
Foreleg Transcriptomic Analysis of the Chemosensory Gene Families in Plagiodera versicolora (Coleoptera: Chrysomelidae)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Tissue Collection
2.2. RNA Extraction, cDNA Library Preparation, and Sequencing
2.3. Assembly and Functional Annotation
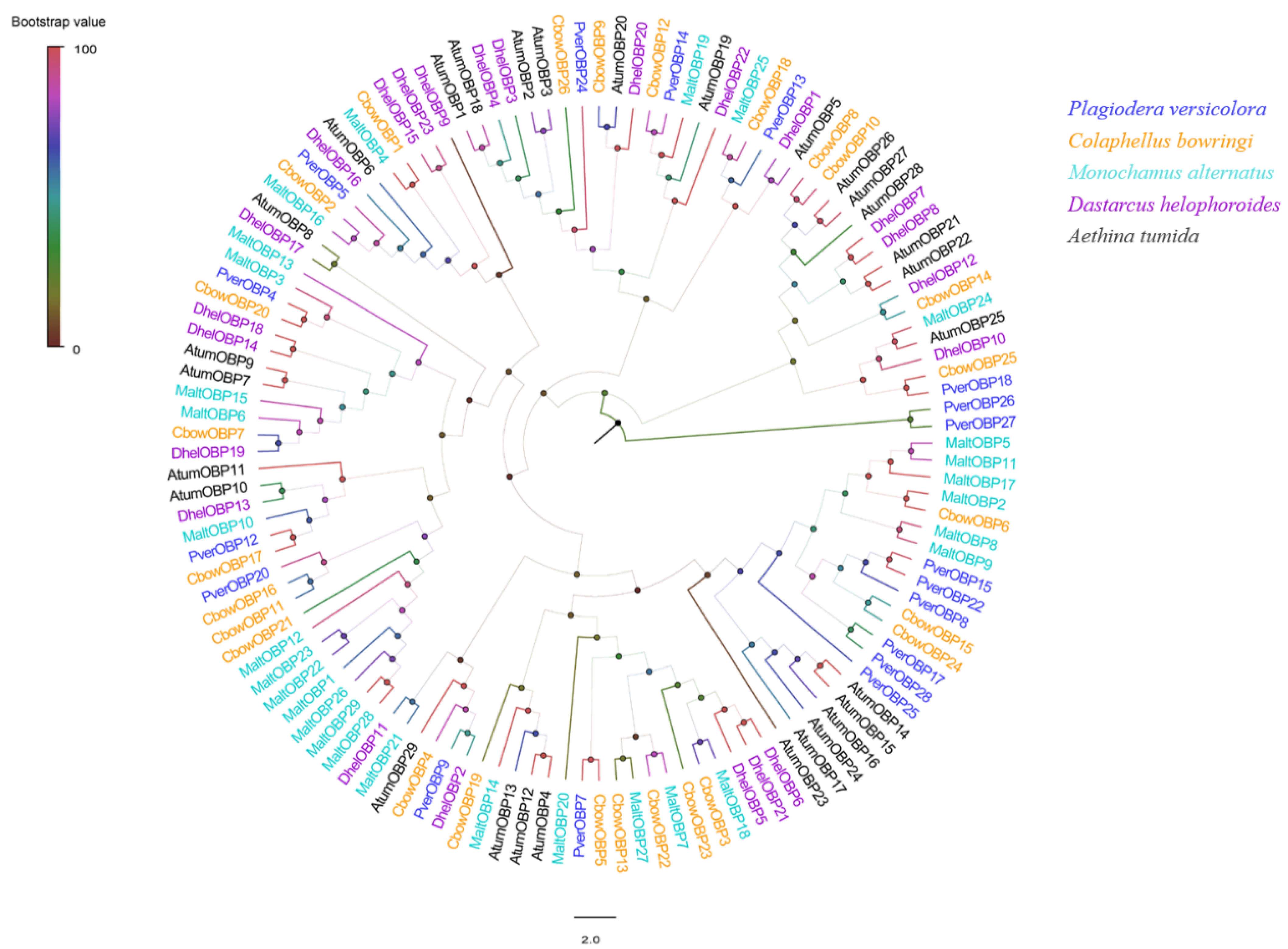
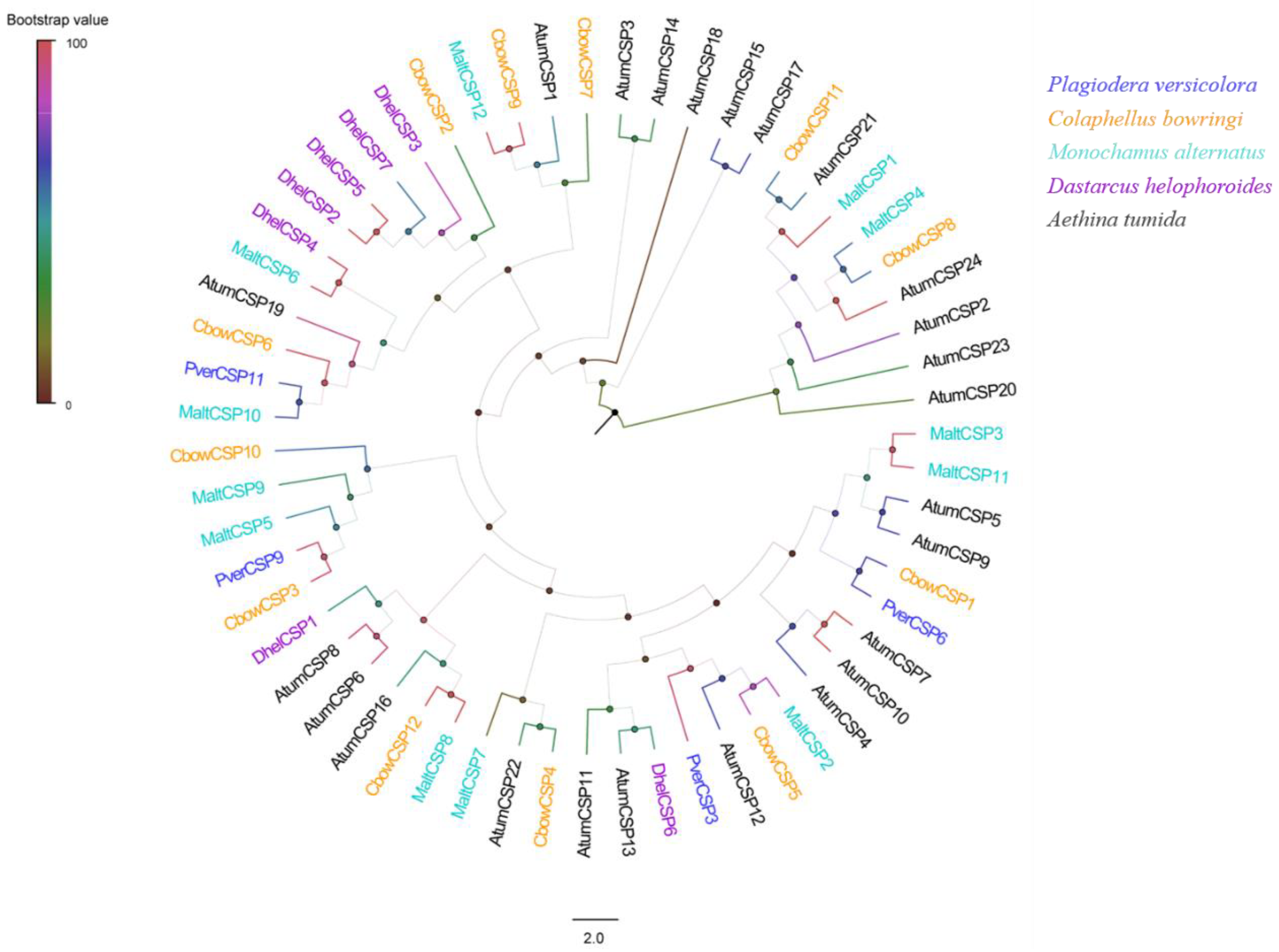
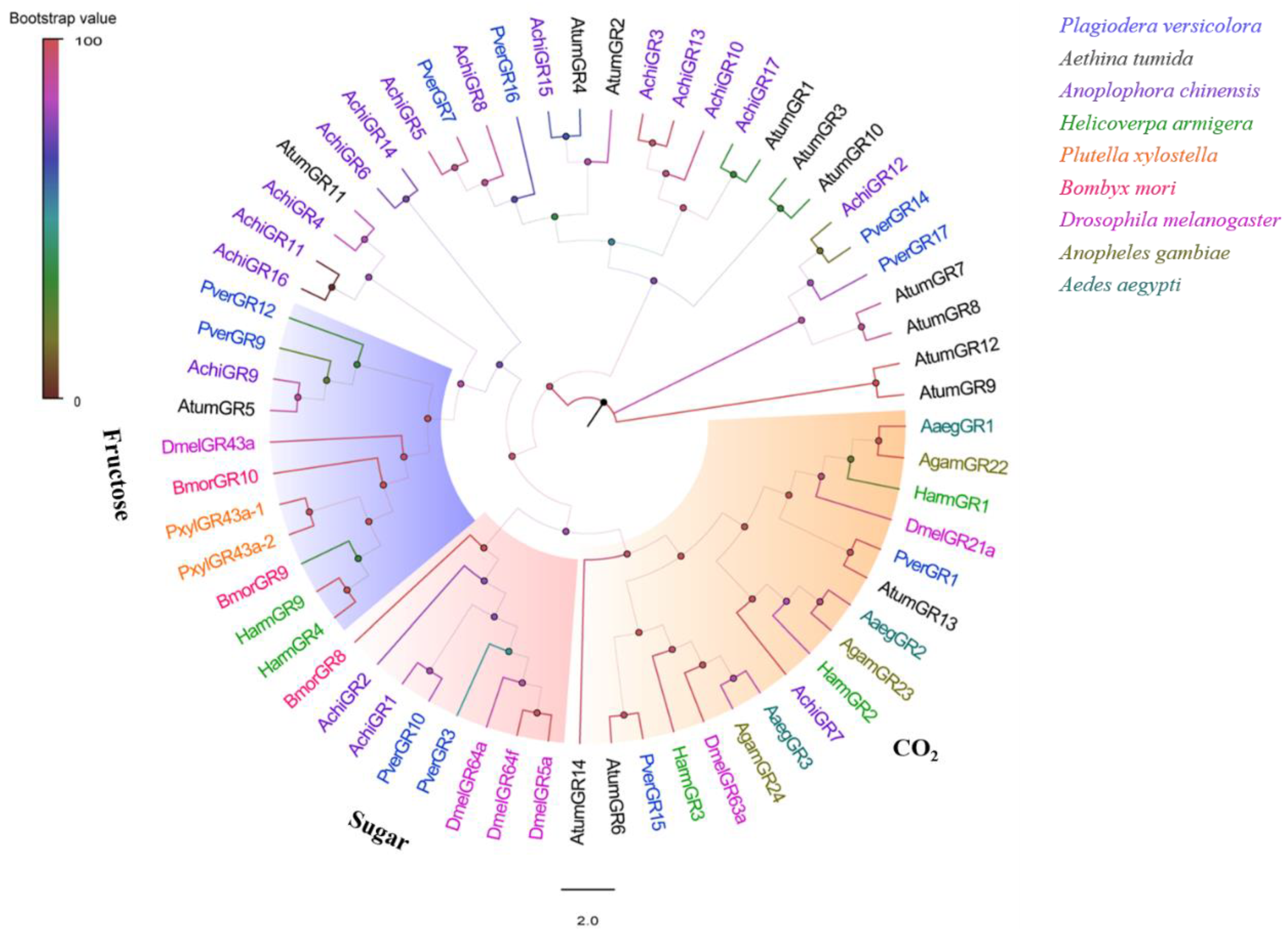
2.4. Sequence Analysis and Phylogenetic Tree Construction
2.5. Expression Analysis by Real-Time Quantitative PCR
3. Results
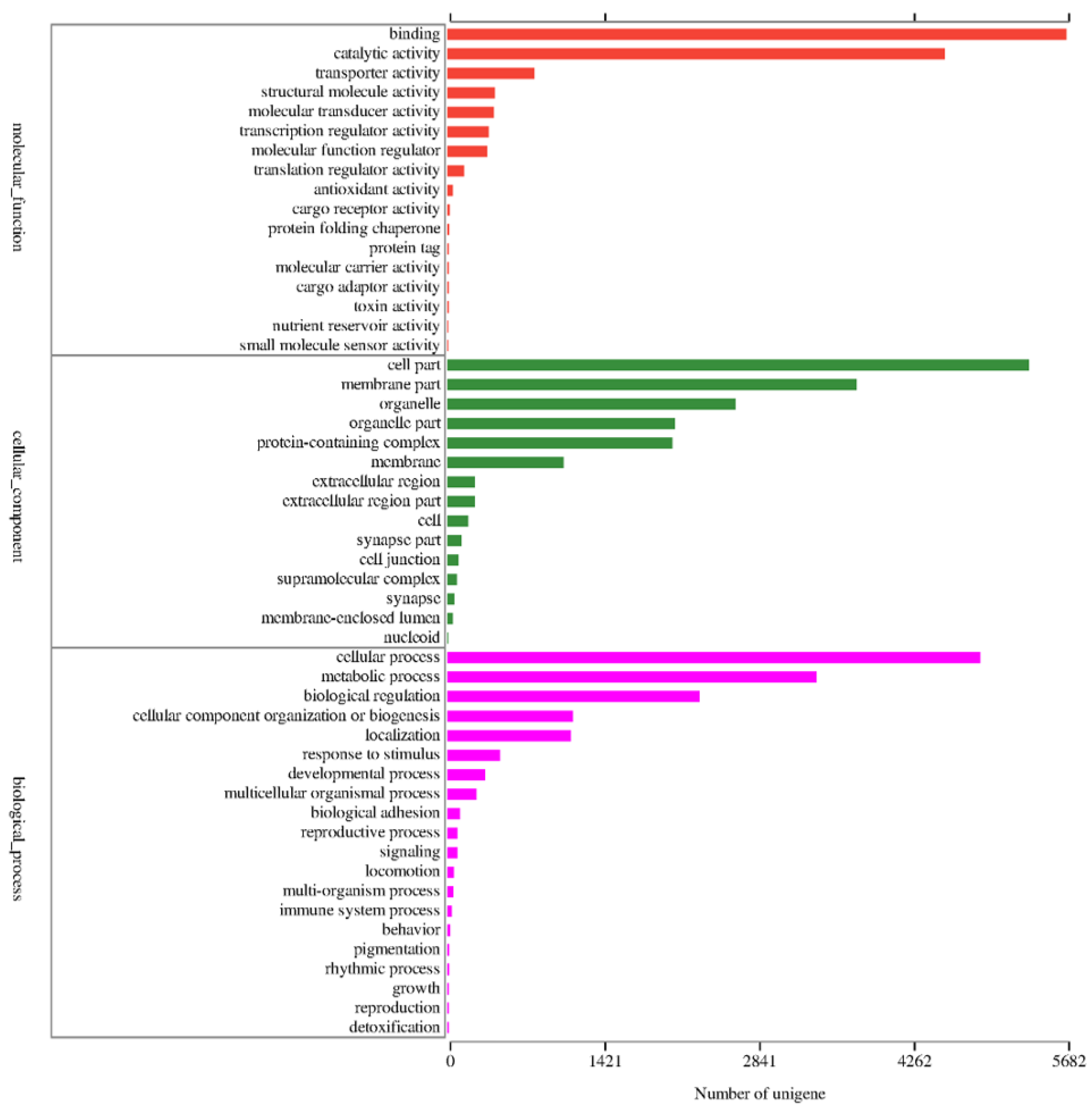
3.1. Transcriptome Analysis and Assembly
3.2. Identification of Putative OBP Genes
3.3. Identification of Putative CSP Genes
3.4. Identification of Putative OR Genes
3.5. Identification of Putative GR Genes
3.6. Identification of Putative IR Genes
3.7. Identification of Putative SNMP Genes
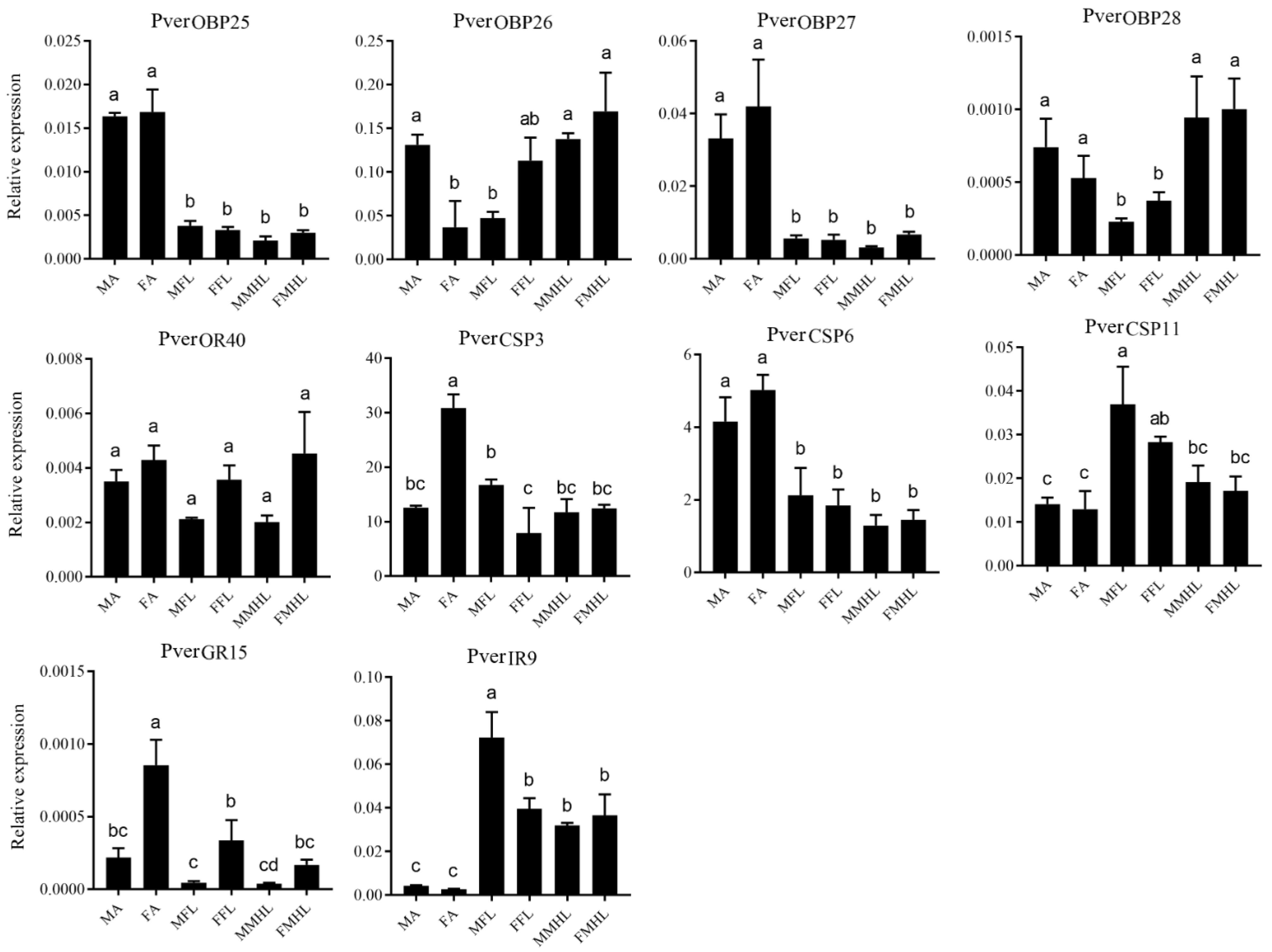
3.8. Tissue Specific Expression Analysis of Putative Chemosensory Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Novotny, V.; Miller, S.E.; Hulcr, J.; Drew, R.A.I.; Basset, Y.; Janda, M.; Setliff, G.P.; Darrow, K.; Stewart, A.J.A.; Auga, J.; et al. Low beta diversity of herbivorous insects in tropical forests. Nature 2007, 448, 692–695. [Google Scholar] [CrossRef]
- Meyerhof, W. Chemosensory Systems in Mammals, Fishes, and Insects; Springer: Heidelberg/Berlin, Germany, 2009; Volume 47. [Google Scholar]
- Liu, X.-L.; Sun, S.-J.; Khuhro, S.A.; Elzaki, M.E.A.; Yan, Q.; Dong, S.-L. Functional characterization of pheromone receptors in the moth Athetis dissimilis (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2019, 158, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-L.; Yan, Q.; Yang, Y.-L.; Hou, W.; Miao, C.-L.; Peng, Y.-C.; Dong, S.-L. A Gustatory receptor GR8 tunes specifically to D-fructose in the common cutworm Spodoptera Litura. Insects 2019, 10, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.-J. Odorant-binding proteins in insects. Vitam. Horm. 2010, 83, 241–272. [Google Scholar]
- Vogt, R.G. Molecular basis of pheromone detection in insects. Compr. Mol. Insect Sci. 2005, 3–6, 753–803. [Google Scholar]
- Vogt, R.G.; Miller, N.E.; Litvack, R.; Fandino, R.A.; Sparks, J.; Staples, J.; Friedman, R.; Dickens, J.C. The insect SNMP gene family. Insect Biochem. Mol. Biol. 2009, 39, 448–456. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhou, J.-J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Robertson, H.M.; Warr, C.G.; Carlson, J.R. Molecular evolution of the insect chemoreceptor gene superfamily in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2003, 100 (Suppl. 2), 14537–14542. [Google Scholar] [CrossRef] [Green Version]
- Vieira, F.G.; Rozas, J. Comparative genomics of the odorant-binding and chemosensory protein gene families across the Arthropoda: Origin and evolutionary history of the chemosensory system. Genome Biol. Evol. 2011, 3, 476–490. [Google Scholar] [CrossRef] [PubMed]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. A typical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef] [Green Version]
- Engsontia, P.; Sangket, U.; Chotigeat, W.; Satasook, C. Molecular evolution of the odorant and gustatory receptor genes in Lepidopteran insects: Implications for their adaptation and speciation. J. Mol. Evol. 2014, 79, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Chyb, S.; Dahanukar, A.; Wickens, A.; Carlson, J.R. Drosophila Gr5a encodes a taste receptor tuned to trehalose. Proc. Natl. Acad. Sci. USA 2003, 100 (Suppl. 2), 14526–14530. [Google Scholar] [CrossRef] [Green Version]
- Crava, C.M.; Ramasamy, S.; Ometto, L.; Anfora, G.; Rota-Stabelli, O. Evolutionary insights into taste perception of the invasive pest Drosophila suzukii. G3 2016, 6, 4185–4196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebbs, M.L.; Amrein, H. Taste and pheromone perception in the fruit fly Drosophila melanogaster. Pflug. Arch. 2007, 454, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Kent, L.B.; Robertson, H.M. Evolution of the sugar receptors in insects. BMC Evol. Biol. 2009, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient protostomeorigin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef] [Green Version]
- Abuin, L.; Bargeton, B.; Ulbrich, M.H.; Isacoff, E.Y.; Kellenberger, S.; Benton, R. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 2011, 69, 44–60. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.A.; Ignell, R.; Carlson, J.R. Chemosensory coding by neurons in the coeloconic sensilla of the Drosophila antenna. J. Neurosci. 2005, 25, 8359–8367. [Google Scholar] [CrossRef]
- Min, S.; Ai, M.; Shin, S.A.; Suh, G.S.B. Dedicated olfactory neurons mediating attraction behavior to ammonia and amines in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, E1321–E1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Bisch-Knaden, S.; Fandino, R.A.; Yan, S.; Obiero, G.F.; Grosse-Wilde, E.; Hansson, B.S.; Knaden, M. The olfactory coreceptor Ir8a governs larval feces-mediated competition avoidance in a hawkmoth. Proc. Natl. Acad. Sci. USA 2019, 116, 21828–21833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.V.; Ni, J.; Montell, C. The molecular basis for attractive salt-taste coding in Drosophila. Science 2013, 340, 1334–1338. [Google Scholar] [CrossRef] [Green Version]
- Ni, L.; Klein, M.; Svec, K.V.; Budelli, G.; Chang, E.C.; Ferrer, A.J.; Benton, R.; Samuel, A.D.; Garrity, P.A. The ionotropic receptors Ir21a and Ir25a mediate cool sensing in Drosophila. eLife 2016, 5, e13254. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Pang, L.; Duong, V.-K.; Lee, A.; Schoniger, H.; Varady, E.; Dahanukar, A. A molecular and cellular context-dependent role for Ir76b in detection of amino acid taste. Cell Rep. 2017, 18, 737–750. [Google Scholar] [CrossRef]
- Seada, M.A.; Ignell, R.; Al Assiuty, A.N.; Anderson, P. Functional characterization of the gustatory sensilla of tarsi of the female polyphagous moth Spodoptera littoralis. Front. Physiol. 2018, 9, 1606. [Google Scholar] [CrossRef]
- Chun, M.W.; Schoonhoven, L.M. Tarsal contact chemosensory hairs of the large white butterfly Pieris brassicae and their possible role in oviposition behaviour. Entomol. Exp. Appl. 1973, 16, 343–357. [Google Scholar] [CrossRef]
- Nayak, S.V.; Singh, R.N. Sensilla on the tarsal segments and mouthparts of adult Drosophila Melanogaster meigen (Diptera: Drosophilidae). Int. J. Insect Morphol. Embryol. 1983, 12, 273–291. [Google Scholar] [CrossRef]
- Amrein, H.; Thorne, N. Gustatory perception and behavior in Drosophila melanogaster. Curr. Biol. 2005, 15, R673–R684. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-F.; van Loon, J.J.A.; Wang, C.-Z. Tarsal taste neuron activity and proboscis extension reflex in response to sugars and amino acids in Helicoverpa armigera (Hubner). J. Exp. Biol. 2010, 213, 2889–2895. [Google Scholar] [CrossRef] [Green Version]
- Rondoni, G.; Roman, A.; Meslin, C.; Montagné, N.; Conti, E.; Jacquin-Joly, E. Antennal transcriptome analysis and identification of candidate chemosensory genes of the harlequin ladybird beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Insects 2021, 12, 209. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.; Johny, J.; Walker, W.B.; Guan, Q.; Mfarrej, S.; Jakše, J.; Montagné, N.; Jacquin-Joly, E.; Alqarni, A.S.; Al-Saleh, M.A.; et al. Antennal transcriptome sequencing and identification of candidate chemoreceptor proteins from an invasive pest, the american palm weevil, Rhynchophorus palmarum. Sci. Rep. 2021, 11, 8334. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, S.; Ando, Y.; Ohgushi, T. Evolution of feeding preference in a leaf beetle: The importance of phenotypic plasticity of a host plant. Ecol. Lett. 2009, 12, 920–929. [Google Scholar] [CrossRef]
- Liu, X.; Tong, N.; Wu, Z.; Li, Y.; Ma, M.; Liu, P.; Lu, M. Identification of chemosensory genes based on the antennal transcriptomic analysis of Plagiodera versicolora. Insects 2021, 13, 36. [Google Scholar] [CrossRef]
- Li, X.-M.; Zhu, X.-Y.; Wang, Z.-Q.; Wang, Y.; He, P.; Chen, G.; Sun, L.; Deng, D.-G.; Zhang, Y.-N. Candidate chemosensory genes identified in Colaphellus bowringi by antennal transcriptome analysis. BMC Genom. 2015, 16, 1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Zhai, X.; Li, L.; Li, Q.; Liu, F.; Zhao, H. Identification and expression profile of chemosensory genes in the small hive beetle Aethina tumida. Insects 2021, 12, 661. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.-Z.; Min, S.-F.; Mi, F.; Zhou, S.-S.; Wang, M.-Q. Analysis of chemosensory gene families in the beetle Monochamus alternatus and its parasitoid Dastarcus helophoroides. Comp. Biochem. Physiol. Part D Genom. Proteom. 2014, 11, 1–8. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Y.-N.; Qian, J.-L.; Kang, K.; Zhang, X.-Q.; Deng, J.-D.; Tang, Y.-P.; Chen, C.; Hansen, L.; Xu, T.; et al. Identification and expression patterns of Anoplophora chinensis (Forster) chemosensory receptorgenes from the antennal transcriptome. Front. Physiol. 2018, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-L.; Sun, S.-J.; Hou, W.; Zhang, J.; Yan, Q.; Dong, S.-L. Functional characterization of two spliced variants of fructose gustatory receptor in the diamondback moth, Plutella xylostella. Pestic. Biochem. Physiol. 2020, 164, 7–13. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, H.-J.; Anderson, A. A sugar gustatory receptor identified from the foregut of cotton bollworm Helicoverpa armigera. J. Chem. Ecol. 2012, 38, 1513–1520. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Anderson, A. Carbon dioxide receptor genes in cotton bollworm Helicoverpa armigera. Naturwissenschaften 2015, 102, 11. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-J.; Ning, C.; Guo, H.; Jia, Y.-Y.; Huang, L.-Q.; Qu, M.-J.; Wang, C.-Z. A gustatory receptor tuned to D-fructose in antennal sensilla chaetica of Helicoverpa armigera. Insect Biochem. Mol. Biol. 2015, 60, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Tanaka, K.; Touhara, K. Sugar-regulated cation channel formed by an insect gustatory receptor. Proc. Natl. Acad. Sci. USA 2011, 108, 11680–11685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.-J.; Anderson, A.R.; Trowell, S.C.; Luo, A.-R.; Xiang, Z.-H.; Xia, Q.-Y. Topological and functional characterization of an insect gustatory receptor. PLoS ONE 2011, 6, e24111. [Google Scholar]
- Kikuta, S.; Endo, H.; Tomita, N.; Takada, T.; Morita, C.; Asaoka, K.; Sato, R. Characterization of a ligand-gated cation channel based on an inositol receptor in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2016, 74, 12–20. [Google Scholar] [CrossRef]
- Jiao, Y.; Moon, S.J.; Wang, X.; Ren, Q.; Montell, C. Gr64f is required in combination with other gustatory receptors for sugar detection in Drosophila. Curr. Biol. 2008, 18, 1797–1801. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Moon, S.J.; Montell, C. A Drosophila gustatory receptor required for the responses to sucrose, glucose, and maltose identified by mRNA tagging. Proc. Natl. Acad. Sci. USA 2007, 104, 14110–14115. [Google Scholar] [CrossRef] [Green Version]
- Jones, W.D.; Cayirlioglu, P.; Kadow, I.G.; Vosshall, L.B. Two chemosensory receptors together mediate carbon dioxide detection in Drosophila. Nature 2007, 445, 86–90. [Google Scholar] [CrossRef]
- Miyamoto, T.; Slone, J.; Song, X.; Amrein, H. A fructose receptor functions as a nutrient sensor in the Drosophila brain. Cell 2012, 151, 1113–1125. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.; Qiu, Y.T.; Wang, G.; Kwon, J.Y.; Rutzler, M.; Kwon, H.-W.; Pitts, R.J.; van Loon, J.J.A.; Takken, W.; Carlson, J.R.; et al. Odor coding in the maxillary palp of the malaria vector mosquito Anopheles gambiae. Curr. Biol. 2007, 17, 1533–1544. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.; Tu, C.; Luo, J.; Lu, M.; Zhang, S.; Xu, L. Metabolic and immunological effects of gut microbiota in leaf beetles at the local and systemic levels. Integr. Zool. 2021, 16, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sparks, J.T.; Bohbot, J.D.; Dickens, J.C. The genetics of chemoreception in the labella and tarsi of Aedes aegypti. Insect Biochem. Mol. Biol. 2014, 48, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, Q.; Wang, Q.; Dong, K.; Xiao, Y.; Zhang, Y.-J. Identification and characterization of odorant binding proteins in the forelegs of Adelphocoris lineolatus (Goeze). Front. Physiol. 2017, 8, 735. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Xu, K.; Ma, W.; Su, W.; Tai, M.; Zhao, H.; Jiang, Y.; Li, X. Contact chemosensory genes identified in leg transcriptome of Apis cerana cerana (Hymenoptera: Apidae). J. Econ. Entomol. 2019, 112, 2015–2029. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; An, X.; Wang, Q.; Khashaveh, A.; Gu, S.; Liu, S.; Zhang, Y. Identification of leg chemosensory genes and sensilla in the Apolygus lucorum. Front. Physiol. 2020, 11, 276. [Google Scholar] [CrossRef]
- Li, L.-L.; Huang, J.-R.; Xu, J.-W.; Yao, W.-C.; Yang, H.-H.; Shao, L.; Zhang, H.-R.; Dewer, Y.; Zhu, X.-Y.; Zhang, Y.-N. Ligand-binding properties of odorant-binding protein 6 in Athetis lepigone to sex pheromones and maize volatiles. Pest Manag. Sci. 2022, 78, 52–62. [Google Scholar] [CrossRef]
- Zhou, J.-J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx mori odorant-binding proteins reveals that a general odorant-binding protein discriminates between sex pheromone components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, Q.; Li, H.; Sun, L.; Zhang, D.; Zhang, Y. Sensilla localization and sex pheromone recognition of odorant binding protein OBP4 in the mirid plant bug Adelphocoris lineolatus (Goeze). J. Insect Physiol. 2020, 121, 104012. [Google Scholar] [CrossRef]
- Hua, J.-F.; Zhang, S.; Cui, J.-J.; Wang, D.-J.; Wang, C.-Y.; Luo, J.-Y.; Lv, L.-M.; Ma, Y. Functional characterizations of one odorant binding protein and three chemosensory proteins from Apolygus lucorum (Meyer-Dur) (Hemiptera: Miridae) legs. J. Insect Physiol. 2013, 59, 690–696. [Google Scholar] [CrossRef]
- Younas, A.; Waris, M.I.; Tahir Ul Qamar, M.; Shaaban, M.; Prager, S.M.; Wang, M.-Q. Functional analysis of the chemosensory protein MsepCSP8 from the oriental armyworm Mythimna separata. Front. Physiol. 2018, 9, 872. [Google Scholar] [CrossRef] [PubMed]
- Kitabayashi, A.N.; Arai, T.; Kubo, T.; Natori, S. Molecular cloning of cDNA for P10, a novel protein that increases in the regenerating legs of Periplaneta americana (American Cockroach). Insect Biochem. Mol. Biol. 1998, 28, 785–790. [Google Scholar] [CrossRef]
- Wanner, K.W.; Isman, M.B.; Feng, Q.; Plettner, E.; Theilmann, D.A. Developmental expression patterns of four chemosensory protein genes from the eastern spruce budworm, Chroistoneura fumiferana. Insect Mol. Biol. 2005, 14, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.-X.; Qin, G.-X.; Liu, T.; Mei, X.-L.; Li, B.; Shen, Z.-Y.; Guo, X.-J. Differentially expressed genes in the cuticle and hemolymph of the silkworm, Bombyx mori, injected with the fungus Beauveria bassiana. J. Insect Sci. 2013, 13, 138. [Google Scholar] [CrossRef] [PubMed]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef] [Green Version]
- Rimal, S.; Sang, J.; Poudel, S.; Thakur, D.; Montell, C.; Lee, Y. Mechanism of acetic acid gustatory repulsion in Drosophila. Cell Rep. 2019, 26, 1432–1442.e4. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Luo, Y.; Shang, X.; Sun, J.S.; Carlson, J.R. Chemosensory sensilla of the Drosophila wing express a candidate ionotropic pheromone receptor. PLoS Biol. 2019, 17, e2006619. [Google Scholar] [CrossRef]
- Lee, Y.; Poudel, S.; Kim, Y.; Thakur, D.; Montell, C. Calcium taste avoidance in Drosophila. Neuron 2018, 97, 67–74.e4. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, A.H.; Stanley, M.; Weiss, Z.F.; Musso, P.-Y.; Chan, R.C.; Zhang, H.; Feldman-Kiss, D.; Gordon, M.D. A complex peripheral code for salt taste in Drosophila. eLife 2018, 7, e37167. [Google Scholar] [CrossRef]
- Stewart, S.; Koh, T.-W.; Ghosh, A.C.; Carlson, J.R. Candidate ionotropic taste receptors in the Drosophila larva. Proc. Natl. Acad. Sci. USA 2015, 112, 4195–4201. [Google Scholar] [CrossRef] [Green Version]
- Briscoe, A.D.; Macias-Muñoz, A.; Kozak, K.M.; Walters, J.R.; Yuan, F.; Jamie, G.A.; Martin, S.H.; Dasmahapatra, K.K.; Ferguson, L.C.; Mallet, J.; et al. Female behaviour drives expression and evolution of gustatory receptors in butterflies. PLoS Genet. 2013, 9, e1003620. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.; Yang, K.; Xu, M.; Huang, L.Q.; Wang, C.Z. Functional validation of the carbon dioxide receptor in labial palps of Helicoverpa armigera moths. Insect Biochem. Mol. Biol. 2016, 73, 12–19. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Tong, N.; Li, Y.; Guo, J.; Lu, M.; Liu, X. Foreleg Transcriptomic Analysis of the Chemosensory Gene Families in Plagiodera versicolora (Coleoptera: Chrysomelidae). Insects 2022, 13, 763. https://doi.org/10.3390/insects13090763
Wu Z, Tong N, Li Y, Guo J, Lu M, Liu X. Foreleg Transcriptomic Analysis of the Chemosensory Gene Families in Plagiodera versicolora (Coleoptera: Chrysomelidae). Insects. 2022; 13(9):763. https://doi.org/10.3390/insects13090763
Chicago/Turabian StyleWu, Zheran, Na Tong, Yang Li, Jinmeng Guo, Min Lu, and Xiaolong Liu. 2022. "Foreleg Transcriptomic Analysis of the Chemosensory Gene Families in Plagiodera versicolora (Coleoptera: Chrysomelidae)" Insects 13, no. 9: 763. https://doi.org/10.3390/insects13090763
APA StyleWu, Z., Tong, N., Li, Y., Guo, J., Lu, M., & Liu, X. (2022). Foreleg Transcriptomic Analysis of the Chemosensory Gene Families in Plagiodera versicolora (Coleoptera: Chrysomelidae). Insects, 13(9), 763. https://doi.org/10.3390/insects13090763