Scholar]
- Carriere, Y.; Deland, J.-P.; Roff, D.A.; Vincent, C. Life-History Costs Associated with the Evolution of Insecticide Resistance. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1994, 258, 35–40. [Google Scholar]
- Layton, M.B. Biology and Damage of the Tarnished Plant Bug, Lygus lineolaris, in Cotton. Southwest. Entomol. 2000, 23, 7–20. [Google Scholar]
- Hardman, J.M.; Jensen, K.I.; Moreau, D.L.; Franklin, J.L.; Bent, E.D. Effects of Ground Cover Treatments and Insecticide Use on Population Density and Damage Caused by Lygus lineolaris (Heteroptera: Miridae) in Apple Orchards. J. Econ. Entomol. 2004, 97, 993–1002. [Google Scholar] [CrossRef]
- Stern, V.; Mueller, A.; Sevacherian, V.; Way, M. Lygus Bug Control in Cotton through Alfalfa Interplanting. Calif. Agric. 1969, 23, 8–10. [Google Scholar]
- Wheeler, A.G. Studies on the Arthropod Fauna of Alfalfa: VI. Plant Bugs (Miridae). Can. Entomol. 1974, 106, 1267–1275. [Google Scholar] [CrossRef]
- Cleveland, T.C. Hibernation and Host Plant Sequence Studies of Tarnished Plant Bugs, Lygus lineolaris, in the Mississippi Delta. Environ. Entomol. 1982, 11, 1049–1052. [Google Scholar] [CrossRef]
- Sanchez, J.A.; Gillespie, D.R.; McGregor, R.R. The Effects of Mullein Plants (Verbascum thapsus) on the Population Dynamics of Dicyphus Hesperus (Heteroptera: Miridae) in Tomato Greenhouses. Biol. Control 2003, 28, 313–319. [Google Scholar] [CrossRef]
- Aubry, O.; Cormier, D.; Chouinard, G.; Lucas, E. Influence of Plant, Animal and Mixed Resources on Development of the Zoophytophagous Plant Bug Campylomma Verbasci (Hemiptera: Miridae). Biocontrol Sci. Technol. 2015, 25, 1426–1442. [Google Scholar] [CrossRef]
- Leigh, T.F.; Jackson, C.E.; Wynholds, P.F.; Cota, J.A. Toxicity of Selected Insecticides Applied Topically to Lygus hesperus. J. Econ. Entomol. 1977, 70, 42–44. [Google Scholar] [CrossRef]
- Snodgrass, G.L. Insecticide Resistance in Field Populations of the Tarnished Plant Bug (Heteroptera: Miridae) in Cotton in the Mississippi Delta. J. Econ. Entomol. 1996, 89, 783–790. [Google Scholar] [CrossRef]
- Tillman, P.G.; Hammes, G.G.; Sacher, M.; Connair, M.; Brady, E.A.; Wing, K.D. Toxicity of a Formulation of the Insecticide Indoxacarb to the Tarnished Plant Bug, Lygus lineolaris (Hemiptera: Miridae), and the Big-Eyed Bug, Geocoris punctipes (Hemiptera: Lygaeidae). Pest Manag. Sci. Former. Pestic. Sci. 2002, 58, 92–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbahi, R.; Merzouki, A.; Guertin, C. Efficacy of Beauveria bassiana against the Strawberry Pests, Lygus lineolaris, Anthonomus Signatus and Otiorhynchus Ovatus. J. Appl. Entomol. 2008, 132, 151–160. [Google Scholar] [CrossRef]
- Sabbahi, R.; Merzouki, A.; Guertin, C. Efficacy of Beauveria bassiana (Bals.) Vuill. against the Tarnished Plant Bug, Lygus lineolaris L., in Strawberries. J. Appl. Entomol. 2008, 132, 124–134. [Google Scholar] [CrossRef]
- Sabbahi, R.; Lavallée, R.; Merzouki, A.; Guertin, C. Differentiation of Entomopathogenic Fungus Beauveria bassiana (Ascomycetes: Hypocreales) Isolates by PCR-RFLP. Phytoprotection 2009, 90, 49. [Google Scholar] [CrossRef] [Green Version]
- Portilla, M.; Snodgrass, G.L.; Luttrell, R. Effect of Morning and Night Application of Beauveria bassiana Strains NI8 and GHA against the Tarnished Plant Bug in Cotton. In Proceedings of the Beltwide Cotton Conferences. National Cotton Council, Memphis, TN, USA, 6–8 January 2014; pp. 729–734. [Google Scholar]
- Portilla, M.; Snodgrass, G.; Luttrell, R. Lethal and Sub-Lethal Effects of Beauveria bassiana (Cordycipitaceae) Strain NI8 on Chrysoperla Rufilabris (Neuroptera: Chrysopidae). Fla. Entomol. 2017, 100, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Catalán, T.P.; Wozniak, A.; Niemeyer, H.M.; Kalergis, A.M.; Bozinovic, F. Interplay between Thermal and Immune Ecology: Effect of Environmental Temperature on Insect Immune Response and Energetic Costs after an Immune Challenge. J. Insect Physiol. 2012, 58, 310–317. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Scott, W.P.; Smith, J.W. Host Plants and Seasonal Distribution of the Tarnished Plant Bug (Hemiptera: Miridae) in the Delta of Arkansas, Louisiana, and Mississippi. Environ. Entomol. 1984, 13, 110–116. [Google Scholar] [CrossRef]
- Chow, T.; Long, G.E.; Tamaki, G. Effects of Temperature and Hunger on the Functional Response of Geocoris bullatus (Say) (Hemiptera: Lygaeidae) to Lygus Spp. (Hemiptera: Miridae) Density. Environ. Entomol. 1983, 12, 1332–1338. [Google Scholar] [CrossRef]
- Foster, W.A.; Treherne, J.E. Evidence for the Dilution Effect in the Selfish Herd from Fish Predation on a Marine Insect. Nature 1981, 293, 466–467. [Google Scholar] [CrossRef]
- Turner, G.F.; Pitcher, T.J. Attack Abatement: A Model for Group Protection by Combined Avoidance and Dilution. Am. Nat. 1986, 128, 228–240. [Google Scholar] [CrossRef]
- Lucas, É.; Brodeur, J. A Fox in Sheep’s Clothing: Furtive Predators Benefit from the Communal Defense of Their Prey. Ecology 2001, 82, 3246–3250. [Google Scholar] [CrossRef]
- Dumont, F.; Lucas, E.; Brodeur, J. Do Furtive Predators Benefit from a Selfish Herd Effect by Living within Their Prey Colony? Behav. Ecol. Sociobiol. 2015, 69, 971–976. [Google Scholar] [CrossRef]
- Snodgrass, G.L.; Scott, W.P. Seasonal Changes in Pyrethroid Resistance in Tarnished Plant Bug (Heteroptera: Miridae) Populations during a Three-Year Period in the Delta Area of Arkansas, Louisiana, and Mississippi. J. Econ. Entomol. 2000, 93, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Snodgrass, G.L.; Chen, M.S. Enhanced Esterase Gene Expression and Activity in a Malathion-Resistant Strain of the Tarnished Plant Bug, Lygus lineolaris. Insect Biochem. Mol. Biol. 2004, 34, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, J.A.; Sparks, T.C.; Story, R.N. Evolution of Insect Resistance to Insecticides: A Reevaluation of the Roles of Physiology and Behavior. Bull. Esa 1984, 30, 41–51. [Google Scholar] [CrossRef]
- Kliot, A.; Ghanim, M. Fitness Costs Associated with Insecticide Resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef]
- Spurgeon, D.W. Efficacy of Beauveria bassiana against Lygus hesperus (Hemiptera: Miridae) at Low Temperatures. J. Entomol. Sci. 2010, 45, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Noma, T.; Strickler, K. Effects of Beauveria bassiana on Lygus hesperus (Hemiptera: Miridae) Feeding and Oviposition. Environ. Entomol. 2000, 29, 394–402. [Google Scholar] [CrossRef]
- Ugine, T.A. The Effect of Temperature and Exposure to Beauveria bassiana on Tarnished Plant Bug Lygus lineolaris (Heteroptera: Miridae) Population Dynamics, and the Broader Implications of Treating Insects with Entomopathogenic Fungi over a Range of Temperatures. Biol. Control 2011, 59, 373–383. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumont, F.; Provost, C. Using Autumnal Trap Crops to Manage Tarnished Plant Bugs (Lygus lineolaris). Insects 2022, 13, 441. https://doi.org/10.3390/insects13050441
Dumont F, Provost C. Using Autumnal Trap Crops to Manage Tarnished Plant Bugs (Lygus lineolaris). Insects. 2022; 13(5):441. https://doi.org/10.3390/insects13050441
Chicago/Turabian StyleDumont, François, and Caroline Provost. 2022. "Using Autumnal Trap Crops to Manage Tarnished Plant Bugs (Lygus lineolaris)" Insects 13, no. 5: 441. https://doi.org/10.3390/insects13050441
APA StyleDumont, F., & Provost, C. (2022). Using Autumnal Trap Crops to Manage Tarnished Plant Bugs (Lygus lineolaris). Insects, 13(5), 441. https://doi.org/10.3390/insects13050441