
Role of Lipids in Phosphine Resistant Stored-Grain Insect Pests Tribolium castaneum and Rhyzopertha dominica

 ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Cultures
2.2. Preparation of Phosphine Gas for Determination of Resistance Factor
2.3. Chemicals and Apparatuses
2.4. Extraction Procedures
2.5. Alalysis of Lipids with Ultra Performance Liquid Chromatography-Quadrupole-Mass Spectrometry (UPLC-Q-ToF-MS) and Analytical Conditions
2.6. Data Processing and Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donahaye, E.J.; Bell, C.; Jayes, D.; Noyas, R.; Phillips, T.W. Integrated pest management strategies used in stored grains in Brazil to manage phosphine resistance. In Proceedings of the International Conference Controlled Atmosphere and Fumigation in Stored Product, Gold Coast, Australia, 8–13 August 2007; pp. 293–300. [Google Scholar]
- Edde, P.A. A review of the biology and control of Rhyzopertha dominica (F.) the lesser grain borer. J. Stored Prod. Res. 2012, 48, 1–18. [Google Scholar] [CrossRef]
- Hameed, A.; Freed, S.; Hussain, A.; Iqbal, M.; Hussain, M.; Naeem, M.; Sajjad, A.; Hussnain, H.; Sadiq, M.A.; Tipu, A.L. Toxicological effects of neem (Azadirachta indica), Kanair (Nerium oleander) and spinosad (Tracer 240 SC) on the red flour beetle (Tribolium castaneum) (Herbst.). Afr. J. Agric. Res. 2012, 7, 555–560. [Google Scholar]
- Oppert, B.; Guedes, R.N.; Aikins, M.J.; Perkin, L.; Chen, Z.; Phillips, T.W.; Zhu, K.Y.; Opit, G.P.; Hoon, K.; Sun, Y. Genes related to mitochondrial functions are differentially expressed in phosphine-resistant and-susceptible Tribolium castaneum. BMC Genom. 2015, 16, 968. [Google Scholar] [CrossRef]
- Chaudhry, M. A Review of the Mechanisms Involved in the Action of Phosphine as an Insecticide and Phosphine Resistance in Stored-Product Insects. Pestic. Sci. 1997, 49, 213–228. [Google Scholar] [CrossRef]
- Benhalima, H.; Chaudhry, M.; Mills, K.; Price, N. Phosphine resistance in stored-product insects collected from various grain storage facilities in Morocco. J. Stored Prod. Res. 2004, 40, 241–249. [Google Scholar] [CrossRef]
- Pimentel, M.A.; Faroni, L.R.; Batista, M.D.; Silva, F.H. Resistance of stored-product insects to phosphine. Pesqui. Agropecu. Bras. 2008, 43, 1671–1676. [Google Scholar] [CrossRef]
- Price, N. Active exclusion of phosphine as a mechanism of resistance in Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). J. Stored Prod. Res. 1984, 20, 163–168. [Google Scholar] [CrossRef]
- Schlipalius, D.I.; Valmas, N.; Tuck, A.G.; Jagadeesan, R.; Ma, L.; Kaur, R.; Goldinger, A.; Anderson, C.; Kuang, J.; Zuryn, S. A core metabolic enzyme mediates resistance to phosphine gas. Science 2012, 338, 807–810. [Google Scholar] [CrossRef]
- Alnajim, I.; Agarwal, M.; Liu, T.; Du, X.; Ren, Y.L. Preliminary Study on the Differences in Hydrocarbons Between Phosphine-Susceptible and-Resistant Strains of Rhyzopertha dominica (Fabricius) and Tribolium castaneum (Herbst) Using Direct Immersion Solid-Phase Microextraction Coupled with GC-MS. Molecules 2020, 25, 1565. [Google Scholar] [CrossRef]
- Patel, M.S.; Roche, T.E. Molecular biology and biochemistry of pyruvate dehydrogenase complexes 1. FASEB J. 1990, 4, 3224–3233. [Google Scholar] [CrossRef]
- Williams, C.H. Lipoamide dehydrogenase, glutathione reductase, thioredoxin reductase, and mercuric ion reductase-a family of flavoenzyme transhydrogenases. Chem. Biochem. Flavoenzymes 1992, 3, 121–211. [Google Scholar]
- Schlipalius, D.I.; Tuck, A.G.; Jagadeesan, R.; Nguyen, T.; Kaur, R.; Subramanian, S.; Barrero, R.; Nayak, M.; Ebert, P.R. Variant linkage analysis using de novo transcriptome sequencing identifies a conserved phosphine resistance gene in insects. Genetics 2018, 209, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahara, E.B.; Barros, M.H.; Oliveira, G.A.; Netto, L.E.; Kowaltowski, A.J. Dihydrolipoyl dehydrogenase as a source of reactive oxygen species inhibited by caloric restriction and involved in Saccharomyces cerevisiae aging. FASEB J. 2007, 21, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Pino Moreno, J.; Ganguly, A. Determination of fatty acid content in some edible insects of Mexico. J. Insects Food Feed. 2016, 2, 37–42. [Google Scholar] [CrossRef]
- Gilbert, L.I.; Chino, H. Transport of lipids in insects. J. Lipid Res. 1974, 15, 439–456. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Meeting the energetic demands of insect diapause: Nutrient storage and utilisation. J. Insect Physiol. 2007, 53, 760–773. [Google Scholar] [CrossRef]
- Ziegler, R.; Van Antwerpen, R. Lipid uptake by insect oocytes. Insect Biochem. Mol. Biol. 2006, 36, 264–272. [Google Scholar] [CrossRef]
- Beenakkers, A.T.; Van der Horst, D.; Van Marrewijk, W. Insect flight muscle metabolism. Insect Biochem. 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Ad, M.T.; Van der Horst, D.J.; Van Marrewijk, W.J. Insect lipids and lipoproteins, and their role in physiological processes. Prog. Lipid Res. 1985, 24, 19–67. [Google Scholar]
- Downer, R.; Matthews, J. Patterns of lipid distribution and utilisation in insects. Am. Zool. 1976, 16, 733–745. [Google Scholar] [CrossRef]
- Chang, F. Effects of vertebrate adipokinetic hormones on the rate of in vitro lipid release in insects. Comp. Biochem. Physiol. B Biochem. Mol. Biol. Comp. Biochem. B 1974, 49, 567–578. [Google Scholar] [CrossRef]
- Arrese, E.L.; Wells, M.A. Adipokinetic hormone-induced lipolysis in the fat body of an insect, Manduca sexta: Synthesis of sn-1, 2-diacylglycerols. J. Lipid Res. 1997, 38, 68–76. [Google Scholar] [CrossRef]
- Jutsum, A.; Goldsworthy, G. Fuels for flight in Locusta. J. Insect Physiol. 1976, 22, 243–249. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Zhang, T.; Wang, C.; Huang, Z.; Luo, X.; Deng, Y. A review on phospholipids and their main applications in drug delivery systems. Asian J. Pharm. Sci. 2015, 10, 81–98. [Google Scholar] [CrossRef]
- Singh, R.P.; Gangadharappa, H.; Mruthunjaya, K. Phospholipids: Unique carriers for drug delivery systems. J. Drug Deliv. Sci. Technol. 2017, 39, 166–179. [Google Scholar] [CrossRef]
- Bohdanowicz, M.; Grinstein, S. Role of phospholipids in endocytosis, phagocytosis, and macropinocytosis. Physiol. Rev. 2013, 93, 69–106. [Google Scholar] [CrossRef]
- Puinean, A.M.; Foster, S.P.; Oliphant, L.; Denholm, I.; Field, L.M.; Millar, N.S.; Williamson, M.S.; Bass, C. Amplification of a cytochrome P450 gene is associated with resistance to neonicotinoid insecticides in the aphid Myzus persicae. PLoS Genet. 2010, 6, e1000999. [Google Scholar] [CrossRef]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juárez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A. Cytochrome P450 associated with insecticide resistance catalyses cuticular hydrocarbon production in Anopheles gambiae. Proc. Natio. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef]
- Pedrini, N.; Mijailovsky, S.J.; Girotti, J.R.; Stariolo, R.; Cardozo, R.M.; Gentile, A.; Juárez, M.P. Control of pyrethroid-resistant Chagas disease vectors with entomopathogenic fungi. PLoS Negl. Trop. Dis. 2009, 3, e434. [Google Scholar] [CrossRef]
- Nath, N.S.; Bhattacharya, I.; Tuck, A.G.; Schlipalius, D.I.; Ebert, P.R. Mechanisms of phosphine toxicity. J. Toxicol. 2011, 2011, 494168. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.A.; Faroni, L.R.; Tótola, M.R.; Guedes, R.N. Phosphine resistance, respiration rate and fitness consequences in stored product insects. Pest Manag. Sci. Former. Pestic. Sci. 2008, 63, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T. Genetics of Phosphine Resistance in the Rice Weevil Sitophilus oryzae (L.) (Coleoptera: Curculionidae). Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2016; p. 151. [Google Scholar]
- FAO. Recommended methods for the detection and measurement of resistance of agricultural pests to pesticides. Tentative method for adults of some major pest species of stored cereals, with methyl bromide and phosphine. FAO Method No. 16. FAO Plant Prot. Bull. 1975, 23, 12–25. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protoc. Bioinform. 2016, 55, 1–14. [Google Scholar] [CrossRef]
- Strycharz, J.P.; Lao, A.; Li, H.; Qiu, X.; Lee, S.H.; Sun, W.; Yoon, K.S.; Doherty, J.J.; Pittendrigh, B.R.; Clark, J.M. Resistance in the highly DDT-resistant 91-R strain of Drosophila melanogaster involves decreased penetration, increased metabolism, and direct excretion. Pestic. Biochem. Physiol. 2013, 107, 207–217. [Google Scholar] [CrossRef]
- Ziegler, R. Changes in lipid and carbohydrate metabolism during starvation in adult Manduca sexta. J. Comp. Physiol. B 1991, 161, 125–131. [Google Scholar] [CrossRef]
- Dua, R.; Sunkaria, A.; Kumar, V.; Gill, K.D. Impaired mitochondrial energy metabolism and kinetic properties of cytochrome oxidase following acute aluminium phosphide exposure in rat liver. Food Chem. Toxicol. 2010, 48, 53–60. [Google Scholar] [CrossRef]
- Price, NR Dance, SJ Some biochemical aspects of phosphine action and resistance in three species of stored product beetles. Comp. Biochem. Physiol. B Comp. Pharmacol. Toxicol. 1983, 76, 277–281. [CrossRef]
- Price, N. Some aspects of the inhibition of cytochrome c oxidase by phosphine in susceptible and resistant strains of Rhyzopertha dominicia. Insect Biochem. 1980, 10, 147–150. [Google Scholar] [CrossRef]
- Zuryn, S.; Kuang, J.; Ebert, P. Mitochondrial modulation of phosphine toxicity and resistance in Caenorhabditis elegans. Toxicol. Sci. 2008, 102, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, M.Q.; Price, N.R. Insect mortality at doses of phosphine which produce equal uptake in susceptible and resistant strains of Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). J. Stored Prod. Res. 1990, 26, 101–107. [Google Scholar] [CrossRef]
- Steele, J. Glycogen phosphorylase in insects. Insect Biochem. 1982, 12, 131–147. [Google Scholar] [CrossRef]
- Athenstaedt, K.; Daum, G. The life cycle of neutral lipids: Synthesis, storage and degradation. Cell. Mol. Life Sci. 2006, 63, 1355–1369. [Google Scholar] [CrossRef] [PubMed]
- McDougall, G.; Steele, J. Free fatty acids as a source of energy for trehalose synthesis in the fat body of the American cockroach (Periplaneta americana). Insect Biochem. 1988, 18, 591–597. [Google Scholar] [CrossRef]
- Gäde, G.; Auerswald, L. Beetles’ choice proline for energy output: Control by AKHs. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 132, 117–129. [Google Scholar] [CrossRef]
- Tsuchida, K.; Arai, M.; Tanaka, Y.; Ishihara, R.; Ryan, R.O.; Maekawa, H. Lipid transfer particle catalyses transfer of carotenoids between lipophorins of Bombyx mori. Insect Biochem. Mol. Biol. 1998, 28, 927–934. [Google Scholar] [CrossRef]
- Pepeu, G.; Pepeu, I.M.; Amaducci, L. A review of phosphatidylserine pharmacological and clinical effects. Is phosphatidylserine a drug for the ageing brain? Pharmacol. Res. 1996, 33, 73–80. [Google Scholar] [CrossRef]
- Al-Azzawi, M.J.; Al-Hakkak, Z.S.; Al-Adhami, B.W. In vitro inhibitory effects of phosphine on human and mouse serum cholinesterase. Toxicol. Environ. Chem. 1990, 29, 53–56. [Google Scholar] [CrossRef]
- Hoch, F.L. Cardiolipins and biomembrane function. Biochim. Biophys. Acta Biomembr. 1992, 1113, 71–133. [Google Scholar] [CrossRef]
- Horvath, S.E.; Daum, G. Lipids of mitochondria. Prog. Lipid Res. 2013, 52, 590–614. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.D.; Basu Ball, W.; Pryce, E.N.; Gohil, V.M. Specific requirements of nonbilayer phospholipids in mitochondrial respiratory chain function and formation. J. Mol. Cell Biol. 2016, 27, 2161–2171. [Google Scholar] [CrossRef] [PubMed]
- Price, N.R.; Walter, C.M. A comparison of some effects of phosphine, hydrogen cyanide and anoxia in the lesser grain borer, Rhyzopertha dominica (F.) (Coleoptera: Bostrychidae). Comp. Biochem. Physiol. C Comp. Pharmacol. Toxicol. 1987, 86, 33–36. [Google Scholar] [CrossRef]




{kind=link}
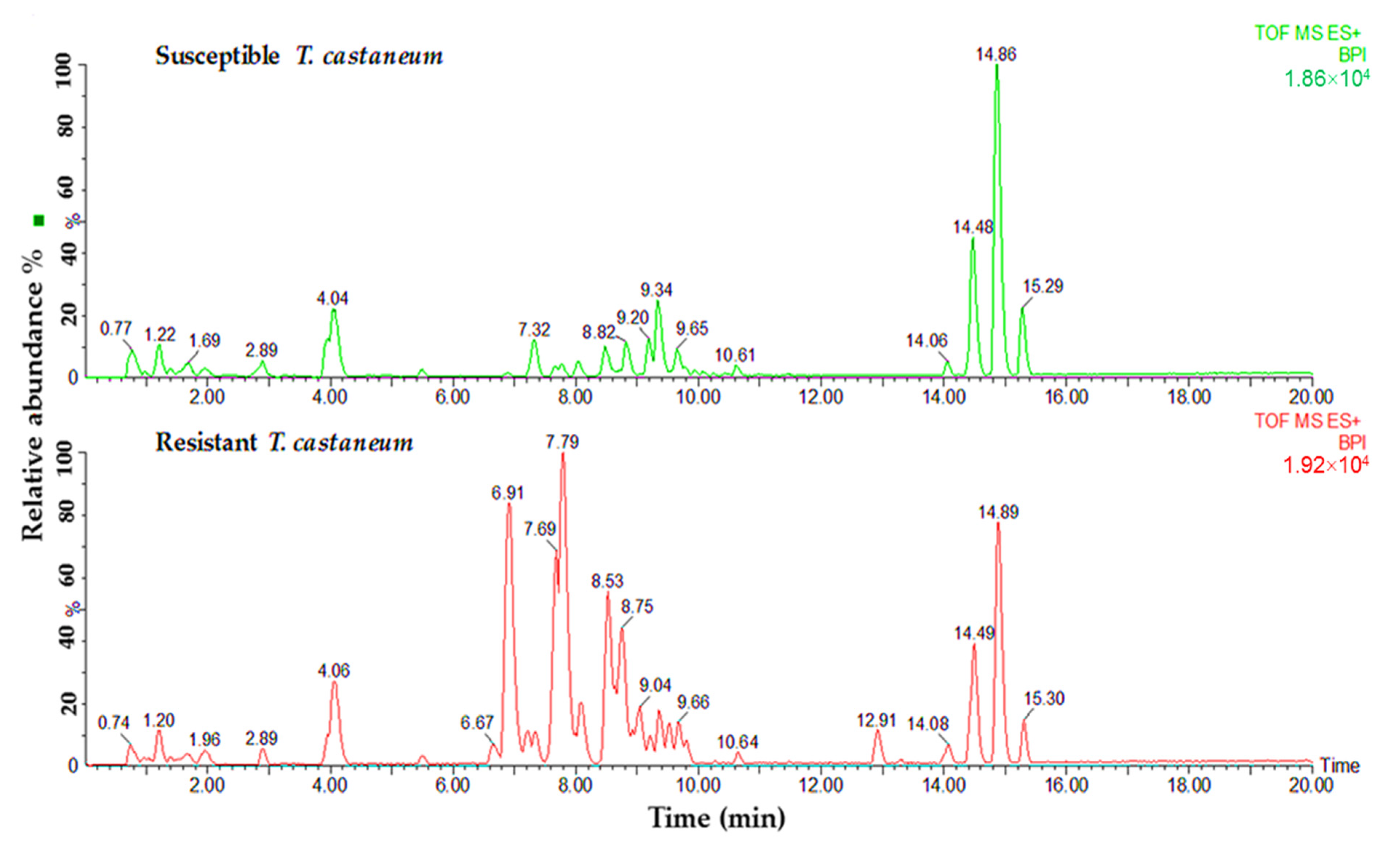{kind=link}
{kind=link}
| No | Lipids ID | Input Mass | Matched Mass | Lipid Category | Name | Ion | LC-MS Response (n = 4) | FC | p Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| Resistant | Susceptible | |||||||||
| 1 | 1.99_369.3 | 369.3532 | 369.3727 | phospholipids | LPA(13:0) | [M+H]+ | 5.02 ± 0.83 | 2.31 ± 1.44 | 2.17 | <0.05 |
| 2 | 1.99_522.3 | 522.2812 | 522.2826 | phospholipids | LPC(18:1) | [M+NH4]+ | 99.91 ± 16.34 | 49.16 ± 7.18 | 2.03 | <0.0005 |
| 3 | 1.99_544.2 | 544.3822 | 544.3609 | phospholipids | LPC(20:4) | [M+NH4]+ | 31.13 ± 4.84 | 43.99 ± 6.80 | 0.70 | ns |
| 4 | 4.04_621.2 | 621.3114 | 621.3034 | phospholipids | LPI(20:4) | [M+H]+ | 832.66 ± 52.05 | 790.97 ± 101.41 | 1.05 | ns |
| 5 | 4.04_666.2 | 666.3628 | 666.3613 | phospholipids | LPI(22:4) | [M+NH4]+ | 68.50 ± 8.76 | 61.30 ± 13.01 | 1.11 | ns |
| 6 | 5.60_457.2 | 457.2211 | 457.2561 | phospholipids | LPG(14:0) | [M+H]+ | 2.00 ± 0.56 | 1.09 ± 0.13 | 1.84 | ns |
| 7 | 5.60_437.3 | 437.3476 | 437.3625 | glycerolipids | MG(24:3) | [M+H]+ | 7.89 ± 0.73 | 4.95 ± 0.90 | 1.59 | ns |
| 8 | 6.88_782.4 | 782.4642 | 782.4896 | glycerolipids | DGDG(23:1) | [M+NH4]+ | 2717.16 ± 44.73 | 54.27 ± 5.57 | 50.10 | <0.0005 |
| 9 | 6.88_804.4 | 804.4523 | 804.474 | glycerolipids | DGDG(25:4) | [M+NH4]+ | 133.56 ± 4.30 | 4.31 ± 1.27 | 31.00 | <0.0005 |
| 10 | 6.88_701.4 | 701.4454 | 701.4259, 701.5715 | glycerolipids | MGDG(31:8), TG(41:4) | [M+H]+ | 74.04 ± 4.77 | 45.81 ± 9.18 | 1.62 | ns |
| 11 | 6.88_601.4 | 601.4694 | 601.4826, 601.4979 | glycerolipids | MGDG(23:2), DG(35:5) | [M+H]+ | 125.56 ± 9.54 | 11.98 ± 1.26 | 10.50 | <0.0005 |
| 12 | 7.20_740.4 | 740.5026 | 740.5307 | glycerolipids | MGDG(32:4) | [M+NH4]+ | 338.85 ± 42.19 | 6.39 ± 0.85 | 53.10 | <0.0005 |
| 13 | 7.27_599.4 | 599.4126 | 599.379, 599.467 | glycerolipids | MGDG(23:3), DG(35:6) | [M+H]+ | 159.43 ± 10.07 | 3.36 ± 0.63 | 47.50 | <0.0005 |
| 14 | 7.22_703.4 | 703.4723 | 703.4416, 703.5871 | glycerolipids | MGDG(31:7), TG(41:3) | [M+H]+ | 273.52 ± 50.17 | 354.10 ± 23.76 | 0.77 | ns |
| 15 | 7.75_784.4 | 784.4761 | 784.5053 | glycerolipids | DGDG(23:0) | [M+NH4]+ | 3137.94 ± 229.11 | 117.26 ± 8.82 | 26.80 | <0.0005 |
| 16 | 7.75_758.4 | 758.4732 | 758.4967, 758.4814 | phospholipids | PG(34:5), PI(27:0) | [M+NH4]+ | 2087.66 ± 76.10 | 104.59 ± 13.47 | 20.00 | <0.0005 |
| 17 | 7.77_806.4 | 806.4587 | 806.4814 | phospholipids | PS(38:7), PC(37:7(OH)) | [M+NH4]+ | 110.65 ± 13.14 | 8.09 ± 0.64 | 13.70 | <0.0005 |
| 18 | 7.77_603.4 | 603.4434 | 603.4103, 603.4983 | glycerolipids | MGDG(23:1), DG(35:4) | [M+H]+ | 313.60 ± 17.51 | 36.05 ± 4.00 | 8.70 | <0.0005 |
| 19 | 8.04_729.4 | 729.4751 | 729.4701, 729.6028 | phospholipids | PE(38:8), PS(34:2) | [M+H]+ | 306.08 ± 6.74 | 176.42 ± 29.97 | 1.74 | ns |
| 20 | 8.03_575.4 | 575.453 | 575.467 | glycerolipids | DG(33:4) | [M+H]+ | 158.93 ± 7.77 | 18.01 ± 3.30 | 8.82 | <0.0005 |
| 21 | 8.48_760.5 | 760.4868 | 760.5123, | phospholipids | PG(34:4) | [M+NH4]+ | 1561.83 ± 22.12 | 172.36 ± 20.44 | 9.06 | <0.0005 |
| 22 | 8.52_731.4 | 731.4872 | 731.5668, 731.6184 | glycerolipids | MGDG(32:0), TG(43:3) | [M+H]+ | 362.89 ± 10.73 | 319.09 ± 50.36 | 1.13 | ns |
| 23 | 8.77_786.5 | 786.4706 | 786.427, 786.5151 | glycerolipids | DGDG(24:6),MGDG(36:9) | [M+NH4]+ | 1827.19 ± 44.90 | 106.02 ± 17.82 | 17.14 | <0.0005 |
| 24 | 8.83_663.3 | 663.3986 | 663.4103, 663.4983 | glycerolipids | MGDG(28:6), DG(40:9) | [M+H]+ | 359.03 ± 23.18 | 296.30 ± 23.71 | 1.21 | ns |
| 25 | 8.77_718.4 | 718.4232 | 718.4525, 718.5017 | phospholipids | PC(30:2(OH)),PE(33:2(OH)) | [M+NH4]+ | 287.33 ± 35.33 | 29.15 ± 6.48 | 9.86 | <0.0005 |
| 26 | 8.83_664.3 | 664.3537 | 664.3456 | phospholipids | LPI(22:5), PS(27:1) | [M+NH4]+ | 154.02 ± 1.65 | 132.65 ± 10.12 | 1.16 | ns |
| 27 | 8.73_577.4 | 577.4341 | 577.3946, 577.4826 | glycerolipids | MGDG(21:0), DG(33:3) | [M+H]+ | 174.22 ± 10.17 | 21.30 ± 2.10 | 8.18 | <0.0005 |
| 28 | 9.03_744.4 | 757.4978 | 757.5824, 757.6341 | glycerolipids | MGDG(34:1), TG(45:4) | [M+H]+ | 704.50 ± 65.61 | 29.31 ± 6.03 | 24.00 | <0.0005 |
| 29 | 9.04_604.4 | 604.4458 | 604.4055, 604.4935 | glycerolipids | MGDG(22:2), DG(34:5) | [M+NH4]+ | 147.95 ± 7.35 | 6.72 ± 1.56 | 22.00 | <0.0005 |
| 30 | 9.37_772.4 | 772.4944 | 772.5933, 772.64 | glycerolipids | MGDG(34:2), TG(45:5) | [M+NH4]+ | 470.23 ± 76.31 | 749.62 ± 135.62 | 0.63 | ns |
| 31 | 9.37_795.4 | 795.4863 | 795.5042, 795.51 | glycerolipids | MGDG(38:10),DGDG(25:0) | [M+H]+ | 14.07 ± 2.71 | 24.61 ± 2.25 | 0.57 | ns |
| 32 | 9.37_762.4 | 762.4745 | 762.5151, 762.427 | glycerolipids | MGDG(34:7), DGDG(22:4) | [M+NH4]+ | 25.54 ± 1.07 | 12.98 ± 0.96 | 1.97 | ns |
| 33 | 9.80_931.5 | 931.5 | 931.5331 | phospholipids | PI(42:10) | [M+H]+ | 103.54 ± 11.13 | 8.10 ± 0.99 | 12.8 | <0.0005 |
| 34 | 9.80_752.5 | 752.50 | 752.5072, 752.5225 | phospholipids | PE(36:6(OH)), PC(34:5) | [M+H]+ | 201.86 ± 18.26 | 17.15 ± 4.22 | 11.80 | <0.005 |
| 35 | 12.93_892.6 | 892.6547 | 892.7389 | glycerolipids | TG(54:8) | [M+NH4]+ | 48.15 ± 9.55 | 9.74 ± 1.50 | 4.95 | <0.0005 |
| 36 | 13.30_895.6 | 895.6443 | 895.681 | glycerolipids | TG(56:12) | [M+H]+ | 41.92 ± 7.31 | 3.86 ± 3.73 | 10.90 | <0.005 |
| 37 | 14.06_900.6 | 900.6378 | 900.7076 | glycerolipids | TG(55:11) | [M+NH4]+ | 101.13 ± 2.30 | 15.72 ± 2.58 | 6.43 | <0.05 |
| 38 | 14.49_874.6 | 874.6416 | 874.6919 | glycerolipids | TG(53:10) | [M+NH4]+ | 1566.44 ± 180.26 | 868.03 ± 114.93 | 1.80 | ns |
| 39 | 14.49_848.6 | 848.6331 | 848.6763 | glycerolipids | TG(51:9) | [M+NH4]+ | 909.28 ± 89.13 | 347.89 ± 51.29 | 2.61 | <0.05 |
| 40 | 14.49_822.6 | 822.6191 | 822.6606 | glycerolipids | TG(49:8) | [M+NH4]+ | 293.52 ± 16.92 | 118.59 ± 17.31 | 2.48 | <0.05 |
| 41 | 14.90_876.6 | 876.6555 | 876.7076 | glycerolipids | TG(53:9) | [M+NH4]+ | 3310.62 ± 139.19 | 2189.69 ± 296.72 | 1.51 | ns |
| 42 | 14.90_850.6 | 850.644 | 850.6919 | glycerolipids | TG(51:8) | [M+NH4]+ | 2272.31 ± 153.12 | 1155.83 ± 164.14 | 1.97 | ns |
| 43 | 14.90_902.6 | 902.6696 | 902.7232 | glycerolipids | TG(55:10) | [M+NH4]+ | 474.79 ± 64.08 | 212.74 ± 22.57 | 2.23 | <0.05 |
| 44 | 15.32_878.6 | 878.6759 | 878.7232 | glycerolipids | TG(53:8) | [M+NH4]+ | 838.12 ± 60.35 | 391.75 ± 57.23 | 2.14 | <0.05 |
| 45 | 15.32_904.6 | 904.6899 | 904.7389 | glycerolipids | TG(55:9) | [M+NH4]+ | 370.22 ± 57.50 | 145.12 ± 24.04 | 2.55 | <0.05 |
| No | Lipids ID | Input Mass | Matched Mass | Lipid Category | Name | Ion | LC-MS response (n = 4) | FC | p Value | |
|---|---|---|---|---|---|---|---|---|---|---|
| Resistant | Susceptible | |||||||||
| 1 | 1.20_333.2 | 331.2029 | 331.2843 | glycerolipids | MG(16:0) | [M+H]+ | 494.20 ± 39.19 | 373.22 ± 30.01 | 1.32 | ns |
| 2 | 1.42_372.2 | 372.2362 | 372.3108 | glycerolipids | MG(18:2) | [M+NH4]+ | 165.21 ± 8.96 | 123.47 ± 11.27 | 1.34 | ns |
| 3 | 1.42_504.3 | 504.3135 | 504.4047 | glycerolipids | MG(28:6) | [M+NH4]+ | 194.33 ± 5.74 | 150.90 ± 5.74 | 1.29 | ns |
| 4 | 1.42_522.3 | 522.3278 | 522.4153 | glycerolipids | DG(28:4) | [M+NH4]+ | 133.57 ± 9.96 | 79.67 ± 15.17 | 1.68 | ns |
| 5 | 1.77_497.3 | 497.3189 | 497.2874 | phospholipids | LPG(17:1) | [M+H]+ | 55.96 ± 3.28 | 79.22 ± 5.13 | 0.71 | ns |
| 6 | 2.85_440.3 | 440.3844 | 440.4462 | phospholipids | LPA (P-18:0) | [M+NH4]+ | 131.61 ± 38.07 | 228.69 ± 74.20 | 0.58 | ns |
| 7 | 2.85_387.2 | 387.2717 | 387.3469 | glycerolipids | MG(20:0) | [M+H]+ | 22.74 ± 2.45 | 26.03 ± 0.93 | 0.87 | ns |
| 8 | 2.85_397.3 | 397.3575 | 397.404 | phospholipids | LPA (O-16:0) | [M+H]+ | 8.03 ± 2.17 | 7.60 ± 2.14 | 1.06 | ns |
| 9 | 2.85_560.3 | 560.3673 | 560.283 | phospholipids | LPI(14:1) | [M+NH4]+ | 22.75 ± 1.48 | 26.00 ± 0.18 | 0.88 | ns |
| 10 | 4.006_621.2 | 621.2667 | 621.3034 | phospholipids | LPI(20:4) | [M+H]+ | 875.73 ± 15.65 | 764.82 ± 134.20 | 1.15 | ns |
| 11 | 4.006_468.4 | 468.4175 | 468.4775 | phospholipids | LPA (P-20:0) | [M+NH4]+ | 147.30 ± 42.49 | 142.10 ± 46.77 | 1.04 | ns |
| 12 | 6.91_782.5 | 782.504 | 782.4896 | glycerolipids | DGDG(23:1), | [M+NH4]+ | 2019.82 ± 235.61 | 68.25 ± 17.86 | 29.60 | <0.0005 |
| 13 | 6.91_701.5 | 701.5061 | 701.5715,701.4259 | glycerolipids | TG(41:4), MGDG(31:8) | [M+H]+ | 48.53 ± 7.57 | 27.33 ± 4.27 | 1.78 | ns |
| 14 | 6.91_805.4 | 805.4935 | 805.4944 | glycerolipids | DGDG(26:2) | [M+H]+ | 48.53 ± 7.57 | 27.33 ± 4.27 | 1.72 | ns |
| 15 | 6.91_601.4 | 601.4752 | 601.4826,601.3946 | glycerolipids | DG(35:5), MGDG(23:2) | [M+H]+ | 122.02 ± 15.25 | 10.84 ± 3.47 | 11.30 | <0.0005 |
| 16 | 7.32_703.5 | 703.5203 | 703.5355,703.5871 | glycerolipids | MGDG(30:0), TG(41:3) | [M+H]+ | 274.50 ± 37.66 | 197.51 ± 29.57 | 1.39 | ns |
| 17 | 7.32_740.4 | 740.4688 | 740.5307 | glycerolipids | MGDG(32:4) | [M+NH4]+ | 200.90 ± 26.49 | 14.97 ± 3.73 | 13.40 | <0.0005 |
| 18 | 7.32_599.4 | 599.4777 | 599.4823, 599.379 | glycerolipids | MGDG(23:3), DG(35:6) | [M+H]+ | 98.52 ± 11.18 | 6.03 ± 0.87 | 16.30 | <0.0005 |
| 19 | 7.32_725.4 | 725.4998 | 725.5198 | glycerolipids | MGDG(32:3) | [M+H]+ | 21.10 ± 2.32 | 16.90 ± 2.12 | 1.25 | ns |
| 20 | 7.73_603.4 | 603.495 | 603.4983,603.4103 | glycerolipids | DG(35:4), MGDG(23:1) | [M+H]+ | 345.30 ± 25.45 | 29.48 ± 8.50 | 11.70 | <0.0005 |
| 21 | 7.73_806.5 | 806.5099 | 806.4814 | phospholipids | PS(38:7), PC(37:7(OH)) | [M+NH4]+ | 100.43 ± 4.92 | 11.88 ± 1.87 | 8.45 | <0.0005 |
| 22 | 8.04_742.4 | 742.4846 | 742.4583,742.5464 | glycerolipids | DGDG(20:0), MGDG(32:3) | [M+NH4]+ | 606.81 ± 28.74 | 224.17 ± 32.49 | 2.71 | <0.05 |
| 23 | 8.04_729.5 | 729.5348 | 729.4701, 729.6028 | phospholipids | PE(38:8), PS(34:2) | [M+H]+ | 244.84 ± 19.95 | 136.24 ± 18.63 | 1.80 | ns |
| 24 | 8.04_716.4 | 716.4632 | 716.4861, 716.4345 | phospholipids | LPG(32:5), PI(24:0) | [M+NH4]+ | 303.01 ± 20.56 | 30.08 ± 6.45 | 10.10 | <0.0005 |
| 25 | 8.04_575.4 | 575.4651 | 575.467 | glycerolipids | DG(33:4) | [M+H]+ | 190.19 ± 23.05 | 19.17 ± 4.16 | 9.92 | <0.0005 |
| 26 | 8.51_760.5 | 760.5245 | 760.5123 | phospholipids | PG(34:4) | [M+NH4]+ | 1675.19 ± 107.08 | 278.83 ± 55.74 | 6.01 | <0.0005 |
| 27 | 8.51_731.5 | 731.5506 | 731.5668, 731.6184 | glycerolipids | MGDG(32:0), TG(43:3) | [M+H]+ | 436.03 ± 22.29 | 340.56 ± 46.97 | 1.28 | ns |
| 28 | 8.51_753.5 | 753.5414 | 753.5511, 753.6028 | glycerolipids | MGDG(34:3), TG(45:6), | [M+H]+ | 40.47 ± 1.43 | 42.42 ± 6.82 | 0.95 | ns |
| 29 | 8.51_744.5 | 744.5378 | 744.481, 744.4658, | phospholipids | PG(33:5), PI(26:0) | [M+NH4]+ | 206.92 ± 21.10 | 124.11 ± 17.26 | 1.67 | ns |
| 30 | 8.78_718.4 | 718.4864 | 718.4525, 718.5017 | phospholipids | PC(30:2(OH), PE(33:2(OH)) | [M+NH4]+ | 423.19 ± 23.43 | 80.76 ± 16.35 | 5.24 | <0.0005 |
| 31 | 8.78_786.5 | 786.5374 | 786.427, 786.5151 | glycerolipids | DGDG(24:6), MGDG(36:9) | [M+NH4]+ | 1252.82 ± 103.59 | 142.53 ± 25.80 | 8.79 | <0.0005 |
| 32 | 8.78_577.4 | 577.4322 | 577.3946, 577.4826 | glycerolipids | MGDG(21:0), DG(33:3) | [M+H]+ | 270.85 ± 30.97 | 55.24 ± 11.17 | 4.90 | <0.0005 |
| 33 | 8.78_663.4 | 663.4093 | 663.4103, 663.4983 | glycerolipids | MGDG(28:6), DG(40:9) | [M+H]+ | 609.72 ± 74.05 | 507.66 ± 27.32 | 1.20 | ns |
| 34 | 9.01_766.4 | 766.4847 | 766.4583, 766.5464 | glycerolipids | DGDG(22:2),MGDG(34:5) | [M+NH4]+ | 71.82 ± 4.59 | 31.68 ± 3.10 | 2.27 | <0.05 |
| 35 | 9.01_757.5 | 757.5558 | 757.5824, 757.6341 | glycerolipids | MGDG(34:1), TG(45:4) | [M+H]+ | 51.64 ± 2.38 | 55.87 ± 6.82 | 0.92 | ns |
| 36 | 9.45_788.5 | 788.5247 | 788.5307, 788.6187 | glycerolipids | MGDG(36:8), DG(48:11) | [M+NH4]+ | 201.30 ± 24.97 | 43.00 ± 8.75 | 4.68 | <0.0005 |
| 37 | 9.45_772.5 | 772.5918 | 772.5933, 772.64 | glycerolipids | MGDG(34:2), TG(45:5) | [M+NH4]+ | 99.85 ± 5.59 | 84.51 ± 9.65 | 1.18 | ns |
| 38 | 9.45_759.5 | 759.5811 | 759.5981, 759.6497 | glycerolipids | MGDG(34:0), TG(45:3) | [M+H]+ | 58.75 ± 1.45 | 45.27 ± 5.51 | 1.30 | ns |
| 39 | 9.45_728.5 | 728.5168 | 728.5307, 728.5824 | glycerolipids | MGDG(31:3), TG(42:6) | [M+NH4]+ | 31.43 ± 4.19 | 14.37 ± 1.82 | 2.19 | <0.05 |
| 40 | 9.45_702.4 | 702.4594 | 702.4341, 702.4188 | phospholipids | PG(30:5), PI(23:0) | [M+NH4]+ | 84.14 ± 7.79 | 41.91 ± 2.24 | 2.00 | ns |
| 41 | 9.68_730.5 | 730.5211 | 730.4654 | phospholipids | PG(32:5) | [M+NH4]+ | 243.07 ± 13.67 | 182.51 ± 19.88 | 1.33 | ns |
| 42 | 9.68_752.5 | 752.5024 | 752.5072, 752.5225 | phospholipids | PE(36:6(OH), PC(34:5) | [M+H]+ | 39.41 ± 3.15 | 33.47 ± 3.80 | 1.18 | ns |
| 43 | 9.68_641.4 | 641.4714 | 641.5139, 641.4259 | glycerolipids | DG(38:6), MGDG(26:3) | [M+H]+ | 14.66 ± 1.26 | 7.88 ± 1.39 | 1.86 | ns |
| 44 | 10.59_566.5 | 566.5204 | 566.4779 | glycerolipids | DG(31:3) | [M+NH4]+ | 15.23 ± 1.20 | 18.96 ± 3.34 | 0.8 | ns |
| 45 | 10.59_588.4 | 588.4946 | 588.4986 | glycerolipids | MG(34:6) | [M+NH4]+ | 1.74 ± 0.39 | 2.61 ± 0.74 | 0.67 | ns |
| 46 | 12.75_600.4 | 600.48 | 600.5198 | glycerolipids | TG(32:0) | [M+NH4]+ | 17.54 ± 2.35 | 0.56 ± 0.07 | 31.5 | <0.0005 |
| 47 | 13.63_896.7 | 896.7049 | 896.7702 | glycerolipids | TG(54:6) | [M+NH4]+ | 175.31 ± 23.13 | 12.12 ± 2.46 | 14.5 | <0.0005 |
| 48 | 13.63_870.6 | 870.6881 | 870.7545 | glycerolipids | TG(52:5) | [M+NH4]+ | 67.03 ± 5.73 | 9.21 ± 2.25 | 7.28 | <0.0005 |
| 49 | 13.63_924.7 | 924.7244 | 924.8015 | glycerolipids | TG(56:6) | [M+NH4]+ | 8.99 ± 1.70 | 0.88 ± 0.21 | 10.2 | <0.0005 |
| 50 | 14.03_872.7 | 872.702 | 872.7702 | glycerolipids | TG(52:4) | [M+NH4]+ | 729.15 ± 64.44 | 131.25 ± 17.75 | 5.56 | <0.0005 |
| 51 | 14.03_898.7 | 898.7174 | 898.7858 | glycerolipids | TG(54:5) | [M+NH4]+ | 406.46 ± 40.88 | 51.77 ± 9.89 | 7.85 | <0.0005 |
| 52 | 14.03_877.6 | 877.6541 | 877.728 | glycerolipids | TG(54:7) | [M+H]+ | 80.83 ± 4.60 | 25.55 ± 2.90 | 3.16 | <0.005 |
| 53 | 14.03_846.6 | 846.6855 | 846.7545 | glycerolipids | TG(50:3) | [M+NH4]+ | 78.61 ± 4.63 | 36.13 ± 0.66 | 2.18 | <0.05 |
| 54 | 14.03_900.7 | 900.7314 | 900.7076 | glycerolipids | TG(55:11) | [M+NH4]+ | 107.36 ± 12.25 | 16.55 ± 3.90 | 6.49 | <0.0005 |
| 55 | 14.46_874.7 | 874.7136 | 874.6919 | glycerolipids | TG(53:10) | [M+NH4]+ | 2201.20 ± 47.19 | 1105.18 ± 102.89 | 2.00 | ns |
| 56 | 14.46_848.7 | 848.704 | 848.6763 | glycerolipids | TG(51:9) | [M+NH4]+ | 364.11 ± 10.02 | 335.58 ± 11.22 | 3.15 | <0.005 |
| 57 | 14.46_880.6 | 880.6754 | 880.7389 | glycerolipids | TG(53:7) | [M+NH4]+ | 90.99 ± 4.68 | 59.83 ± 3.47 | 1.52 | ns |
| 58 | 14.85_876.7 | 876.7307 | 876.7076 | glycerolipids | TG(53:9) | [M+NH4]+ | 1030.29 ± 120.17 | 1355.54 ± 62.69 | 0.76 | ns |
| 59 | 14.85_850.7 | 850.7169 | 850.6919 | glycerolipids | TG(51:8) | [M+NH4]+ | 66.44 ± 2.03 | 63.60 ± 3.82 | 1.04 | ns |
| 60 | 14.85_902.7 | 902.7433 | 902.7232 | glycerolipids | TG(55:10) | [M+NH4]+ | 327.72 ± 35.09 | 207.45 ± 23.05 | 1.58 | ns |
| 61 | 14.85_881.6 | 881.699 | 881.7593 | glycerolipids | TG(54:5) | [M+H]+ | 110.66 ± 3.85 | 117.84 ± 10.57 | 0.94 | ns |
| 62 | 14.85_855.6 | 855.6716 | 855.7436 | glycerolipids | TG(52:4) | [M+H]+ | 56.90 ± 4.62 | 62.22 ± 2.34 | 0.91 | ns |
| 63 | 15.26_878.7 | 878.7495 | 878.7232 | glycerolipids | TG(53:8) | [M+NH4]+ | 297.63 ± 42.81 | 239.60 ± 19.02 | 1.24 | ns |
| 64 | 15.26_905.7 | 905.7624 | 905.7593 | glycerolipids | TG(56:7) | [M+H]+ | 83.15 ± 9.77 | 53.37 ± 4.27 | 1.56 | ns |
| 65 | 15.26_883.7 | 883.7335 | 883.681 | glycerolipids | TG(55:11) | [M+H]+ | 20.88 ± 1.10 | 16.90 ± 0.51 | 1.24 | ns |
| 66 | 15.26_909.6 | 909.6959 | 909.7906 | glycerolipids | TG(56:5) | [M+H]+ | 8.24 ± 2.20 | 5.48 ± 0.14 | 1.5 | ns |
| 67 | 15.26_852.7 | 852.7589 | 852.7076 | glycerolipids | TG(51:7) | [M+NH4]+ | 14.22 ± 1.03 | 9.74 ± 0.58 | 1.46 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alnajim, I.; Aldosary, N.; Agarwal, M.; Liu, T.; Du, X.; Ren, Y. Role of Lipids in Phosphine Resistant Stored-Grain Insect Pests Tribolium castaneum and Rhyzopertha dominica. Insects 2022, 13, 798. https://doi.org/10.3390/insects13090798
Alnajim I, Aldosary N, Agarwal M, Liu T, Du X, Ren Y. Role of Lipids in Phosphine Resistant Stored-Grain Insect Pests Tribolium castaneum and Rhyzopertha dominica. Insects. 2022; 13(9):798. https://doi.org/10.3390/insects13090798
Chicago/Turabian StyleAlnajim, Ihab, Naser Aldosary, Manjree Agarwal, Tao Liu, Xin Du, and Yonglin Ren. 2022. "Role of Lipids in Phosphine Resistant Stored-Grain Insect Pests Tribolium castaneum and Rhyzopertha dominica" Insects 13, no. 9: 798. https://doi.org/10.3390/insects13090798
APA StyleAlnajim, I., Aldosary, N., Agarwal, M., Liu, T., Du, X., & Ren, Y. (2022). Role of Lipids in Phosphine Resistant Stored-Grain Insect Pests Tribolium castaneum and Rhyzopertha dominica. Insects, 13(9), 798. https://doi.org/10.3390/insects13090798