
Tomato Varieties Influence the Performance of Tamarixia triozae (Hymenoptera: Eulophidae) on Bactericera cockerelli (Hemiptera: Triozidae) Nymphs

 , , , and
, , , and 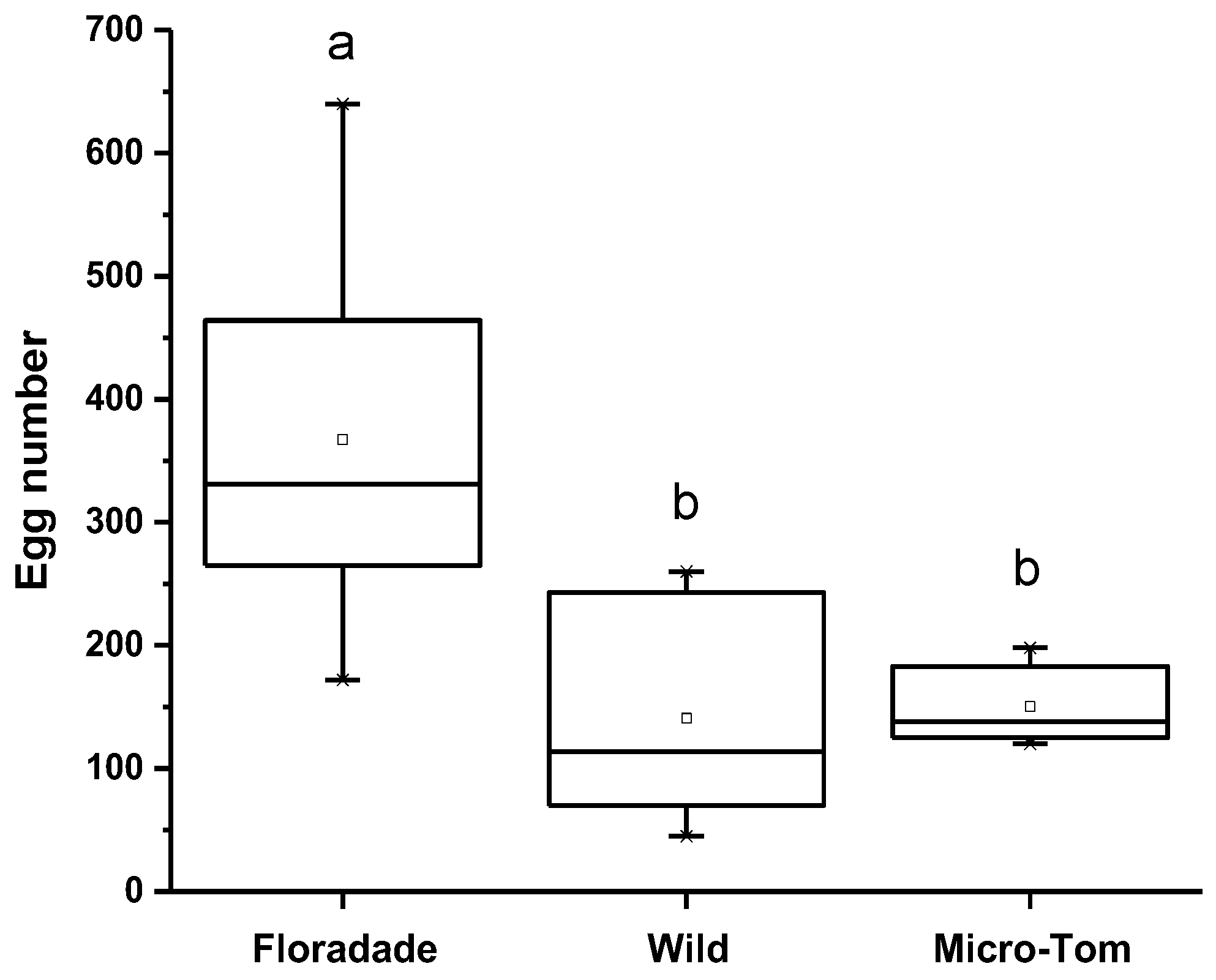{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Growth and Insect Colony
2.2. Parasitism Preference Assay
2.3. Olfactometer Assay
2.4. Statistical Analysis
3. Results
3.1. Parasitism Preference Assay
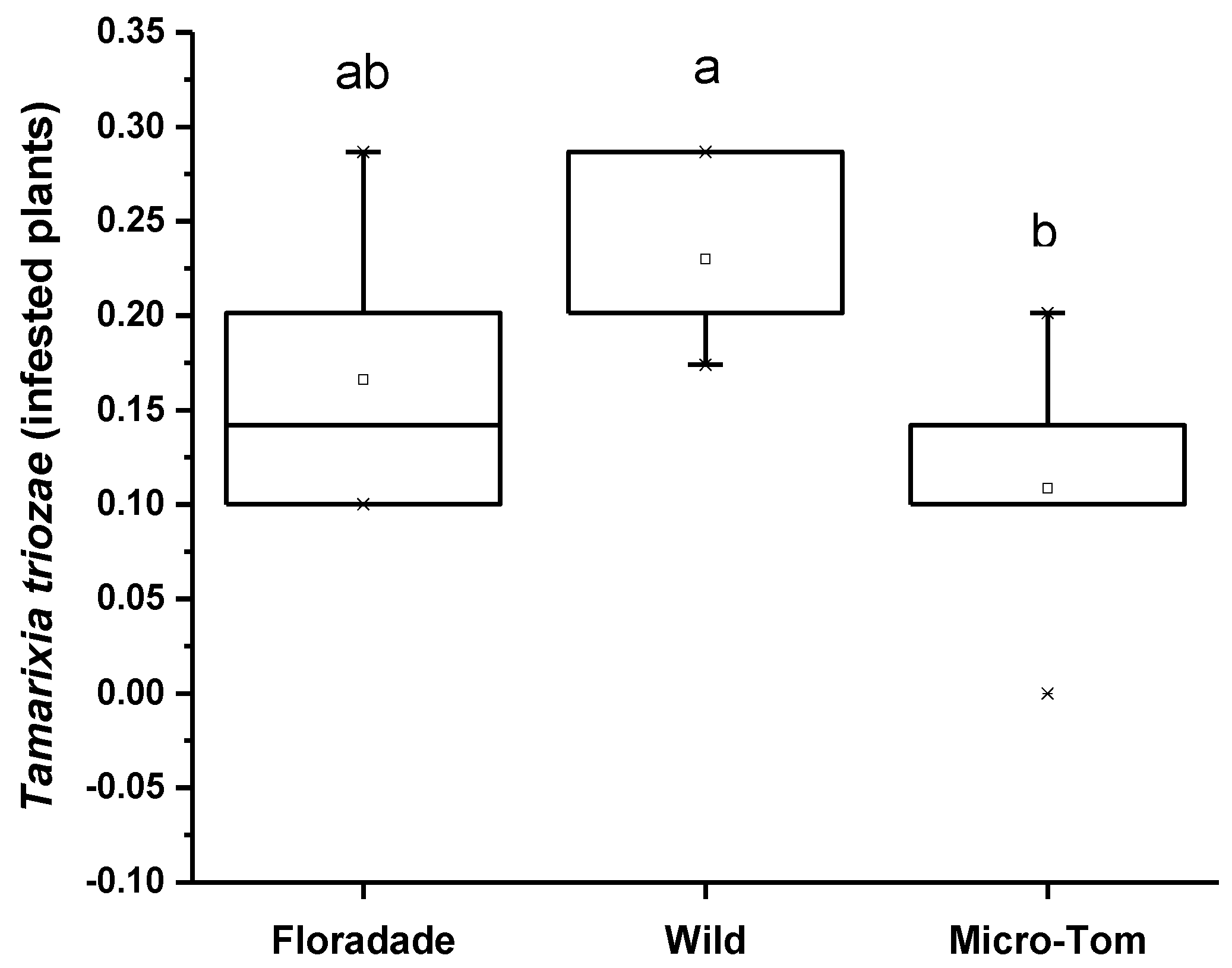
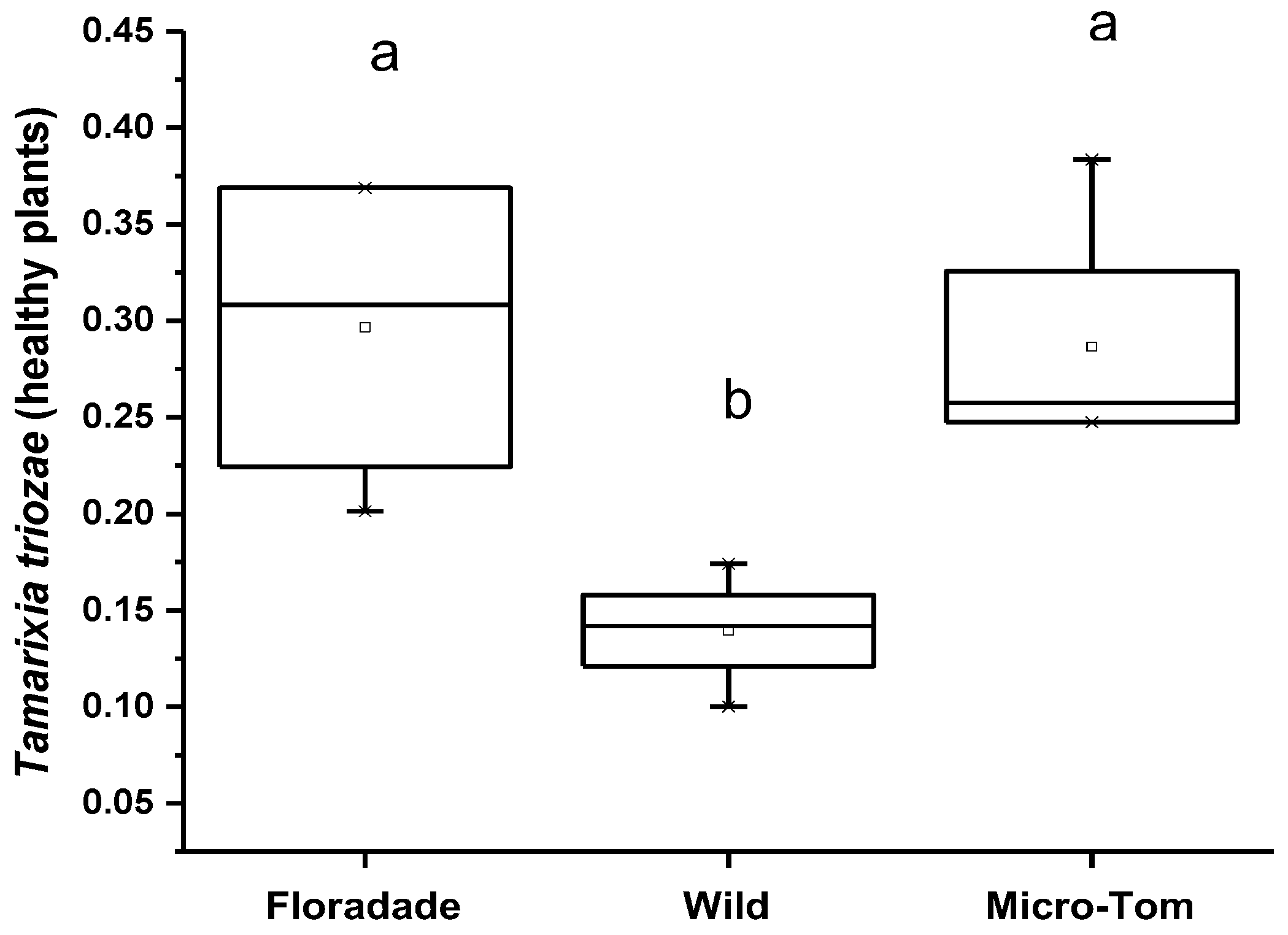
3.2. Olfactometer Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perilla-Henao, L.M.; Casteel, C.L. Vector-Borne Bacterial Plant Pathogens: Interactions with Hemipteran Insects and Plants. Front. Plant Sci. 2016, 7, 1163. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus. Viruses 2020, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, G.; Kavalappara, S.R.; Gautam, S.; Silva, A.d.; Simmons, A.; Srinivasan, R.; Dutta, B. Field Screen and Genotyping of Phaseolus vulgaris against Two Begomoviruses in Georgia, USA. Insects 2021, 12, 49. [Google Scholar] [CrossRef] [PubMed]
- Marchant, W.G.; Gautam, S.; Dutta, B.; Srinivasan, R. Whitefly-Mediated Transmission and Subsequent Acquisition of Highly Similar and Naturally Occurring Tomato Yellow Leaf Curl Virus Variants. Phytopathology 2022, 112, 720–728. [Google Scholar] [CrossRef] [PubMed]
- LaTora, A.G.; Lai, P.C.; Chen, Y.J.; Gautam, S.; Abney, M.R.; Srinivasan, R. Frankliniella fusca (Thysanoptera: Thripidae), The Vector of Tomato Spotted Wilt Orthotospovirus Infecting Peanut in the Southeastern United States. J. Integr. Pest Manag. 2022, 13, 3. [Google Scholar] [CrossRef]
- Butler, C.D.; Trumble, J.T. The potato psyllid, Bactericera cockerelli (Sulc) (Hemiptera: Triozidae): Life history, relationship to plant diseases, and management strategies. Terr. Arthropod Rev. 2012, 5, 87–111. [Google Scholar] [CrossRef]
- Hansen, A.H.K.; Trumble, J.T.; Stouthamer, R.; Paine, T.D. A New Huanglongbing species, Candidatus Liberibacter psyllaurous, found to infect tomato and potato, is vectored by the psyllid Bactericera cockerelli (Sulc). AEM 2008, 74, 5862–5865. [Google Scholar] [CrossRef]
- Liefting, L.W.; Sutherland, P.W.; Ward, L.I.; Paice, K.L.; Weir, B.S. A New “Candidatus Liberibacter” species associated with diseases of solanaceous crops. Plant Dis. 2009, 93, 208–214. [Google Scholar] [CrossRef]
- CNAS. Economic Impacts of Zebra Chip on the Texas Potato Industry. Center for North American Studies. 2006. Available online: http://cnas.tamu.edu/zebra%20chip%20impacts%20final.pdf (accessed on 6 September 2019).
- Wen, A.; Mallik, I.; Alvarado, V.Y.; Pasche, J.S.; Wang, X.; Lin, H.; Scholthof, H.B.; Mirkov, T.E.; Rush, C.M.; Gudmestad, N.C. Detection, distribution, and genetic variability of ‘Candidatus Liberibacter’ species associated with Zebra complex disease of potato in North America. Plant Dis. 2009, 93, 1102–1115. [Google Scholar] [CrossRef]
- Munyaneza, J.E. Zebra Chip disease of potato: Biology, Epidemiology, and Management. Am. J. Potato Res. 2012, 89, 329–350. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.B.; Zhang, Y.M.; Hua, L.; Peng, L.N.; Munyaneza, J.E.; Trumble, J.T.; Liu, T.X. Repellency of selected biorational insecticides to potato psyllid, Bactericera cockerelli (Hemiptera: Psyllidae). Crop Prot. 2010, 29, 1320–1324. [Google Scholar] [CrossRef]
- Granados-Echegoyen, C.; Pérez-Pacheco, R.; Bautista-Martínez, N.; Alonso-Hernández, N.; Sánchez-García, J.A.; Martinez-Tomas, S.H.; Sánchez-Mendoza, S. Insecticidal Effect of Botanical Extracts on Developmental Stages of Bactericera cockerelli (Sulc) (Hemiptera: Triozidae). Southwest. Entomol. 2015, 40, 97–110. [Google Scholar] [CrossRef]
- Landero-Valenzuela, N.; Alonso-Hernández, N.; Lara-Viveros, F.; Gómez-Domínguez, N.S.; Juárez-Pelcastre, J.; Aguado-Rodríguez, J.; Luna-Cruz, A.; Lagunez-Rivera, L.; Aguilar-Pérez, L.A.; Hinojosa-Garro, D.; et al. Efficiency of Schinus molle Essential Oil against Bactericera cockerelli (Hemiptera: Triozidae) and Sitophilus zeamais (Coleoptera: Dryophthoridae). Agriculture 2022, 12, 554. [Google Scholar] [CrossRef]
- Morales, A.S.I.; Martínez, A.M.; Figueroa, J.I.; Espino, H.A.M.; Chavarrieta, Y.J.M.; Ortíz, R.R.; Rodríguez, E.C.L.; Pineda, S. Parámetros de vida del parasitoide sinovigénico Tamarixia triozae (Hymenoptera: Eulophidae). Rev. Colomb. Entomol. 2013, 39, 243–249. [Google Scholar] [CrossRef]
- Rojas, P.; Rodríguez-Leyva, E.; Lomelí-Flores, J.R.; Liu, T.X. Biology and life history of Tamarixia triozae, a parasitoid of the potato psyllid Bactericera cockerelli. BioControl 2015, 60, 27–35. [Google Scholar] [CrossRef]
- Luna-Cruz, A.; Rodríguez-Leyva, E.; Lomelí-Flores, J.R.; Ortega-Arenas, L.D.; Bautista-Martínez, N.; Pineda, S. Toxicity and Residual Activity of Insecticides against Tamarixia triozae (Hymenoptera: Eulophidae), a Parasitoid of Bactericera cockerelli (Hemiptera: Triozidae). J. Econ. Entomol. 2015, 108, 2289–2295. [Google Scholar] [CrossRef]
- Cerón-González, C.; Lomelí-Flores, J.R.; Rodríguez-Leyva, E.; Torres-Ruíz, A. Fecundidad y alimentación de Tamarixia triozae (Hymenoptera: Eulophidae) sobre el psílido de la papa Bactericera cockerelli. Rev. Mex. De Cienc. Agrícolas 2014, 5, 893–899. [Google Scholar] [CrossRef]
- Bravo, M.E.; López, L.P. Principales plagas del chile de agua en los Valles centrales de Oaxaca. Agroproduce Fund. Prod. Oaxaca AC 2007, 7, 12–15. [Google Scholar]
- Liu, T.X.; Yong-Mei, Z.; Li-Nian, P.; Rojas, P.; Trumble, J.T. Risk assessment of selected insecticides on Tamarixia triozae (Hymenoptera: Eulophidae), a parasitoid of Bactericera cockerelli (Hemiptera: Trizoidae). J. Econ. Entomol. 2012, 5, 490–496. [Google Scholar] [CrossRef]
- Trujillo-García, J.; Lozano-Gutiérrez, J.F.; España-Luna, M.P.; Lara-Herrera, A.; Balleza-Cadengo, J.J. Parasitismo de Bactericera cockerelli (Sulc) (Hemiptera: Psyllidae) en dos cultivares de chile guajillo en invernadero. Entomol. Mex. 2018, 5, 160–163. [Google Scholar]
- Butler, C.D.; Trumble, J.T. New records of hyperparasitism of Tamarixia triozae (Burks) (Hymenoptera: Eulophidae) by Encarsia spp. (Hymenoptera: Aphelinidae) in California. Pan-Pac. Entomol. 2011, 87, 130–133. [Google Scholar] [CrossRef]
- Halitschke, R.; Stenberg, J.A.; Kessler, D.; Kessler, A.; Baldwin, I.T. Shared signals—‘alarm calls’ from plants increase apparency to herbivores and their enemies in nature. Ecol. Lett. 2008, 11, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Garvey, M.; Kaplan, I.; Li, B.; Carrillo, J. Domestication of tomato has reduced the attraction of herbivore natural enemies to pest-damaged plants. Agric. For. Entomol. 2017, 20, 390–401. [Google Scholar] [CrossRef]
- Hernández-Moreno, S.; Pérez-Panduro, A.; Lomelí-Flores, J.R.; Rodríguez-Leyva, E.; Bueno-Aguilar, G.; Cibrián-Tovar, J. Aptitud Biológica de Tamarixia triozae Mediada por la Planta Hospedera. Southwest. Entomol. 2017, 42, 225–236. [Google Scholar] [CrossRef]
- Steiner, A.A. The universal nutrient solution. In Proceedings of the 6th International Congress on Soilless Culture, Lunteren, The Netherlands, 29 April–5 May 1984. [Google Scholar]
- Hernández Moreno, S. Biología y Respuesta Comportamental de Tamarixia triozae (Hymenoptera: Eulphidae) Sobre Bactericera cockerelli (Hemiptera: Triozidae) en Jitomate y Chile. Ph.D. Thesis, Colegio de Postgraduados, Texcoco, México, 2016. [Google Scholar]
- Díaz-Montano, J.; Trumble, J.T. Behavioral Responses of the Potato Psyllid (Hemiptera: Triozidae) to Volatiles from Dimethyl Disulfide and Plant Essential Oils. J. Insect Behav. 2012, 26, 336–351. [Google Scholar] [CrossRef]
- Koschier, E.H.; de Kogel, W.J.; Visser, J.H. Assessing the attractiveness of volatile plant compounds to western flower thrips Frankliniella occidentalis. J. Chem. Ecol. 2000, 26, 2643–2655. [Google Scholar] [CrossRef]
- Davidson, M.M.; Perry, N.B.; Larsen, L.; Green, V.C.; Butler, R.C.; Teulon, D.A.J. 4-Pyridyl Carbonyl compounds as thrips lures: Effectiveness for Western flower thrips in Y-tube bioassays. J. Agric. Food Chem. 2008, 56, 6554–6561. [Google Scholar] [CrossRef]
- Williams III, L.; Cheng, Z.Y.; Snodgrass, L.G.; Manrique, V. Plant-mediated decisions by an herbivore affect oviposition pattern and subsequent egg parasitism. Arthropod-Plant Interact. 2012, 6, 159–169. [Google Scholar] [CrossRef]
- Mayo-Hernández, J.; Ramírez-Chávez, E.; Molina-Torres, J.; Guillén-Cisneros, M.L.; Rodríguez-Herrera, R.; Hernández-Castillo, F.; Flores-Olivas, A.; Valenzuela-Soto, J.H. Effects of Bactericera Cockerelli Herbivory on Volatile Emissions of Three Varieties of Solanum Lycopersicum. Plants 2019, 8, 509. [Google Scholar] [CrossRef] [Green Version]
- Salas-Araiza, M.D.; González-Márquez, M.A.; Martínez-Jaime, O.A.; Guzmán-Mendoza, R.; Pizano, J. Preferencia de Bactericera cockerelli (Sulc.) (Hemiptera: Psyllidae) y su parasitoide Tamarixia triozae (Burk) (Hymenoptera: Eulophidae) por diferentes solanáceas. Entomol. Mex. 2014, 1, 814–819. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayo-Hernández, J.; Vega-Chávez, J.L.; Hernández-Juárez, A.; Rodríguez-Pagaza, Y.; Valenzuela-Soto, J.H.; Flores-Olivas, A. Tomato Varieties Influence the Performance of Tamarixia triozae (Hymenoptera: Eulophidae) on Bactericera cockerelli (Hemiptera: Triozidae) Nymphs. Insects 2022, 13, 825. https://doi.org/10.3390/insects13090825
Mayo-Hernández J, Vega-Chávez JL, Hernández-Juárez A, Rodríguez-Pagaza Y, Valenzuela-Soto JH, Flores-Olivas A. Tomato Varieties Influence the Performance of Tamarixia triozae (Hymenoptera: Eulophidae) on Bactericera cockerelli (Hemiptera: Triozidae) Nymphs. Insects. 2022; 13(9):825. https://doi.org/10.3390/insects13090825
Chicago/Turabian StyleMayo-Hernández, Juan, Jorge Luis Vega-Chávez, Agustín Hernández-Juárez, Yolanda Rodríguez-Pagaza, José Humberto Valenzuela-Soto, and Alberto Flores-Olivas. 2022. "Tomato Varieties Influence the Performance of Tamarixia triozae (Hymenoptera: Eulophidae) on Bactericera cockerelli (Hemiptera: Triozidae) Nymphs" Insects 13, no. 9: 825. https://doi.org/10.3390/insects13090825
APA StyleMayo-Hernández, J., Vega-Chávez, J. L., Hernández-Juárez, A., Rodríguez-Pagaza, Y., Valenzuela-Soto, J. H., & Flores-Olivas, A. (2022). Tomato Varieties Influence the Performance of Tamarixia triozae (Hymenoptera: Eulophidae) on Bactericera cockerelli (Hemiptera: Triozidae) Nymphs. Insects, 13(9), 825. https://doi.org/10.3390/insects13090825