
Microscopic Identification of Anatomical Elements and Chemical Analysis of Secondary Nests of Vespa velutina nigrithorax du Buyson

 , ,
, ,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
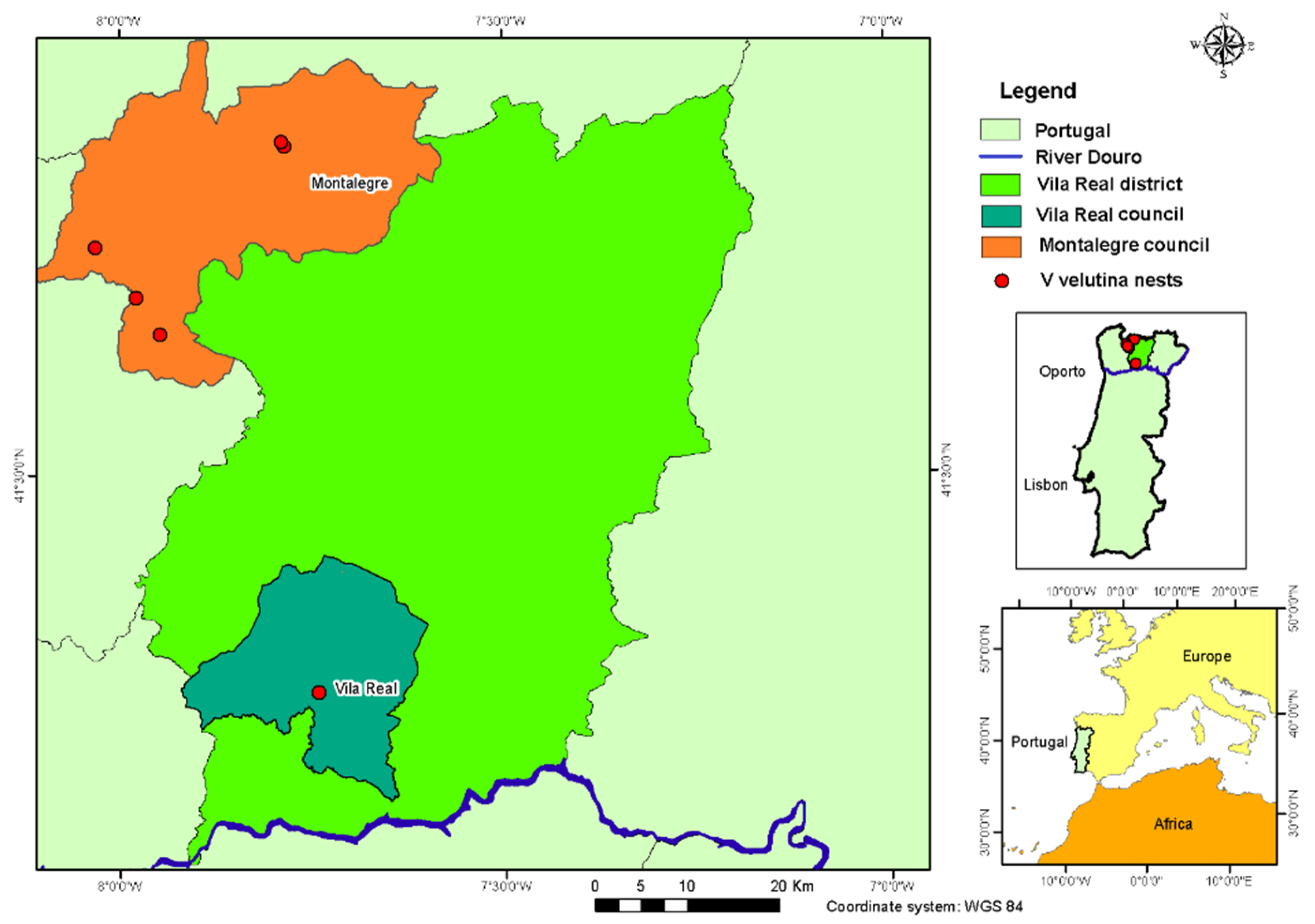
2.1. Location and Characterization of the Study Area
2.2. Nests Data
2.3. Microscopic Identification of Anatomical Elements
2.3.1. First stage: Intact Material (Non-Disaggregated Material) Analysis
2.3.2. Second Stage: Disaggregated Material in Distilled Water
2.3.3. Second Stage: Disaggregated Material by the Franklin Method
2.4. Chemical Analysis
2.4.1. X-ray Diffraction (XRD)
2.4.2. Scanning Electron Microscopy (SEM) Analysis with Energy Dispersive Spectroscopy (EDS)
2.5. Statistical Analysis
3. Results
3.1. First Stage: Intact Material (Non-Disaggregated Material)
3.2. Second Stage: Disaggregated Material Analysis
3.3. Chemical Analysis
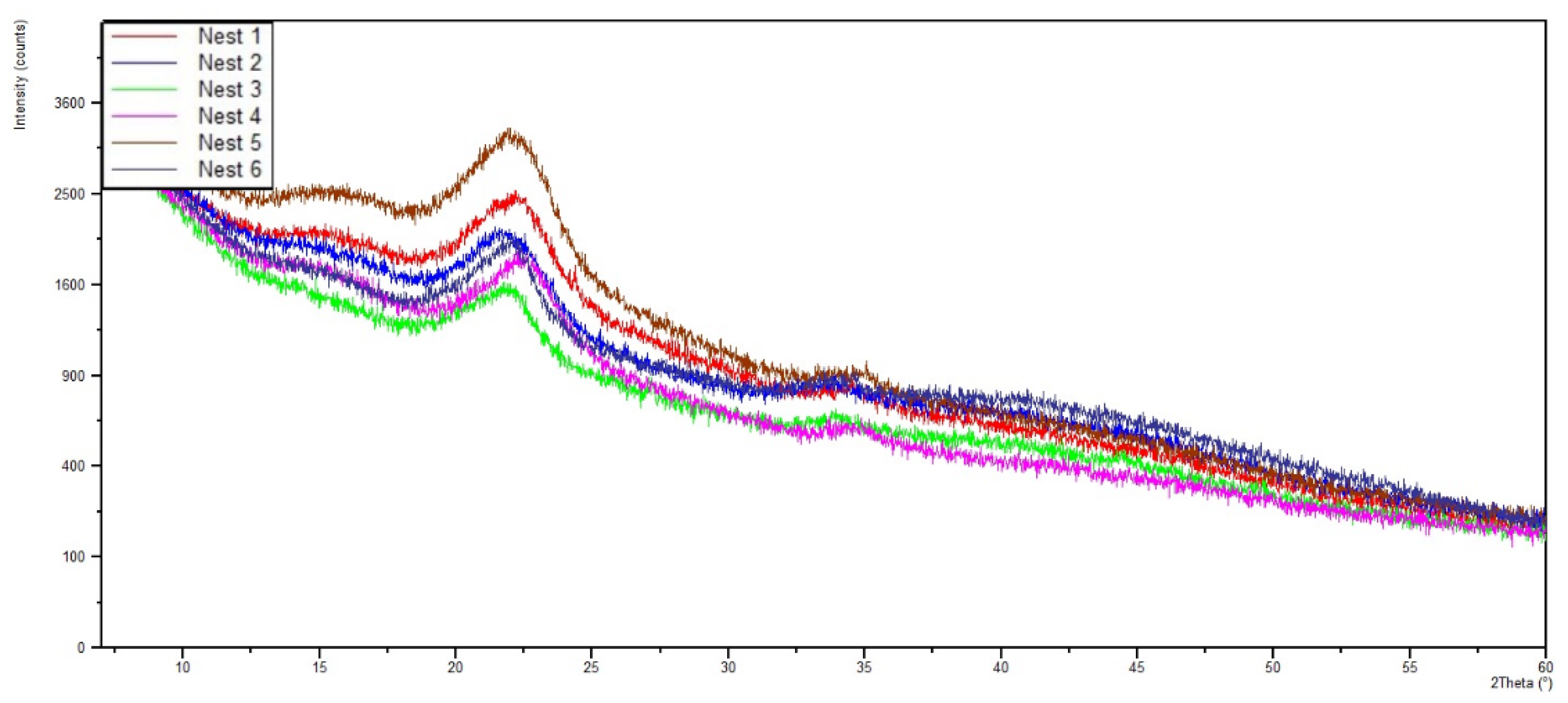
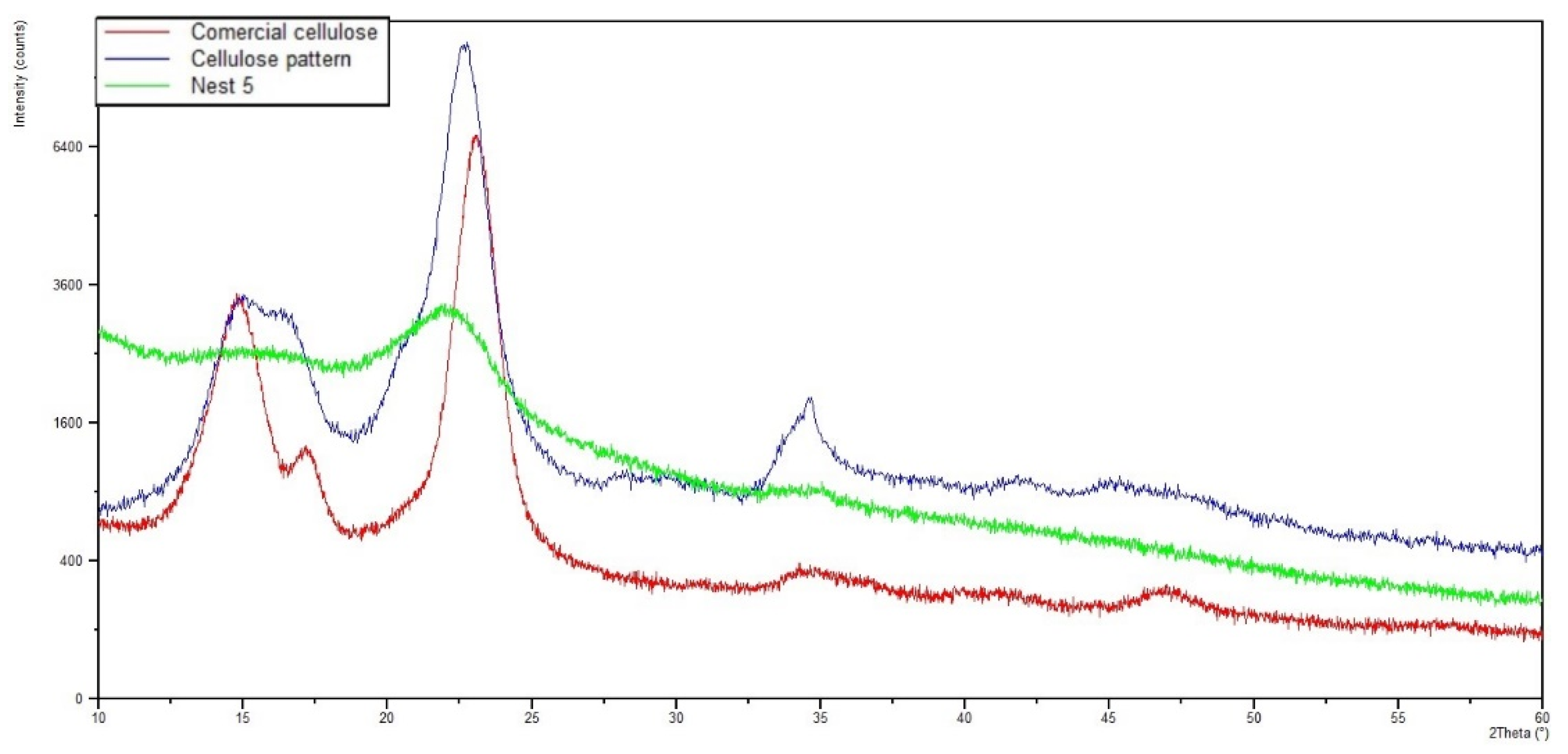
3.3.1. X-ray Diffraction (XRD)
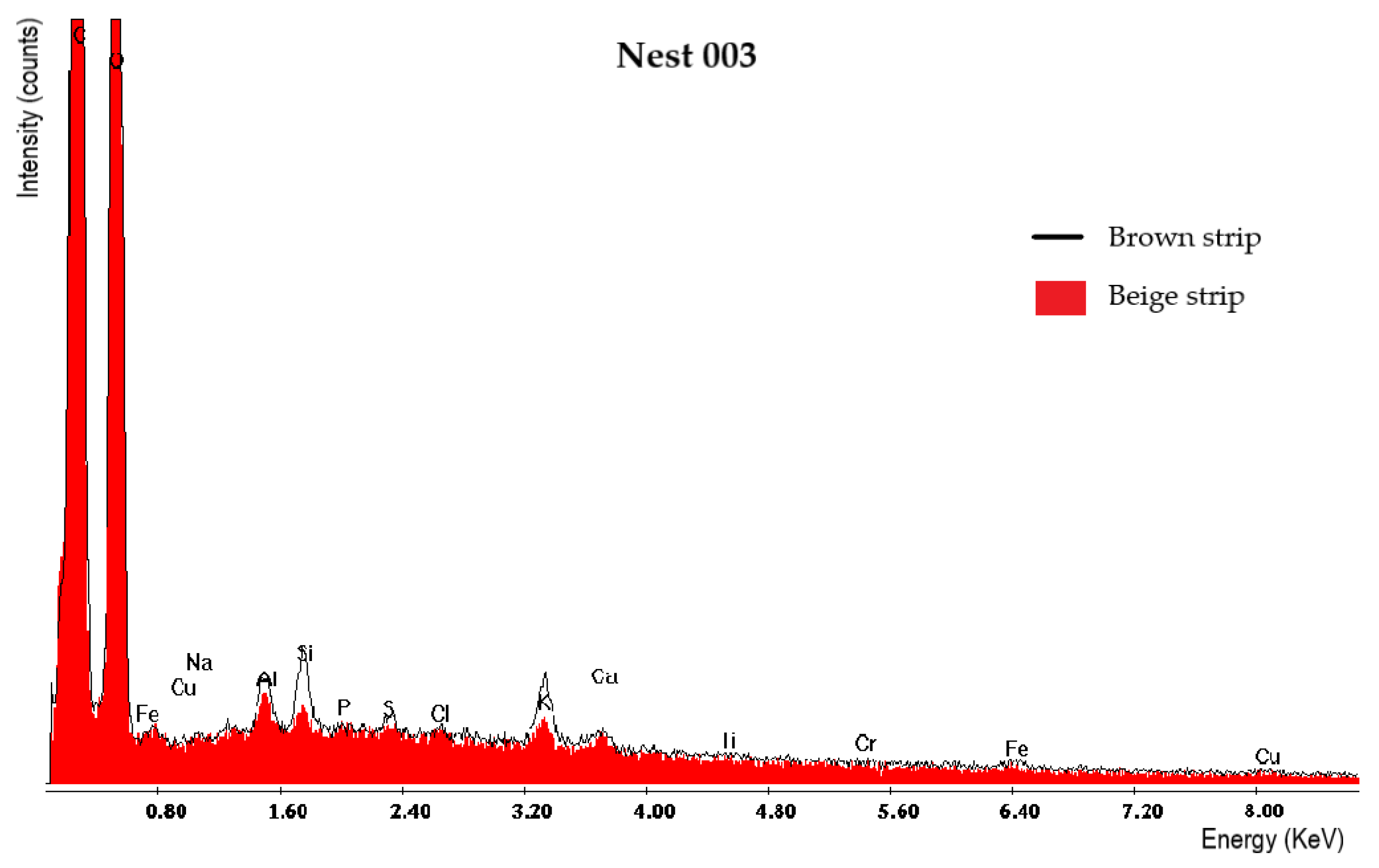
3.3.2. Scanning Electron Microscopy (SEM) Analysis with EDS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huber, J.T. Biodiversity of Hymenoptera. In Insect Biodiversity: Science and Society, 1st ed.; Foottit, R.G., Adler, P., Eds.; Wiley-Blackwell: Oxford, UK, 2009. [Google Scholar]
- Collins, P. The nests and nurseries of insects. Sci. Am. 1907, 97, 396–398. [Google Scholar] [CrossRef]
- Archer, M. Taxonomy, distribution and nesting biology of the Vespa bicolor group (Hym., Vespinae). Entomol. Mon. Mag. 1994, 130, 149–158. [Google Scholar]
- Monceau, K.; Bonnard, O.; Thiéry, D. Vespa velutina: A new invasive predator of honeybees in Europe. J. Pest Sci. 2014, 87, 1–16. [Google Scholar] [CrossRef]
- Villemant, C.; Haxaire, J.; Streito, J. La découverte du Frelon asiatique Vespa velutina, en France. Insectes 2006, 143, 3–7. [Google Scholar]
- López, S.; González, M.; Goldarazena, A. Vespa velutina Lepeletier, 1836 (Hymenoptera: Vespidae): First records in Iberian Peninsula. EPPO Bull. 2011, 41, 439–441. [Google Scholar] [CrossRef]
- Rome, Q.; Dambrine, L.; Onate, C.; Muller, F.; Villemant, C.; Garcia-Perez, L.; Maia, M.; Carvalho Esteves, P.; Bruneau, E. Spread of the invasive hornet Vespa velutina Lepeletier, 1836, in Europe in 2012 (Hym., Vespidae). Bull. Soc. Entomol. Fr. 2013, 118, 21–22. [Google Scholar] [CrossRef]
- Grosso-Silva, J.M.; Maia, M. Vespa velutina Lepeletier, 1836 (Hymenoptera, Vespidae), new species for Portugal. Arq. Entomol. 2012, 6, 53–54. [Google Scholar]
- Demichelis, S.; Manino, A.; Minuto, G.; Mariotti, M.; Porporato, M. Social wasp trapping in north west Italy: Comparison of different bait-traps and first detection of Vespa velutina. Bull. Insectol. 2014, 67, 307–317. [Google Scholar]
- Witt, R. Erstfund eines Nestes der Asiatischen Hornisse Vespa velutina Lepeletier, 1838 in Deutschland und Details zum Nestbau (Hymenoptera, Vespinae). Ampulex 2015, 7, 42–53. [Google Scholar]
- Leza, M.; Miranda, M.Á.; Colomar, V. First detection of Vespa velutina nigrithorax (Hymenoptera: Vespidae) in the Balearic Islands (Western Mediterranean): A challenging study case. Biol. Invasions 2018, 20, 1643–1649. [Google Scholar] [CrossRef]
- Budge, G.E.; Hodgetts, J.; Jones, E.P.; Ostojá-Starzewski, J.C.; Hall, J.; Tomkies, V.; Semmence, N.; Brown, M.; Wakefield, M.; Stainton, K. The invasion, provenance and diversity of Vespa velutina Lepeletier (Hymenoptera: Vespidae) in Great Britain. PLoS ONE 2017, 12, e0185172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ries, C.; Schneider, N.; Vitali, F.; Weigand, A. First records and distribution of the invasive alien hornet Vespa velutina nigrithorax du Buysson, 1905 (Hymenoptera: Vespidae) in Luxembourg. Bull. Soc. Nat. Luxemb. 2021, 123, 181–193. [Google Scholar]
- European Union. Regulation (EU) No 1143/2014 of the European Parliament and the Council of 22 October 2014 on the prevention and management of the introduction and spread of invasive alien species. Off. J. Eur. Union 2014, 317, 35–55. [Google Scholar]
- Carvalho, J.; Hipólito, D.; Santarém, F.; Martins, R.; Gomes, A.; Carmo, P.; Rodrigues, R.; Grosso-Silva, J.; Fonseca, C. Patterns of Vespa velutina invasion in Portugal using crowdsourced data. Insect Conserv. Divers. 2020, 13, 501–507. [Google Scholar] [CrossRef]
- Verdasca, M.J.; Rebelo, H.; Carvalheiro, L.G.; Rebelo, R. Invasive hornets on the road: Motorway-driven dispersal must be considered in management plans of Vespa velutina. NeoBiota 2021, 69, 177–198. [Google Scholar] [CrossRef]
- Requier, F.; Rome, Q.; Villemant, C.; Henry, M. A biodiversity-friendly method to mitigate the invasive Asian hornet’s impact on European honey bees. J. Pest Sci. 2019, 93, 1–9. [Google Scholar] [CrossRef]
- Rome, Q.; Perrard, A.; Muller, F.; Fontaine, C.; Quilès, A.; Zuccon, D.; Villemant, C. Not just honeybees: Predatory habits of Vespa velutina (Hymenoptera: Vespidae) in France. Ann. Soc. Entomol. Fr. 2021, 57, 1–11. [Google Scholar] [CrossRef]
- Laurino, D.; Lioy, S.; Carisio, L.; Manino, A.; Porporato, M. Vespa velutina: An alien driver of honey bee colony losses. Diversity 2020, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Perrard, A.; Haxaire, J.; Rortais, A.; Villemant, C. Observations on the colony activity of the Asian hornet Vespa velutina Lepeletier 1836 (Hymenoptera: Vespidae: Vespinae) in France. Ann. Soc. Entomol. Fr. 2009, 45, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Feás, X. Human fatalities caused by hornet, wasp and bee stings in spain: Epidemiology at state and sub-state level from 1999 to 2018. Biology 2021, 10, 73. [Google Scholar] [CrossRef]
- DGAV. Plano de Ação para a Vigilância e Controlo da Vespa velutina em Portugal; DGAV: Lisbon, Portugal, 2014; pp. 1–46.
- Lima, C.G.; Sofia Vaz, A.; Honrado, J.P.; Aranha, J.; Crespo, N.; Vicente, J.R. The invasion by the Yellow-legged hornet: A systematic review. J. Nat. Conserv. 2022, 67, 126173. [Google Scholar] [CrossRef]
- Matsuura, M. Biology of the Vespine Wasps; Springer Verlag: Berlin/Heidelberg, Germany, 1990; ISBN 3540519009. [Google Scholar]
- Matsuura, M. Vespa and Provespa. Soc. Biol. Wasps 1991, 232–262. [Google Scholar]
- Buysson, R. Du Monographie des guêpes ou vespa. Ann. Soc. Entomol. Fr. 1904, 527–528. [Google Scholar]
- Jeong, H.; Kim, J.M.; Kim, B.; Nam, J.O.; Hahn, D.; Choi, M.B. Nutritional Value of the Larvae of the Alien Invasive Wasp Vespa velutina nigrithorax and Amino Acid Composition of the Larval Saliva. Foods 2020, 9, 885. [Google Scholar] [CrossRef] [PubMed]
- Feás, X.; Vázquez-Tato, M.P.; Seijas, J.A.; Nikalje, A.P.G.; Fraga-López, F. Extraction and Physicochemical Characterization of Chitin Derived from the Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae). Molecules 2020, 25, 384. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Namin, S.M.; Meyer-Rochow, V.B.; Jung, C. Chemical composition and nutritional value of different species of Vespa hornets. Foods 2021, 10, 418. [Google Scholar] [CrossRef]
- Rome, Q.; Muller, F.J.; Touret-Alby, A.; Darrouzet, E.; Perrard, A.; Villemant, C. Caste differentiation and seasonal changes in Vespa velutina (Hym.: Vespidae) colonies in its introduced range. J. Appl. Entomol. 2015, 139, 771–782. [Google Scholar] [CrossRef]
- Pérez-De-Heredia, I.; Darrouzet, E.; Goldarazena, A.; Romón, P.; Iturrondobeitia, J.C. Differentiating between gynes and workers in the invasive hornet Vespa velutina (Hymenoptera, Vespidae) in Europe. J. Hymenopt. Res. 2017, 60, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-K.; Choi, M.; Moon, T.-Y. Occurrence of Vespa velutina Lepeletier from Korea, and a revised key for Korean Vespa species (Hymenoptera: Vespidae). Entomol. Res. 2006, 36, 112–115. [Google Scholar] [CrossRef]
- Kishi, S.; Goka, K. Review of the invasive yellow-legged hornet, Vespa velutina nigrithorax (Hymenoptera: Vespidae), in Japan and its possible chemical control. Appl. Entomol. Zool. 2017, 52, 361–368. [Google Scholar] [CrossRef]
- Lioy, S.; Laurino, D.; Capello, M.; Romano, A.; Manino, A.; Porporato, M. Effectiveness and selectiveness of traps and baits for catching the invasive hornet Vespa velutina. Insects 2020, 11, 706. [Google Scholar] [CrossRef] [PubMed]
- Perveen, F.; Shah, M. Nest Architectural Patterns by Three Wasp Species (Vespa velutina, Polistes flavus and Sceliphron formosum) with Reference to Their Behavior. Int. J. Insect Sci. 2013, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haouzi, M.; Gévar, J.; Khalil, A.; Darrouzet, E. Nest structures display specific hydrocarbon profiles: Insights into the chemical ecology of the invasive yellow-legged hornet Vespa velutina nigrithorax. Chemoecology 2021, 31, 227–238. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Archer, M.E. Taxonomy, distribution and nesting biology of species of the genera Provespa Ashmead and Vespa Linnaeus (Hymenoptera, Vespidae). Entomol. Mon. Mag. 2008, 144, 69. [Google Scholar]
- Lioy, S.; Manino, A.; Porporato, M.; Laurino, D.; Romano, A.; Capello, M.; Bertolino, S. Establishing surveillance areas for tackling the invasion of Vespa velutina in outbreaks and over the border of its expanding range. NeoBiota 2019, 69, 51–69. [Google Scholar] [CrossRef]
- Choi, M.B.; Martin, S.J.; Lee, J.W. Distribution, spread, and impact of the invasive hornet Vespa velutina in South Korea. J. Asia Pac. Entomol. 2012, 15, 473–477. [Google Scholar] [CrossRef]
- Tsoumis, G. Science and Technology of Wood—Structure, Properties, Utilization; Van Nostrand Reinhold: New York, NY, USA, 1991; p. 494. [Google Scholar]
- Haygreen, J.G.; Bowyer, J.L. Forest Products and Wood Science: An Introduction; Iowa State University Press: Ames, IA, USA, 1982; ISBN 9780813818009. [Google Scholar]
- Monceau, K.; Tourat, A.; Arca, M.; Bonnard, O.; Arnold, G.; Thiéry, D. Daily and Seasonal Extranidal Behaviour Variations in the Invasive Yellow-Legged Hornet, Vespa velutina Lepeletier (Hymenoptera: Vespidae). J. Insect Behav. 2017, 30, 220–230. [Google Scholar] [CrossRef]
- Feás Sánchez, X.; Charles, R.J. Notes on the Nest Architecture and Colony Composition in Winter of the Yellow-Legged Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), in Its Introduced Habitat in Galicia (NW Spain). Insects 2019, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Kudô, K.; Yamane, S.; Mateus, S.; Tsuchida, K.; Itô, Y.; Miyano, S.; Yamamoto, H.; Zucchi, R. Nest materials and some chemical characteristics of nests of a New World swarm-founding polistine wasp, Polybia paulista (Hymenoptera Vespidae). Ethol. Ecol. Evol. 2010, 20, 351–360. [Google Scholar] [CrossRef]
- Waldhoff, D.P.P. Morphology and Anatomy of Leaves; Springer: Dordrecht, The Netherlands, 2010; Volume 210, ISBN 9789048187256. [Google Scholar]
- Kushwaha, U.; Shrivastava, R.; Mishra, A. Dust Pollution Effects on the Leaves Anatomy of Catharanthus roseus and Nerium oleander Growing Along the Road Side of Rewa City. Int. J. Eng. Sci. 2018, 7, 1–7. [Google Scholar]
- Martel, C.; Rojas, N.; Marín, M.; Avilés, R.; Neira, E.; Santiago, J. Caesalpinia spinosa (Caesalpiniaceae) leaves: Anatomy, histochemistry, and secondary metabolites. Braz. J. Bot. 2014, 37, 167–174. [Google Scholar] [CrossRef]
- Pettersen, R.C. The chemical composition of wood. Chem. Solid Wood 1984, 207, 57–126. [Google Scholar]
- Sjöström, E. Wood Chemistry: Fundamentals and Applications; Academic Press, Inc.: San Diego, CA, USA, 1993; ISBN 0-1264-7481-8. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nest Number | Nest Place | LgtN | CircN | WthN | NumbNC |
|---|---|---|---|---|---|
| 001 | Tree | 21.0 | 60.3 | 19.0 | 4.0 |
| 002 | Indoor | 19.2 | 52.0 | 18.7 | 5.0 |
| 003 | Tree | 41.2 | 58.0 | 39.0 | 5.0 |
| 004 | Tree | 46.0 | 110.0 | 37.5 | 7.0 |
| 005 | Tree | 47.7 | 95.5 | 33.0 | 7.0 |
| 006 | Tree | 65.0 | 134.0 | 45.0 | 9.0 |
| Nest Number | Date Removed Nest | Municipality | Nest Place | Tree’s Species Support | HN (m) | Long | Lat | Altitude (m) |
|---|---|---|---|---|---|---|---|---|
| 001 | 13/09/2019 | Montalegre | Tree | Pseudotsuga menziesii | 20 | −7.785705 | 41.822405 | 1050 |
| 002 | 13/09/2019 | Montalegre | Indoor | 4 | −7.789125 | 41.826872 | 1000 | |
| 003 | 20/09/2019 | Vila Real | Tree | Pinus pinaster | 24 | −7.741917 | 41.287344 | 500 |
| 004 | 04/11/2019 | Montalegre | Tree | Fraxinus angustifolia | 17 | −8.032647 | 41.723639 | 350 |
| 005 | 06/11/2019 | Montalegre | Tree | Quercus robur | 15 | −7.948567 | 41.638379 | 850 |
| 006 | 13/12/2019 | Montalegre | Tree | Quercus robur | 18 | −7.981200 | 41.674153 | 700 |
| Elements | Average | Tukey Test | |||
|---|---|---|---|---|---|
| All ± sd | Beige | Brown | Sig. | p-Value | |
| C | 63.01 ± 1.31 | 62.41 | 63.61 | ns | 0.1165 |
| O | 36.35 ± 1.36 | 37.00 | 35.70 | ns | 0.0977 |
| Na | 0.08 ± 0.02 | 0.08 | 0.08 | ns | 0.7961 |
| Al | 0.19 ± 0.07 | 0.22 | 0.17 | ns | 0.2094 |
| Si | 0.13 ± 0.07 | 0.07 | 0.19 | *** | <0.0001 |
| P | 0.04 ± 0.02 | 0.04 | 0.04 | ns | 0.7618 |
| S | 0.04 ± 0.02 | 0.04 | 0.05 | ns | 0.0588 |
| Cl | 0.01 ± 0.01 | 0.02 | 0.01 | ns | 0.0959 |
| K | 0.05 ± 0.02 | 0.04 | 0.05 | ns | 0.3913 |
| Ca | 0.04 ± 0.02 | 0.04 | 0.05 | ns | 0.1256 |
| Ti | 0.01 ± 0.00 | 0.01 | 0.01 | ns | 1.0000 |
| Cr | 0.01 ± 0.01 | 0.01 | 0.00 | ns | 0.0734 |
| Fe | 0.02 ± 0.01 | 0.02 | 0.02 | ns | 0.5701 |
| Cu | 0.02 ± 0.01 | 0.02 | 0.02 | ns | 0.3628 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crespo, N.; Louzada, J.; Fernandes, L.S.; Tavares, P.B.; Aranha, J. Microscopic Identification of Anatomical Elements and Chemical Analysis of Secondary Nests of Vespa velutina nigrithorax du Buyson. Insects 2022, 13, 537. https://doi.org/10.3390/insects13060537
Crespo N, Louzada J, Fernandes LS, Tavares PB, Aranha J. Microscopic Identification of Anatomical Elements and Chemical Analysis of Secondary Nests of Vespa velutina nigrithorax du Buyson. Insects. 2022; 13(6):537. https://doi.org/10.3390/insects13060537
Chicago/Turabian StyleCrespo, Nazaret, José Louzada, Lisete S. Fernandes, Pedro B. Tavares, and José Aranha. 2022. "Microscopic Identification of Anatomical Elements and Chemical Analysis of Secondary Nests of Vespa velutina nigrithorax du Buyson" Insects 13, no. 6: 537. https://doi.org/10.3390/insects13060537
APA StyleCrespo, N., Louzada, J., Fernandes, L. S., Tavares, P. B., & Aranha, J. (2022). Microscopic Identification of Anatomical Elements and Chemical Analysis of Secondary Nests of Vespa velutina nigrithorax du Buyson. Insects, 13(6), 537. https://doi.org/10.3390/insects13060537