
Positive Interactions between Aceria pallida and Bactericera gobica on Goji Berry Plants



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
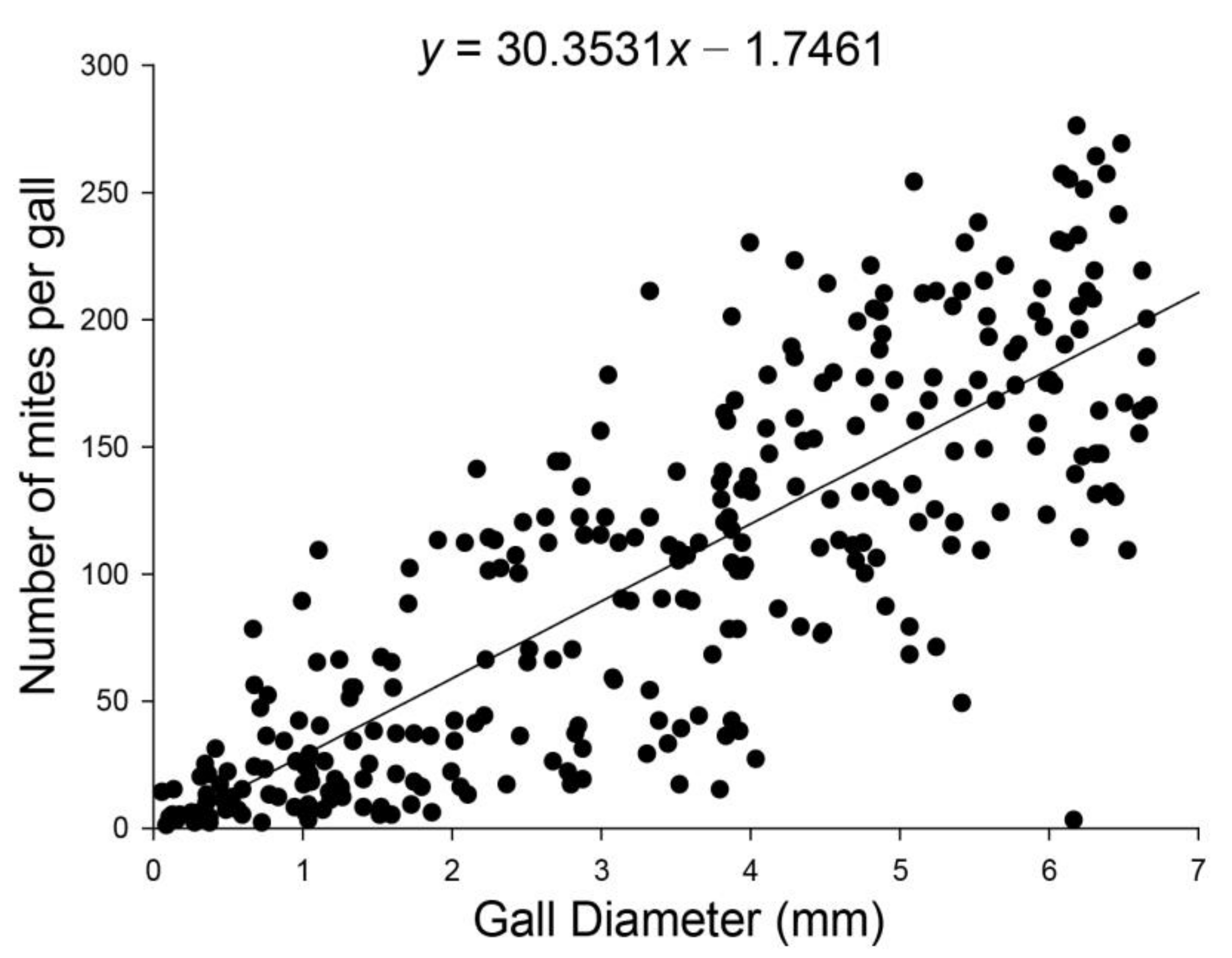
2.2. Relationship between Gall Diameter and A. pallida Abundance in The Gall
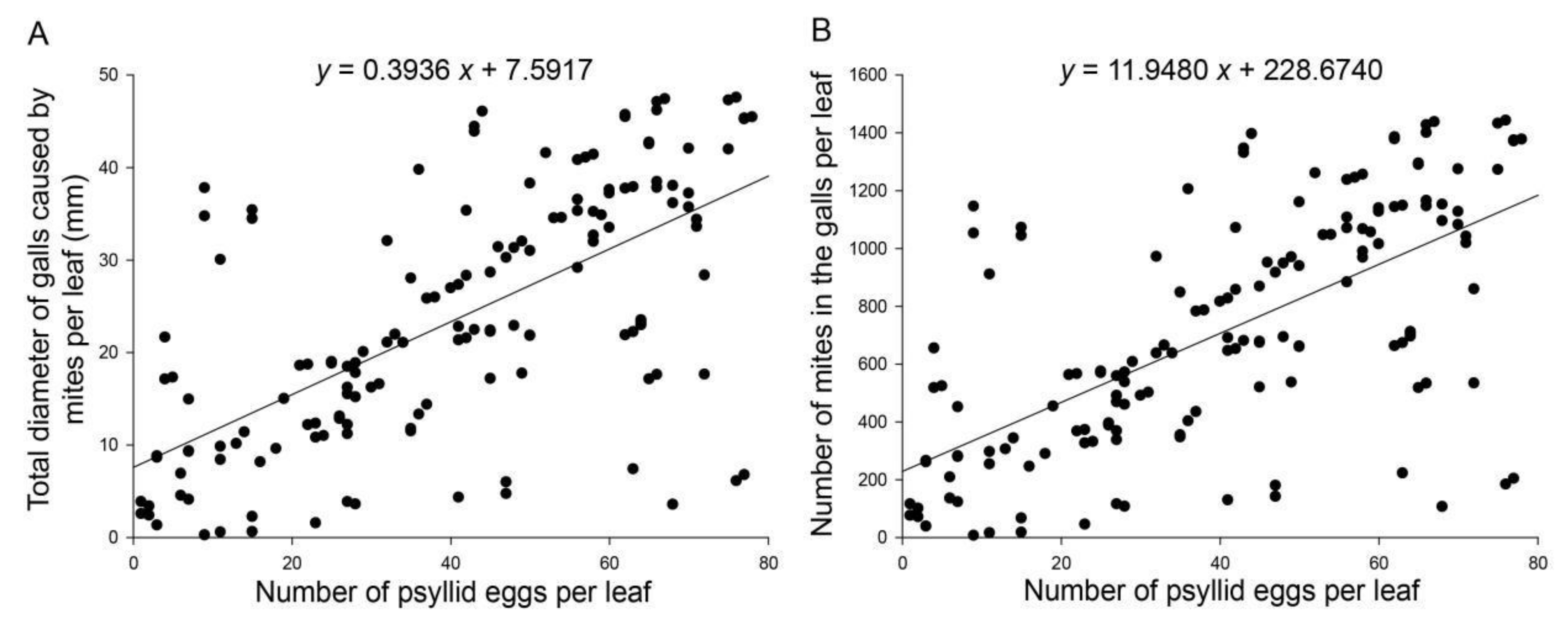
2.3. Relationship between Abundances of A. pallida and B. gobica Eggs on Goji Berry Leaves
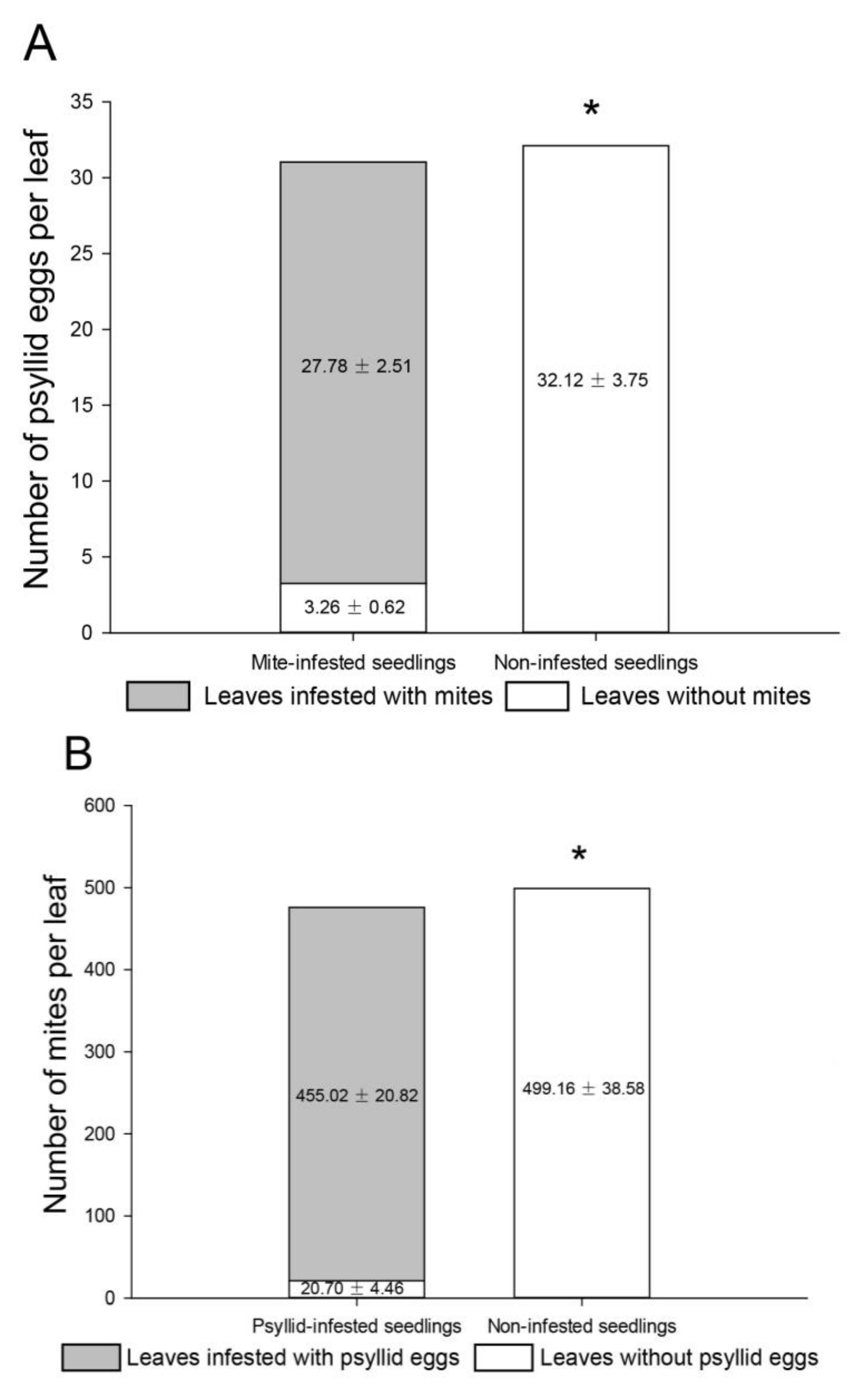
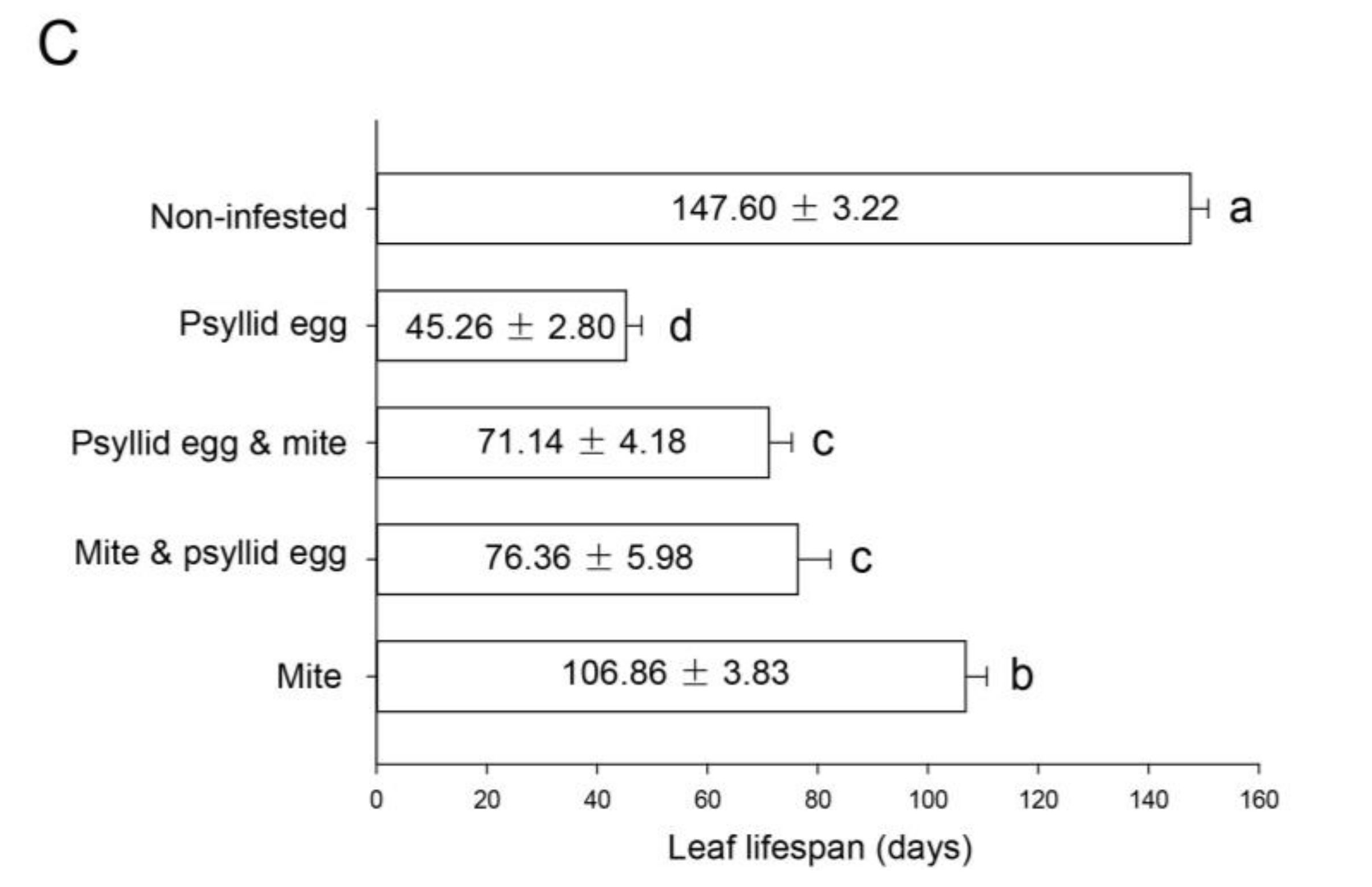
2.4. Habitat Selection Preferences for A. pallida and B. gobica
2.5. Statistical Analysis
3. Results
3.1. Relationship between Diameter of Gall and A. pallida Abundance in the Gall
3.2. Relationship between Abundances of A. pallida and B. gobica Eggs on Goji Berry Leaves
3.3. Habitat Selection Preferences for A. pallida and B. gobica
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glas, J.J.; Alba, J.M.; Simoni, S.; Villarroel, C.A.; Stoops, M.; Schimmel, B.C.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Defense suppression benefits herbivores that have a monopoly on their feeding site but can backfire within natural communities. BMC Biol. 2014, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Kiełkiewicz, M.; Barczak-Brzyżek, A.; Karpińska, B.; Filipecki, M. Unravelling the Complexity of Plant Defense Induced by a Simultaneous and Sequential Mite and Aphid Infestation. Int. J. Mol. Sci. 2019, 20, 806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindquist, E.E. External anatomy and notation of structures. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, pp. 3–32. [Google Scholar]
- Sabelis, M.W.; Bruin, J. Evolutionary ecology: Life history patterns, food plant choice and dispersa. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, pp. 329–366. [Google Scholar]
- Chong, J.H. A witch’s brew of troubles with the bermudagrass mite. USGA Green. Sect. Rec. 2013, 51, 4p. [Google Scholar]
- Michalska, K.; Skoracka, A.; Navia, D.; Amrine, J.W. Behavioural studies on eriophyoid mites: An overview. Exp. Appl. Acarol. 2010, 51, 31–59. [Google Scholar] [CrossRef] [Green Version]
- Hofstetter, R.W.; Moser, J.C. The role of mites in insect-fungus associations. Annu. Rev. Entomol. 2014, 59, 537–557. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.L.; Chen, Q.F.; Guo, T.; Wang, X.R.; Li, P.; Wang, X.W.; Liu, S.S. Differential efficiency of a begomovirus to cross the midgut of different species of whiteflies results in variation of virus transmission by the vectors. Sci. China Life Sci. 2018, 61, 1254–1265. [Google Scholar] [CrossRef]
- Perotti, M.A.; Braig, H.R. Phoretic mites associated with animal and human decomposition. Exp. Appl. Acarol. 2009, 49, 85–124. [Google Scholar] [CrossRef]
- Houck, M.A.; OConnor, B.M. Ecological and evolutionary significance of phoresy in the Astigmata. Annu. Rev. Entmol. 1991, 36, 611–636. [Google Scholar] [CrossRef]
- White, P.S.; Morran, L.; de Roode, J. Phoresy. Curr. Biol. 2017, 27, 578–580. [Google Scholar] [CrossRef]
- Shi, P.J.; Hui, C.; Men, X.Y.; Zhao, Z.H.; Ouyang, F.; Ge, F.; Jin, X.S.; Cao, H.F.; Li, B.L. Cascade effects of crop species richness on the diversity of pest insects and their natural enemies. Sci. China Life Sci. 2014, 57, 718–725. [Google Scholar] [CrossRef] [Green Version]
- Pfammatter, J.A.; Rafa, K.F. Do phoretic mites influence the reproductive success of Ips grandicollis (Coleoptera: Curculionidae)? Environ. Entomol. 2015, 44, 1498–1511. [Google Scholar] [CrossRef]
- Wu, P.X.; Xu, J.; Zhang, R.; He, J.; Zhang, R.Z. New found of overwintering gall mites carried by adult Poratrioza sinica. Chin. J. Appl. Ecol. 2016, 53, 417–419. [Google Scholar]
- Wang, K.; Xiao, J.; Peng, B.; Xing, F.Y.; So, K.F.; Tipoe, G.; Lin, B. Retinal structure and function preservation by polysaccharides of wolfberry in a mouse model of retinal degeneration. Sci. Rep. 2014, 4, 7601. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.X.; Ma, B.X.; Yan, S.; Xu, J.; Zhang, R.Z. The hyperparasitoid Marietta picta (Hymenoptera: Aphelinidae) mediates competitive interactions between two parasitoids of Paratrioza sinica (Hemiptera: Psyllidae): Tamarixia lyciumi (Hymenoptera: Eulophidae) and Psyllaephagus arenarius (Hymenoptera: Encyrtidae). Biol. Control 2018, 126, 169–176. [Google Scholar]
- Wu, P.X.; Ma, B.X.; Wu, F.M.; Xu, J.; Zhang, R.Z. The endoparasitoid Psyllaephagus arenarius benefits from the ectoparasitic venom via multiparasitism with the ectoparasitoid Tamarixia lyciumi. Insect Sci. 2020, 27, 815–825. [Google Scholar] [CrossRef]
- Wu, P.X.; Ma, B.X.; Wu, F.M.; Ouyang, H.Y.; Fan, J.Y.; Xu, J.; Zhang, R.Z. The hyperparasitoid Marietta picta mediates the coexistence of primary parasitoids of goji berry psyllid. Entomol. Gen. 2020, 2, 187–194. [Google Scholar] [CrossRef]
- Liu, S. Studies on the Phoresy of Aceria pallida. Ph.D. Dissertation, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, China, 2016. [Google Scholar]
- Westphal, E.; Bronner, R.; Ret, M.L. Changes in leaves of susceptible and resistant Solanum dulcamara infested by the gall mite Eriophyes cladophthirus (Acarina, Eriophyoidea). Can. J. Bot. 1981, 59, 875–882. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K. The adaptive signifcance of insect gall morphology. Trends Ecol. Evol. 2003, 18, 512–522. [Google Scholar] [CrossRef]
- Tooker, J.F.; Rohr, J.R.; Abrahamson, W.G.; De Moraes, C.M. Gall insects can avoid and alter indirect plant defenses. New Phytol. 2008, 178, 657–671. [Google Scholar] [CrossRef]
- Westphal, E.; Perrot-Minnot, M.J.; Kreiter, S.; Gutierrez, J. Hypersensitive reaction of Solanum dulcamara to the gall mite Aceria cladophthirus causes an increased susceptibility to Tetranychus urticae. Exp. Appl. Acarol. 1992, 15, 15–26. [Google Scholar] [CrossRef]
- Prior, K.M.; Hellmann, J.J. Impact of an invasive oak gall wasp on a native butterfly: A test of plant-mediated competition. Ecology 2010, 91, 3284–3293. [Google Scholar] [CrossRef]
- Savage, A.; Peterson, M. Mutualism in a community context: The positive feedback between an ant–aphid mutualism and a gallmaking midge. Oecologia 2007, 151, 280–291. [Google Scholar] [CrossRef]
- Wu, P.X.; Ma, B.X.; Shuo, Y.; Xu, J.; He, J.; Zhang, R.; Zhang, R.Z. Protective effects of the egg stalk of Paratrioza sinica (Hemiptera: Psyllidae) at various angles and spacings against three predaceous coccinellids, Harmonia axyridis, Coccinella septempunctata and Hippodamia variegate (Coleoptera: Coccinellidae). Pest Manag. Sci. 2018, 74, 356–365. [Google Scholar] [CrossRef]
- Deshpande, V.G. Miscellaneuos observations on the biology of Aleurodidae (Aleyrodes brassica). J. Bombay Nat. Hist. Soc. 1936, 39, 190–193. [Google Scholar]
- Paulson, G.S.; Beardsley, J.W. Whitefly (Hemiptera: Aleyrodidae) egg pedicel insertion into plant stomata. Ann. Entomol. Soc. Am. 1985, 78, 506–508. [Google Scholar] [CrossRef]
- Buckner, J.S.; Freeman, T.P.; Ruud, R.L.; Chu, C.C.; Henneberry, T.J. Characterization and functions of the whitefly egg pedicel. Arch. Insect Biochem. Physiol. 2002, 49, 22–33. [Google Scholar] [CrossRef]
- Brown, M.S.; Blubaugh, C.K.; Chong, J.H. Biology and management of eriophyid mites in turfgrass. J. Integr. Pest Manag. 2021, 12, 25. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Burns, A.E.; Hall, A.; Riegler, M.; Taylor, G.S. Nutritional enhancement of leaves by a psyllid through senescence-like processes: Insect manipulation or plant defence? Oecologia 2014, 176, 1061–1074. [Google Scholar] [CrossRef] [Green Version]
- Mapes, C.C.; Davies, P.J. Indole-3-acetic acid and ball gall development on Solidago altissima. New Phytol. 2001, 151, 195–202. [Google Scholar] [CrossRef]
- De Lillo, E.; Monfreda, R. ‘Salivary secretions’ of eriophyoids (Acari: Eriophyoidea): First results of an experimental model. Exp. Appl. Acarol. 2004, 34, 291–306. [Google Scholar]
- Gan, S.; Amasino, R.M. Inhibition of leaf senescence by autoregulated production of cytokinin. Science 1995, 270, 1986–1988. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant. Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, E.E.; Oldfield, G.N. Evolution of eriophyoid mites in relation to their host plants. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, pp. 277–300. [Google Scholar]
- Morath, S.U.; Pratt, P.D.; Silvers, C.S.; Center, T.D. Herbivory by Boreioglycaspis melaleucae (Hemiptera: Psyllidae) accelerates foliar senescence and abscission in the invasive tree Melaleuca quinquenervia. Environ. Entomol. 2006, 35, 1372–1378. [Google Scholar] [CrossRef]
- Casteel, C.L.; Hansen, A.K.; Walling, L.L.; Paine, T.D. Correction: Manipulation of plant defense responses by the tomato psyllid (Bactericerca cockerelli) and its associated endosymbiont Candidatus Liberibacter psyllaurous. PLoS ONE 2021, 7, e35191. [Google Scholar] [CrossRef]
- Kaplan, I.; Denno, R.F. Interspecifc interactions in phytophagous insects revisited: A quantitative assessment of competition theory. Ecol. Lett. 2007, 10, 977–994. [Google Scholar] [CrossRef] [PubMed]
- Petanović, R.; Kielkiewicz, M. Plant–eriophyoid mite interactions: Cellular biochemistry and metabolic responses induced in mite-injured plants. Part I. Exp. Appl. Acarol. 2010, 51, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Hrudová, E.; Šafránková, I. Goji berry gall mite Aceria kuko occurrence in the Czech Republic—short communication. Plant Protect. Sci. 2018, 54, 39–42. [Google Scholar]
- Wu, P.X.; Head, M.L.; Liu, C.; Haseeb, M.; Zhang, R.Z. The high invasion success of fall armyworm is related to life-history strategies across a range of stressful temperatures. Pest Manag. Sci. 2022, 78, 2398–2404. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.; Ge, Y.; He, J.; Haseeb, M.; Zhang, R. Positive Interactions between Aceria pallida and Bactericera gobica on Goji Berry Plants. Insects 2022, 13, 577. https://doi.org/10.3390/insects13070577
Wu P, Ge Y, He J, Haseeb M, Zhang R. Positive Interactions between Aceria pallida and Bactericera gobica on Goji Berry Plants. Insects. 2022; 13(7):577. https://doi.org/10.3390/insects13070577
Chicago/Turabian StyleWu, Pengxiang, Yang Ge, Jia He, Muhammad Haseeb, and Runzhi Zhang. 2022. "Positive Interactions between Aceria pallida and Bactericera gobica on Goji Berry Plants" Insects 13, no. 7: 577. https://doi.org/10.3390/insects13070577
APA StyleWu, P., Ge, Y., He, J., Haseeb, M., & Zhang, R. (2022). Positive Interactions between Aceria pallida and Bactericera gobica on Goji Berry Plants. Insects, 13(7), 577. https://doi.org/10.3390/insects13070577