
The Impacts of Climate Change on the Potential Distribution of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. CLIMEX Model
2.2. Climate Data
2.3. Collection of P. interpunctella Distribution Data in China
2.4. Fitting CLIMEX Parameters
2.4.1. Temperature Index (TI) and Length of the Growing Season (PDD)
2.4.2. Moisture Index (MI)
2.4.3. Stress Index (SI)
3. Results
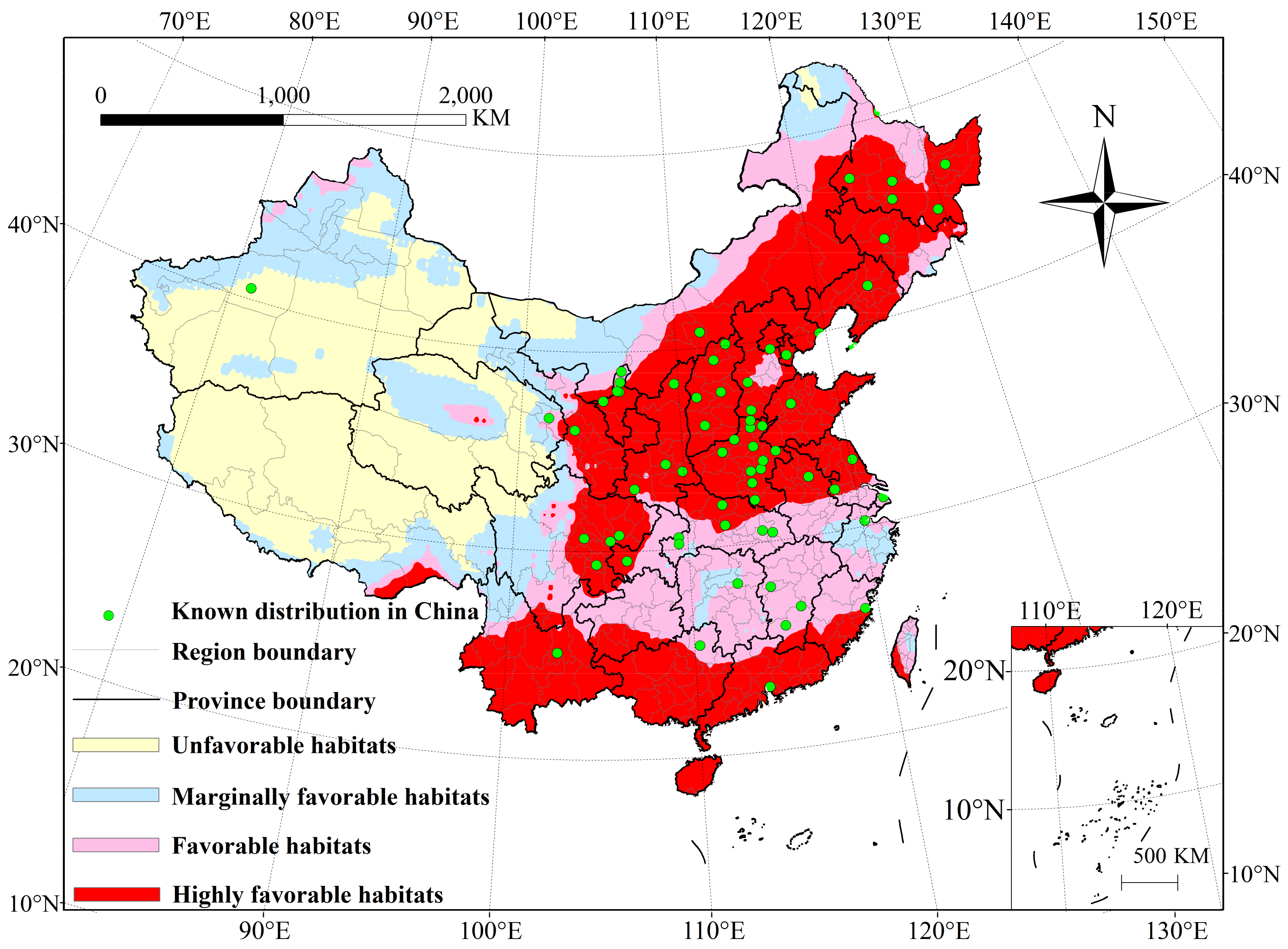
3.1. Potential Distribution of P. interpunctella under Historical Climate Conditions
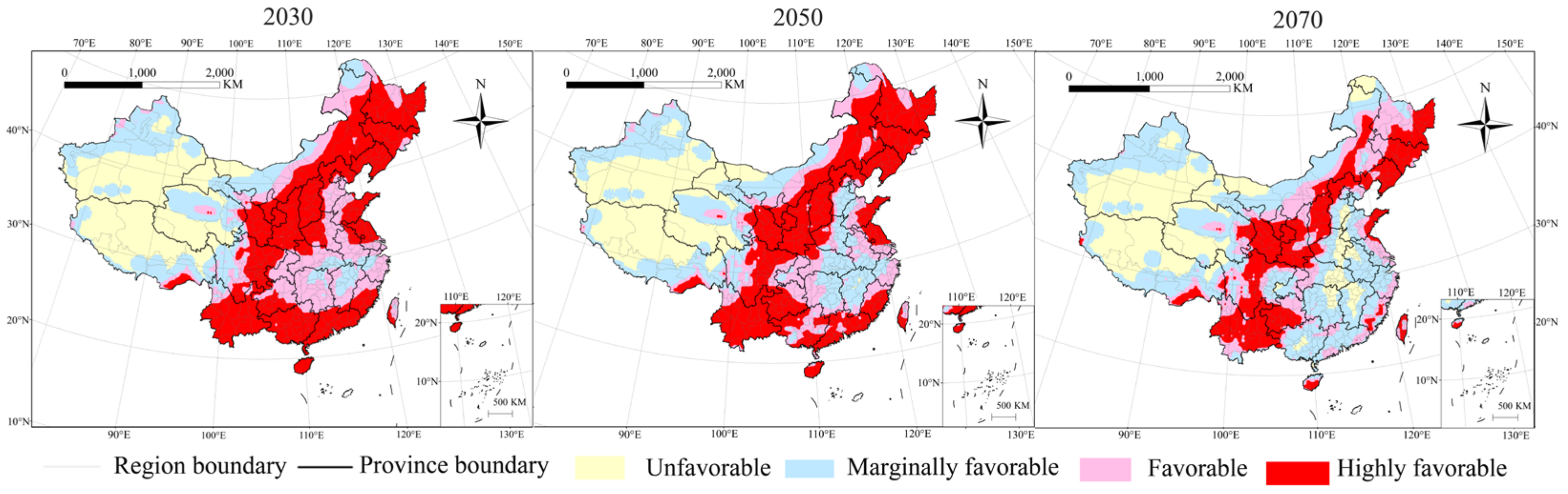
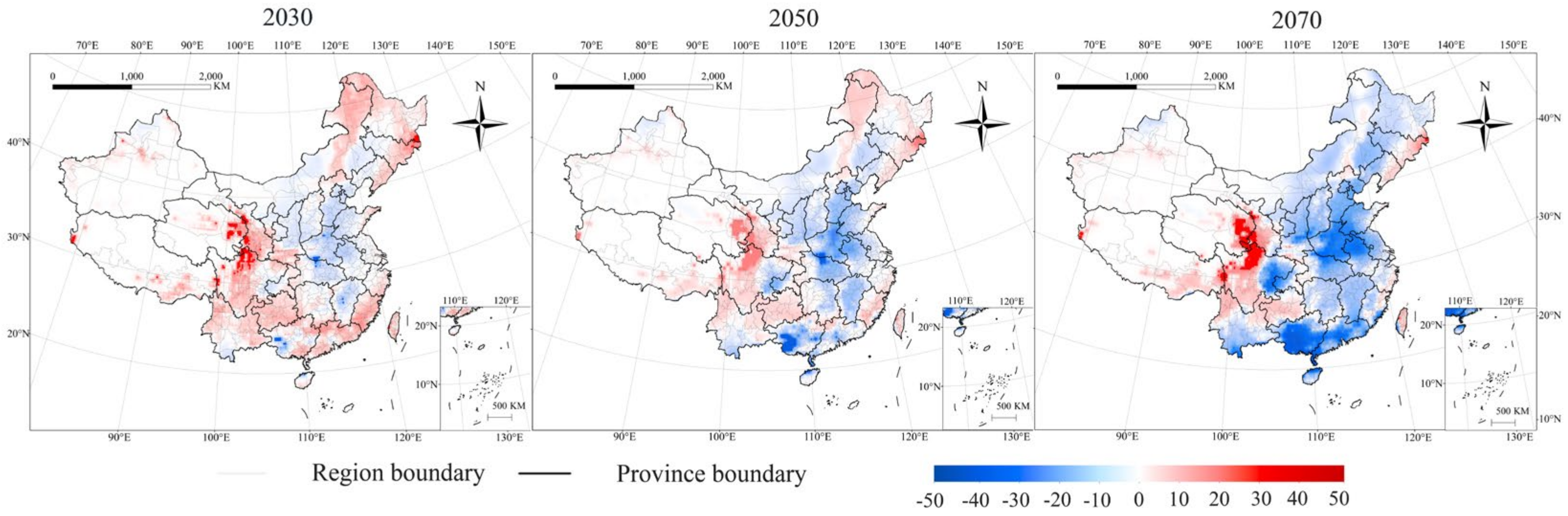
3.2. Potential Distribution of P. interpunctella in Future Climate
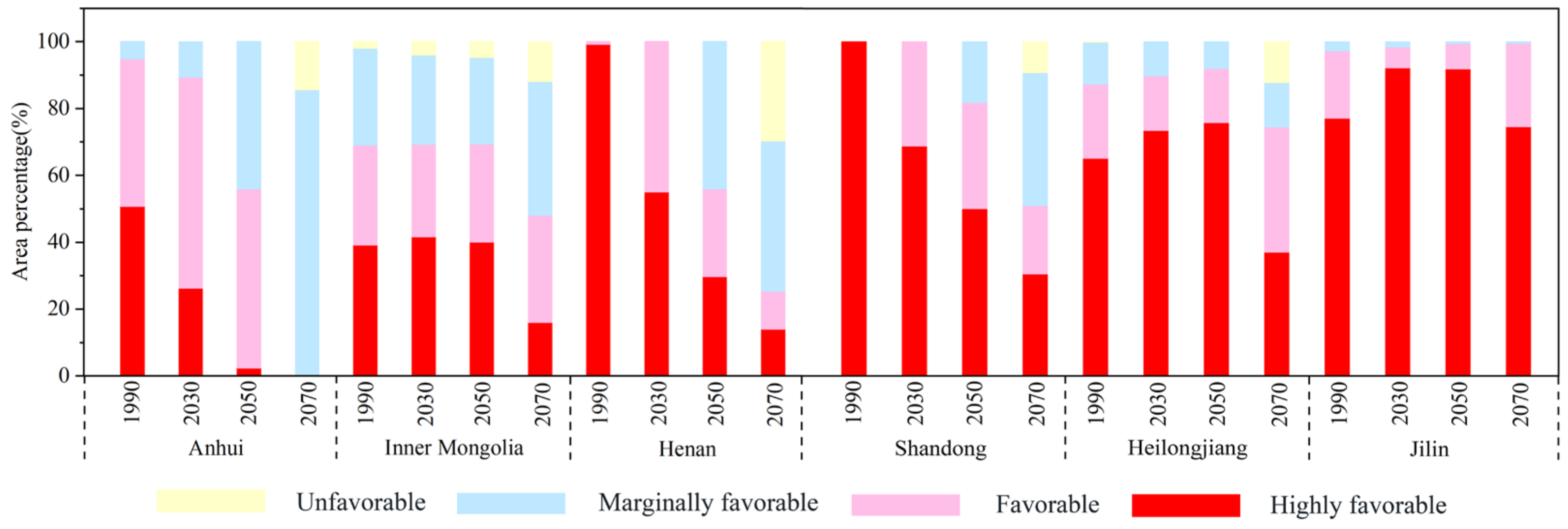
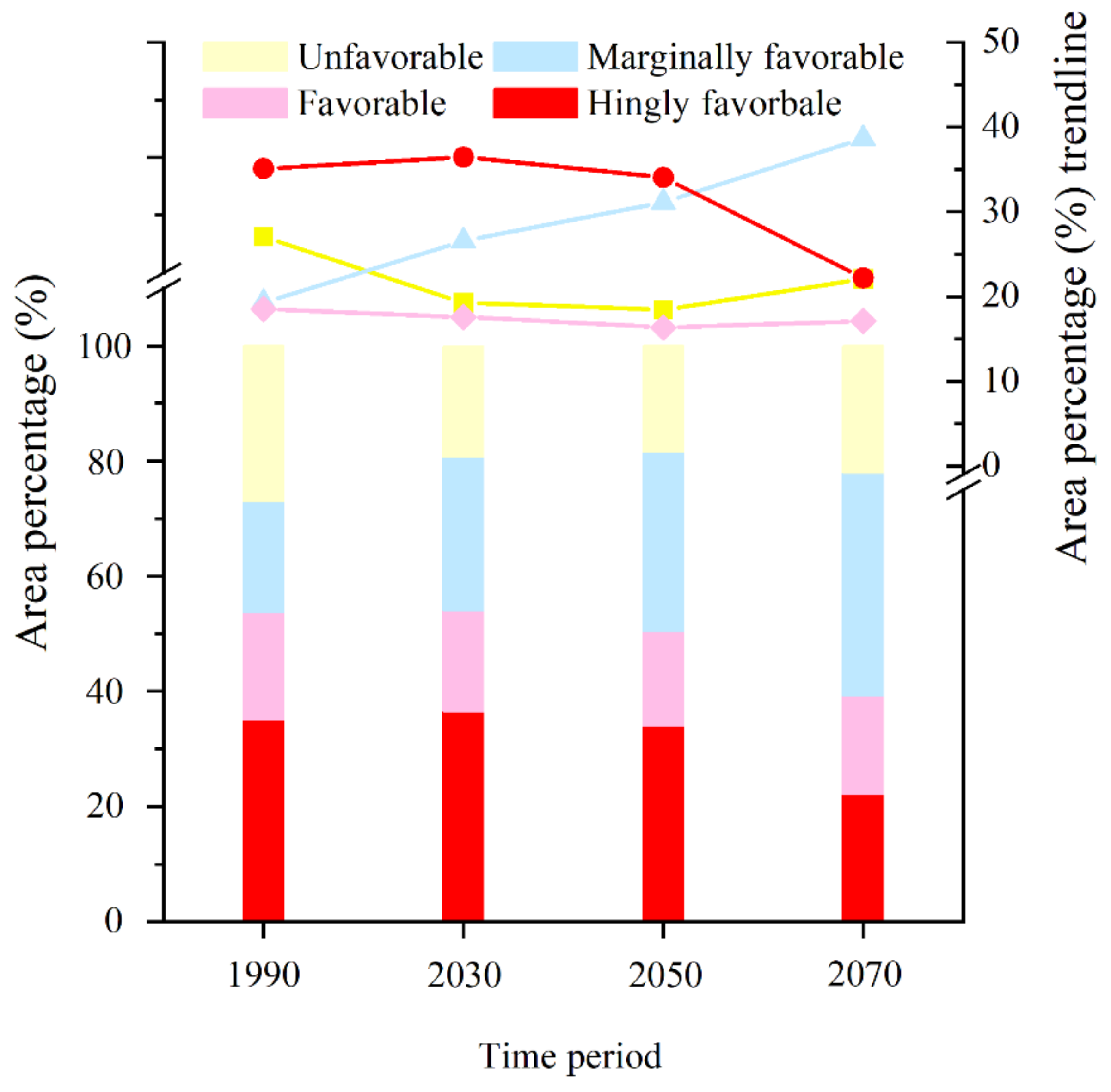
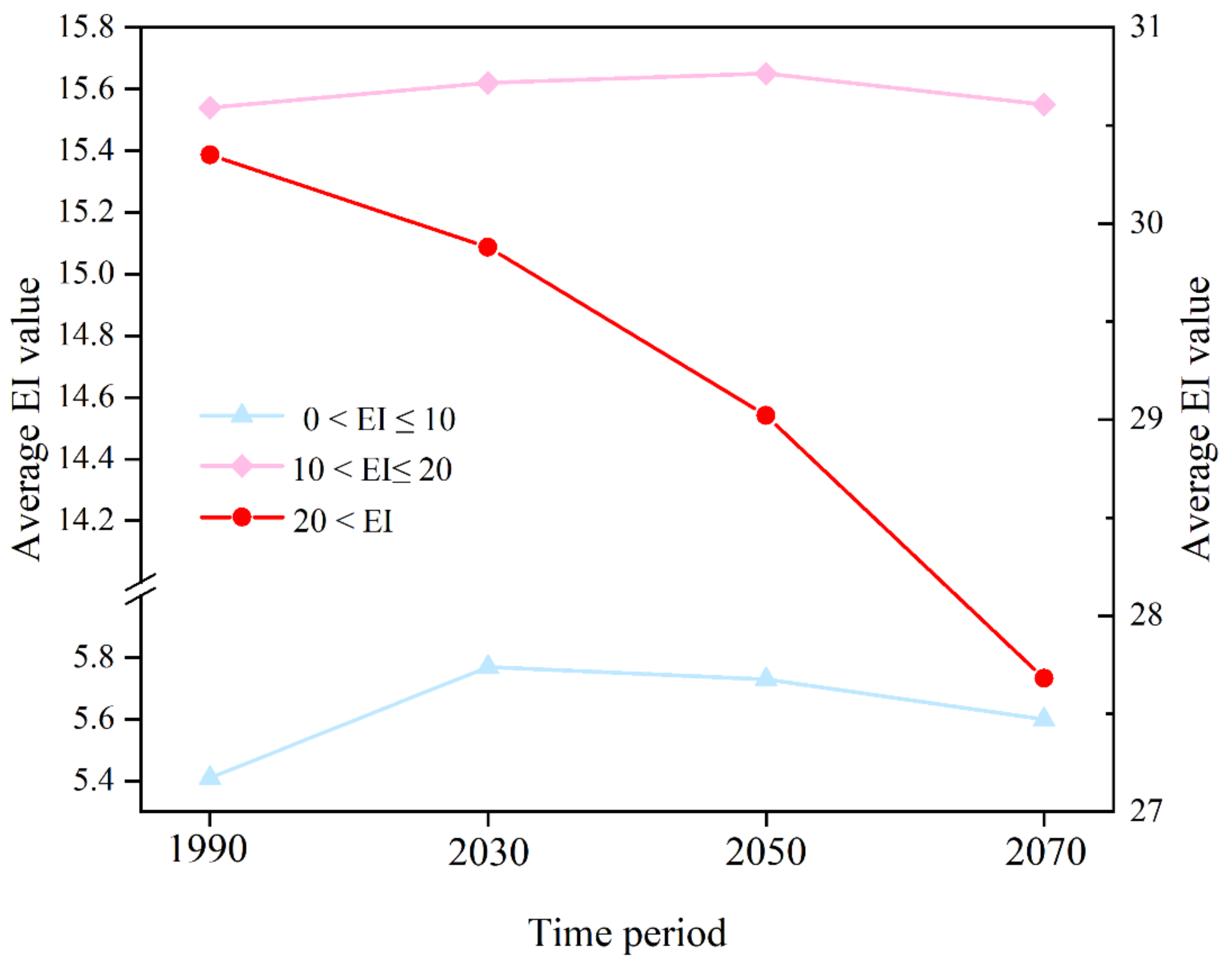
3.3. Change in EI Value on the Potential Distribution under Future Climate Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nansen, C.; Phillips, T.W. Attractancy and toxicity of an attracticide for Indianmeal moth, Plodia interpunctella (Lepidoptera: Pyralidae). J. Econ. Entomol. 2004, 97, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Razazzian, S.; Hassani, M.R.; Imani, S.; Shojai, M. Life table parameters of Plodia interpunctella (Lepidoptera: Pyralidae) on four commercial pistachio cultivars. J. Asia-Pac. Entomol. 2015, 18, 55–59. [Google Scholar] [CrossRef]
- Scheff, D.S.; Sehgal, B.; Subramanyam, B. Evaluating penetration ability of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) larvae into multilayer polypropylene packages. Insects 2018, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stejskal, V.; Stara, J.; Pekar, S.; Nesvorna, M.; Hubert, J. Sensitivity of polyphagous (Plodia interpunctella) and stenophagous (Ephestia kuehniella) storage moths to residual insecticides: Effect of formulation and larval age. Insect Sci. 2021, 28, 1734–1744. [Google Scholar] [CrossRef] [PubMed]
- Stejskal, V.; Bostlova, M.; Nesvorna, M.; Volek, V.; Dolezal, V.; Hubert, J. Comparison of the resistance of mono-and multilayer packaging films to stored-product insects in a laboratory test. Food Control 2017, 73, 566–573. [Google Scholar] [CrossRef]
- Baxter, I.H. Entomopathogen Based Autodissemination for the Control of Plodia interpunctella (Hübner): An Examination of the Critical Components. Ph.D. Thesis, University of Southampton, Southampton, UK, 2008. [Google Scholar]
- Mohandass, S.; Arthur, F.; Zhu, K.; Throne, J.E. Biology and management of Plodia interpunctella (Lepidoptera: Pyralidae) in stored products. J. Stored Prod. Res. 2007, 43, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Ndomo-Moualeu, A.F.; Ulrichs, C.; Radek, R.; Adler, C. Structure and distribution of antennal sensilla in the Indianmeal moth, Plodia interpunctella (Hübner, 1813) (Lepidoptera: Pyralidae). J. Stored Prod. Res. 2014, 59, 66–75. [Google Scholar] [CrossRef]
- Neme, K.; Tola, Y.B.; Mohammed, A.; Tadesse, E.; Astatkie, T. Effects of variety, storage container, and duration on the physical properties, oil content, germination capacity, and seed loss due to Plodia interpunctella infestation of Ethiopian sesame. J. Sci. Food Agric. 2021, 101, 843–852. [Google Scholar] [CrossRef]
- Johnson, J. Effect of relative humidity and product moisture on response of diapausing and nondiapausing Indianmeal moth (Lepidoptera: Pyralidae) larvae to low pressure treatments. J. Econ. Entomol. 2010, 103, 612–618. [Google Scholar] [CrossRef]
- Shao, Y.; Ning, A.; An, Y.; Hou, Z.; Zhou, F.; He, C.; Zhang, J.; Liang, J. Evaluation of the composition and fumigant toxicity against Plodia interpunctella of essential oils from Ajania potaninii and Ajania fruticulosa. J. Asia-Pac. Entomol. 2021, 24, 1235–1238. [Google Scholar] [CrossRef]
- Na, J.H.; Ryoo, M.I. The influence of temperature on development of Plodia interpunctella (Lepidoptera: Pyralidae) on dried vegetable commodities. J. Stored Prod. Res. 2000, 36, 125–129. [Google Scholar] [CrossRef]
- Wang, P.Y. Occurrence of Plodia interpunctella (Hübner) as a new field pest of jujube in peking with notes on its biology. Acta Entomol. Sin. 1964, 13, 628–631. (In Chinese) [Google Scholar] [CrossRef]
- Xia, C.K. Observation on Life History and Habit of Plodia interpunctella (Lepidoptera: Pyralidae) in Alar Region. Xinjiang Agric. Sci. Technol. 1985, 4, 28–29. [Google Scholar]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Piao, S.L.; Ciais, P.; Huang, Y.; Shen, Z.H.; Peng, S.S.; Li, J.S.; Zhou, L.P.; Liu, H.Y.; Ma, Y.C.; Ding, Y.H. The impacts of climate change on water resources and agriculture in China. Nature 2010, 467, 43–51. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.M.; Lei, X.P.; Du, L.X.; Yin, J.; Shi, H.Z.; Zhang, T.; Zhang, Q. Antennae-specific carboxylesterase genes from Indian meal moth: Identification, tissue distribution and the response to semiochemicals. J. Stored Prod. Res. 2019, 84, 101528. [Google Scholar] [CrossRef]
- Chen, E.H.; Tao, Y.X.; Song, W.; Shen, F.; Yuan, M.L.; Tang, P.A. Transcriptome-wide identification of microRNAs and analysis of their potential roles in development of Indian meal moth (Lepidoptera: Pyralidae). J. Econ. Entomol. 2020, 113, 1535–1546. [Google Scholar] [CrossRef]
- Hu, W.L.; Du, J.W. Sex pheromone secondary components of Indian meal moth Plodia interpunctella in China. J. Appl. Ecol. 2005, 16, 1751–1755. (In Chinese) [Google Scholar] [CrossRef]
- Hou, J.C.; Zhao, L.S.; Wang, S.J. Effects of Low Pressure Treatment on the Mortality of Different Life Stages of Indianmeal moth (Plodia interpunctella) and the Quality of Dried Chinese Jujube. Insects 2018, 9, 136. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; Cai, W.L.; Wang, Z.J.; Zhao, J.; Hua, H.X. A new method for evaluating the effects of insecticidal proteins expressed by transgenic plants on ectoparasitoid of target pest. Environ. Sci. Pollut. Res. 2020, 27, 29983–29992. [Google Scholar] [CrossRef]
- Ge, X.Z.; He, S.Y.; Zhu, C.Y.; Wang, T.; Xu, Z.C.; Zong, S.X. Projecting the current and future potential global distribution of Hyphantria cunea (Lepidoptera: Arctiidae) using CLIMEX. Pest Manag. Sci. 2019, 75, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriticos, D.J.; Maywald, G.F.; Yonow, T.; Zurcher, E.J.; Herrmann, N.I.; Sutherst, R. Exploring the effects of climate on plants, animals and diseases. CLIMEX Version 2015, 4, 184. [Google Scholar]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Early, R.; Sax, D.F. Climatic niche shifts between species’ native and naturalized ranges raise concern for ecological forecasts during invasions and climate change. Glob. Ecol. Biogeogr. 2014, 23, 1356–1365. [Google Scholar] [CrossRef]
- Zou, Y.; Ge, X.Z.; Guo, S.W.; Zhou, Y.T.; Wang, T.; Zong, S.X. Impacts of climate change and host plant availability on the global distribution of Brontispa longissima (Coleoptera: Chrysomelidae). Pest Manag. Sci. 2020, 76, 244–256. [Google Scholar] [CrossRef]
- McConnachie, A.; Strathie, L.; Mersie, W.; Gebrehiwot, L.; Zewdie, K.; Abdurehim, A.; Abrha, B.; Araya, T.; Asaregew, F.; Assefa, F. Current and potential geographical distribution of the invasive plant Parthenium hysterophorus (Asteraceae) in eastern and southern Africa. Weed Res. 2011, 51, 71–84. [Google Scholar] [CrossRef]
- Bebber, D.P. Range-expanding pests and pathogens in a warming world. Annu. Rev. Phytopathol. 2015, 53, 335–356. [Google Scholar] [CrossRef]
- Early, R.; Rwomushana, I.; Chipabika, G.; Day, R. Comparing, evaluating and combining statistical species distribution models and CLIMEX to forecast the distributions of emerging crop pests. Pest Manag. Sci. 2022, 78, 671–683. [Google Scholar] [CrossRef]
- Jung, J.M.; Nam, Y.; Jung, S.; Lee, W.H. Spatial analysis of changes in Plodia interpunctella (Lepidoptera: Pyralidae) distribution depending on diets. J. Stored Prod. Res. 2021, 91, 101777. [Google Scholar] [CrossRef]
- Falla, C.M.; Avila, G.A.; McColl, S.T.; Minor, M.; Najar-Rodríguez, A.J. The current and future potential distribution of Gargaphia decoris: A biological control agent for Solanum mauritianum (Solanaceae). Biol. Control 2021, 160, 104637. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Webber, B.L.; Leriche, A.; Ota, N.; Macadam, I.; Bathols, J.; Scott, J.K. CliMond: Global high-resolution historical and future scenario climate surfaces for bioclimatic modelling. Methods Ecol. Evol. 2012, 3, 53–64. [Google Scholar] [CrossRef]
- Jansen, E.; Overpeck, J.; Briffa, K.; Duplessy, J.; Joos, F.; Masson-Delmotte, V.; Olago, D.; Otto-Bliesner, B.; Peltier, W.; Rahmstorf, S. The Physical Science Basis: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007; pp. 433–497. [Google Scholar]
- Santana, P.; Kumar, L.; Da Silva, R.; Picanço, M. Global geographic distribution of Tuta absoluta as affected by climate change. J. Pest Sci. 2019, 92, 1373–1385. [Google Scholar] [CrossRef]
- Da Silva, R.S.; Kumar, L.; Shabani, F.; Picanço, M.C. Potential risk levels of invasive Neoleucinodes elegantalis (small tomato borer) in areas optimal for open-field Solanum lycopersicum (tomato) cultivation in the present and under predicted climate change. Pest Manag. Sci. 2017, 73, 616–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.M.; Chen, X.Y.; Zhu, S.; Sun, M.; Zhou, X.L. Understanding the flavor signature of the rice grown in different regions of China via metabolite profiling. J. Sci. Food. Agric. 2021, 102, 3010–3020. [Google Scholar] [CrossRef] [PubMed]
- Elert, E. Rice by the numbers: A good grain. Nature 2014, 514, S50. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.A.; Wofford, P.L.; Whitehand, L.C. Effect of diet and temperature on development rates, survival, and reproduction of the Indianmeal moth (Lepidoptera: Pyralidae). J. Econ. Entomol. 1992, 85, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.H.; Jung, J.M.; Kim, J.; Lee, H.; Jung, S. Analysis of the spatial distribution and dispersion of Plodia interpunctella (Lepidoptera: Pyralidae) in South Korea. J. Stored Prod. Res. 2020, 86, 101577. [Google Scholar] [CrossRef]
- Feng, Y.; He, W. Studies on elevating the supercooling point of indian meal moth larvae with bacterial ice nucleators. Acta Entomol. Sin. 1996, 39, 53–57. (In Chinese) [Google Scholar] [CrossRef]
- Song, L. Composition Analysis of Essential Oils from Four Spices and EAG of Plodia interpunctlla (Hübuer). Master’s Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2021. (In Chinese) [Google Scholar] [CrossRef]
- Wang, D.X.; Ji, L.; Bai, C.Q.; Bai, X.G.; LI, H.; Yan, L.; Lv, J.H.; Tang, P.; Shao, X.L.; He, Y.P. Investigation on occurrence and distribution of Plodia interpunctella and Sitotroga cerealella in seventy-nine prefevture-level cities of eleven previnces of China. J. Henan Univ. Technol. Nat. Sci. Ed. 2017, 38, 110–114. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, D.H.; Wang, H.Q.; Lou, S. Research on grain production efficiency in China’s main grain-producing areas from the perspective of grain subsidy. Environ. Technol. Innov. 2021, 22, 101530. [Google Scholar] [CrossRef]
- Wei, J.F.; Zhao, Q.; Zhao, W.Q.; Zhang, H.F. Predicting the potential distributions of the invasive cycad scale Aulacaspis yasumatsui (Hemiptera: Diaspididae) under different climate change scenarios and the implications for management. PeerJ 2018, 6, e4832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, W.C.; Li, H.Y.; Wen, J.B. Climate Change Impacts on the Potential Distribution of Apocheima cinerarius (Erschoff) (Lepidoptera: Geometridae). Insects 2022, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Lum, P. High Temperature Inhibition of Development of Eupyrene Sperm and of Reproduction in Plodia interpuntella and Ephestia cautella. J. Ga. Entomol. Soc. 1977, 12, 199–201. [Google Scholar]
- Sun, S.; Wang, Z.Y.; Hu, C.Y.; Gao, G. Understanding Climate Hazard Patterns and Urban Adaptation Measures in China. Sustainability 2021, 13, 13886. [Google Scholar] [CrossRef]
- Zhang, X.H.; Wang, Z.L.; Hou, F.H.; Yang, J.Y.; Guo, X.W. Terrain evolution of China seas and land since the Indo-china movement and characteristics of the stepped landform. Chin. J. Geophys. 2014, 57, 3968–3980. [Google Scholar] [CrossRef]
- Li, H.; Song, W. Spatiotemporal Distribution and Influencing Factors of Ecosystem Vulnerability on Qinghai-Tibet Plateau. Int. J. Environ. Res. Public Health 2021, 18, 6508. [Google Scholar] [CrossRef]
- Xu, D.Y.; You, X.Y.; Xia, C.L. Assessing the spatial-temporal pattern and evolution of areas sensitive to land desertification in North China. Ecol. Indic. 2019, 97, 150–158. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.Q.; Li, Z.; Hou, M.Y.; Dong, G.F.; Liu, T.Y.; Sun, T.; Grattan, K.T. Quasi-distributed fiber optic temperature and humidity sensor system for monitoring of grain storage in granaries. IEEE Sens. J. 2020, 20, 9226–9233. [Google Scholar] [CrossRef]
- Zhang, X.X.; Zhang, H.; Wang, Z.Q.; Chen, X.; Chen, Y. Research on the temperature field of grain piles in underground grain silos lined with plastic. J. Food Process. Eng. 2022, 45, e13971. [Google Scholar] [CrossRef]
- Manandhar, A.; Milindi, P.; Shah, A. An overview of the post-harvest grain storage practices of smallholder farmers in developing countries. Agriculture 2018, 8, 57. [Google Scholar] [CrossRef] [Green Version]
- Neethirajan, S.; Karunakaran, C.; Jayas, D.S.; White, N. Detection techniques for stored-product insects in grain. Food Control 2007, 18, 157–162. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Feng, J.C.; Han, F.; Wu, W.F.; Gao, S.C. Analysis and prediction of grain temperature from air temperature to ensure the safety of grain storage. Int. J. Food Prop. 2020, 23, 1200–1213. [Google Scholar] [CrossRef]
- Arthur, F.H.; Campbell, J.F.; Toews, M.D. Distribution, abundance, and seasonal patterns of Plodia interpunctella (Hübner) in a commercial food storage facility. J. Stored Prod. Res. 2013, 53, 7–14. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Z.; Régnière, J.; Vasseur, L.; Lin, J.; Huang, S.; Ke, F.; Chen, S.; Li, J.; Huang, J.; et al. Large-scale genome-wide study reveals climate adaptive variability in a cosmopolitan pest. Nat. Commun. 2021, 12, 7206. [Google Scholar] [CrossRef]
- Ye, Y.J.; Qi, Q.W.; Jiang, L.L.; Li, X.F. Spatial–temporal changes in grain input–output and the driving mechanism in China since 1985. Int. J. Agric. Sustain. 2017, 15, 445–456. [Google Scholar] [CrossRef]
- Biancolillo, A.; Firmani, P.; Bucci, R.; Magrì, A.; Marini, F. Determination of insect infestation on stored rice by near infrared (NIR) spectroscopy. Microchem. J. 2019, 145, 252–258. [Google Scholar] [CrossRef]
- Tefera, T.; Kanampiu, F.; Groote, H.D.; Hellin, J.; Mugo, S.; Kimenju, S.; Beyene, Y.; Boddupalli, P.M.; Shiferaw, B.; Banziger, M. The metal silo: An effective grain storage technology for reducing post-harvest insect and pathogen losses in maize while improving smallholder farmers’ food security in developing countries. Crop Prot. 2011, 30, 240–245. [Google Scholar] [CrossRef]
- Ayvaz, A.; Albayrak, S.; Karaborklu, S. Gamma radiation sensitivity of the eggs, larvae and pupae of Indian meal moth Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae). Pest Manag. Sci. 2010, 64, 505–512. [Google Scholar] [CrossRef]
- Hansen, L.S.; Hansen, P.; Jensen, K.M.V. Lethal doses of ozone for control of all stages of internal and external feeders in stored products. Pest Manag. Sci. 2012, 68, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, D.; Jian, F.; Jayas, D.S.; Chen, C.; Wang, D. Mortality of Different Stages of Plodia interpunctella (Lepidoptera: Pyralidae) at Three Temperatures in Controlled Atmosphere of High Nitrogen. J. Econ. Entomol. 2020, 113, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Adarkwah, C.; Sch, M. Biological control of Plodia interpunctella (Lepidoptera: Pyralidae) by single and double releases of two larval parasitoids in bulk stored wheat. J. Stored Prod. Res. 2012, 51, 1–5. [Google Scholar] [CrossRef]
- Environmental Protection Agency (EPA). Protection of stratospheric ozone; process for exempting quarantine and pre-shipment application of methyl bromide. Fed. Regist. 2001, 66, 37752–37769. [Google Scholar]
- Hagstrum, D.W.; Phillips, T.W. Evolution of stored-product entomology: Protecting the world food supply. Annu. Rev. Entomol. 2017, 62, 379–397. [Google Scholar] [CrossRef]
- Boyer, S.; Zhang, H.; Lempérière, G. A review of control methods and resistance mechanisms in stored-product insects. Bull. Entomol. Res. 2012, 102, 213–229. [Google Scholar] [CrossRef]
- Ma, C.S.; Zhang, W.; Peng, Y.; Zhao, F.; Chang, X.Q.; Xing, K.; Zhu, L.; Ma, G.; Yang, H.P.; Rudolf, V.H. Climate warming promotes pesticide resistance through expanding overwintering range of a global pest. Nat. Commun. 2021, 12, 5351. [Google Scholar] [CrossRef]
- Mostafiz, M.M.; Hassan, E.; Acharya, R.; Shim, J.-K.; Lee, K.-Y. Methyl Benzoate Is Superior to Other Natural Fumigants for Controlling the Indian Meal Moth (Plodia interpunctella). Insects 2021, 12, 23. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CLIMEX Parameters | Values | Unit | |
|---|---|---|---|
| Temperature | |||
| DV0 | Starting temperature | 12.5 | °C |
| DV1 | Lower optimal temperature | 20 | °C |
| DV2 | Upper optimal temperature | 30 | °C |
| DV3 | Upper temperature threshold | 35 | °C |
| PDD | Degree days to complete one generation | 461 | °C days |
| Moisture | |||
| SM0 | Lower soil moisture threshold | 0.05 | |
| SM1 | Lower optimal soil moisture | 0.2 | |
| SM2 | Upper optimal soil moisture | 0.8 | |
| SM3 | Upper soil moisture threshold | 1.5 | |
| Heat stress | |||
| TTHS | Heat stress temperature threshold | 35 | °C |
| THHS | Heat stress temperature rate | 0.0048 | Week−1 |
| Cold stress | |||
| TTCS | Cold stress temperature threshold | −12.2 | °C |
| THCS | Cold stress temperature rate | −0.0003 | Week−1 |
| Dry stress | |||
| SMDS | Dry stress threshold | 0.05 | |
| HDS | Dry stress rate | −0.005 | Week−1 |
| Wet stress | |||
| SMWS | Wet stress threshold | 1.5 | |
| HWS | Wet stress rate | 0.002 | Week−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Song, C.; Ma, L.; Yan, X.; Shi, J.; Hao, C. The Impacts of Climate Change on the Potential Distribution of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in China. Insects 2022, 13, 636. https://doi.org/10.3390/insects13070636
Zhao J, Song C, Ma L, Yan X, Shi J, Hao C. The Impacts of Climate Change on the Potential Distribution of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in China. Insects. 2022; 13(7):636. https://doi.org/10.3390/insects13070636
Chicago/Turabian StyleZhao, Jinyu, Chengfei Song, Li Ma, Xizhong Yan, Juan Shi, and Chi Hao. 2022. "The Impacts of Climate Change on the Potential Distribution of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in China" Insects 13, no. 7: 636. https://doi.org/10.3390/insects13070636
APA StyleZhao, J., Song, C., Ma, L., Yan, X., Shi, J., & Hao, C. (2022). The Impacts of Climate Change on the Potential Distribution of Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) in China. Insects, 13(7), 636. https://doi.org/10.3390/insects13070636