
Flight Performance, Fecundity, and Ovary Development of Grapholita molesta (Lepidoptera: Torticidae) at Different Ages


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Vitellogenin Levels
2.3. Ovary Development
2.4. Daily Fecundity
2.5. Flight Ability
2.6. Statistical Analysis
3. Results
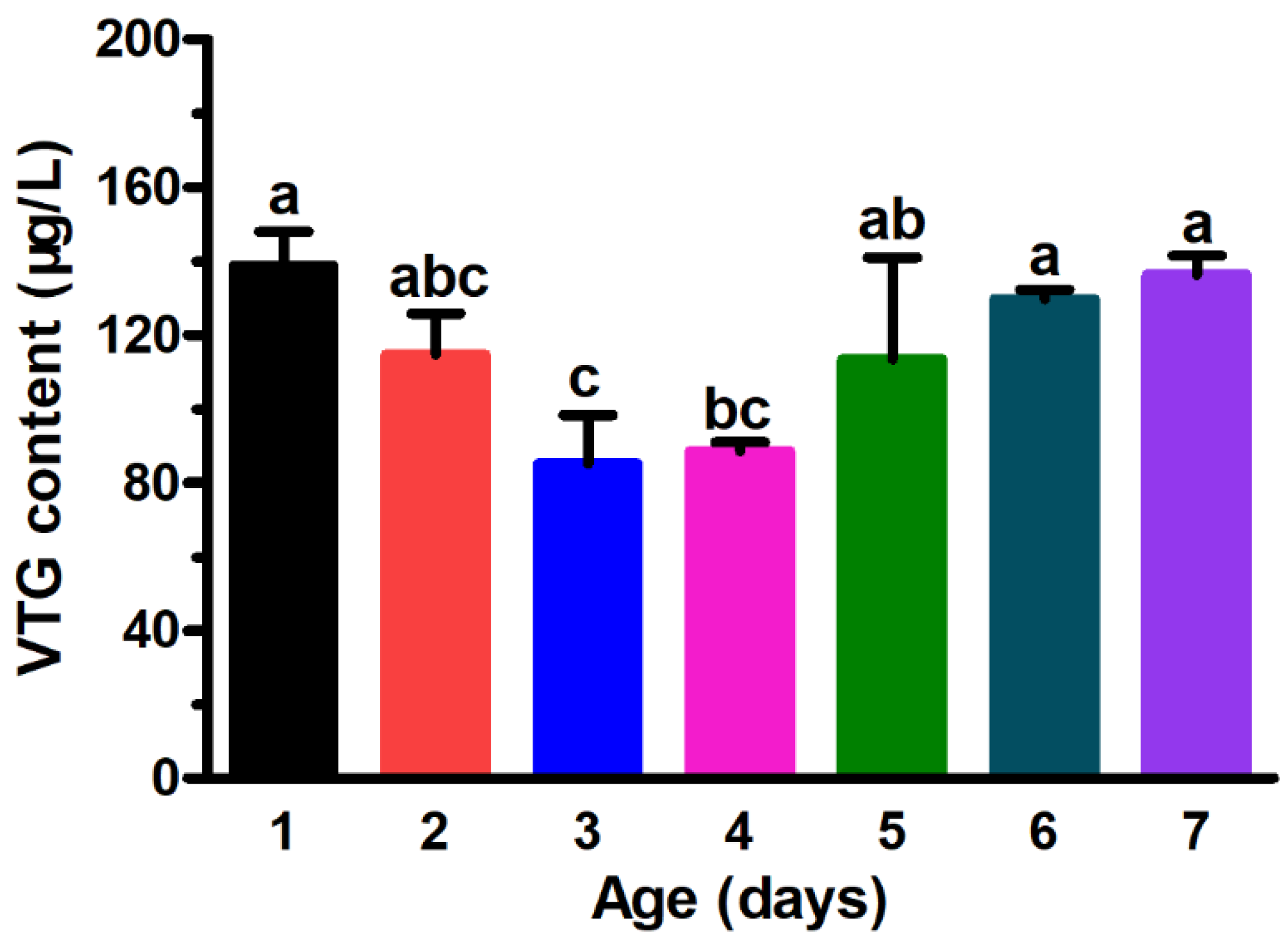
3.1. Vitellogenin Levels
3.2. Ovary Development
3.3. Fecundity
3.4. Flight Ability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Piñero, J.C.; Dorn, S. Response of female oriental fruit moth to volatiles from apple and peach trees at three phenological stages. Entomol. Exp. Appl. 2009, 131, 67–74. [Google Scholar] [CrossRef]
- Myers, C.T.; Hull, L.A.; Krawczyk, G. Effects of orchard host plants on the oviposition preference of the oriental fruit moth (Lepidoptera: Tortricidae). J. Econ. Entomol. 2006, 99, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Kirk, H.; Dorn, S.; Mazzi, D. Worldwide population genetic structure of the oriental fruit moth (Grapholita molesta), a globally invasive pest. BMC Ecol. 2013, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Najar-Rodriguez, A.; Orschel, B.; Dorn, S. Season-long volatile emissions from peach and pear trees in situ, overlapping profiles, and olfactory attraction of an oligophagous fruit moth in the laboratory. J. Chem. Ecol. 2013, 39, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.F.; Qiao, H.L.; Xu, Z.C.; Cheng, J.; Zong, S.X.; Luo, Y.Q. Comparative analysis of peach and pear fruit volatiles attractive to the oriental fruit moth, Cydia molesta. J. Plant Interact. 2014, 9, 388–395. [Google Scholar] [CrossRef]
- Zheng, Y.; Peng, X.; Liu, G.M.; Pan, H.Y.; Dorn, S.; Chen, M.H. High genetic diversity and structured populations of the oriental fruit moth in its range of origin. PLoS ONE. 2013, 8, e78476. [Google Scholar] [CrossRef]
- Monteiro, L.B.; Witt, L.G.; Guiloski, I.C.; dos Santos, R.S.S.; de Assis, H.C.S. Evaluation of resistance management for the oriental fruit moth (Lepidoptera: Tortricidae) to insecticides in Brazilian apple orchards. J. Econ. Entomol. 2020, 3, 1411–1418. [Google Scholar] [CrossRef]
- Pree, D.J.; Whitty, K.J.; Van Driel, L.; Walker, G.M. Resistance to insecticides in oriental fruit moth populations (Grapholita molesta) from the Niagara Peninsula of Ontario. Can. Entomol. 1998, 130, 245–256. [Google Scholar] [CrossRef]
- Kanga, L.H.B.; Pree, D.J.; van Lier, J.L.; Walker, G.M. Monitoring for resistance to organophosphorus, carbamate, and pyrethroid insecticides in the oriental fruit moth (Lepidoptera: Tortricidae). Can. Entomol. 1999, 131, 441–450. [Google Scholar] [CrossRef]
- Siegwart, M.; Monteiro, L.B.; Maugin, S.; Olivares, J.; Carvalho, S.M.; Sauphanor, B. Tools for resistance monitoring in oriental fruit moth (Lepidoptera: Tortricidae) and first assessment in Brazilian populations. J. Econ. Entomol. 2011, 2, 636–645. [Google Scholar] [CrossRef]
- Knipling, E. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Saito, O. Flight activity changes of the cotton bollworm, Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae), by aging and copulation as measured by flight actograph. Appl. Entomol. Zool. 2000, 35, 53–61. [Google Scholar] [CrossRef]
- Tu, Y.G.; Wu, K.M.; Xue, F.S.; Lu, Y.H. Laboratory evaluation of flight activity of the common cutworm, Spodoptera litura (Lepidoptera: Noctuidae). Insect Sci. 2010, 17, 53–59. [Google Scholar] [CrossRef]
- Davey, K.G. The female reproductive tract. In Comprehensive Insect Physiology Biochemistry and Pharmacology; Kerkut, G., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 15–36. [Google Scholar]
- Büning, J. The Insect Ovary: Ultrastructure, Previtellogenic Growth and Evolution; Chapman and Hall: London, UK, 1999. [Google Scholar]
- Holland, R.A.; Wiketski, M.; Wilcove, D. How and why do insects migrate? Science 2006, 313, 794–796. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.R.; Taylor, P.W. Flight ability procedures for mass-reared Queensland fruit flies, Bactrocera tryoni: An assessment of some variations. Entomol. Exp. Appl. 2010, 136, 308–311. [Google Scholar] [CrossRef]
- Marec, F.; Vreysen, M.J.B. Advances and challenges of using the sterile insect technique for the management of pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef]
- Fu, X.W.; Chang, H.; He, L.M.; Zhao, S.Y.; Wu, K.M. Flight performance of Macdunnoughia crassisigna (Lepidoptera: Noctuidae). Bull. Entomol. Res. 2017, 107, 715–723. [Google Scholar] [CrossRef]
- Yu, Q.; Feng, Y.T.; Guo, X.J.; Guo, G.M.; Hao, C.; Fan, R.J. Effects of humidity on the survival and fecundity of Grapholita molesta (Lepidoptera Tortricidae). Acta Entomol. Sin. 2017, 60, 659–665. [Google Scholar]
- Zhu, L.F.; Wei, L.; Zha, J.M.; Na, L.; Wang, Z.J. Chronic thiamethoxam exposure impairs the HPG and HPT axes in adult Chinese rare minnow (Gobiocypris rarus): Docking study, hormone levels, histology, and transcriptional responses. Ecotox. Environ. Saf. 2019, 185, 109683. [Google Scholar] [CrossRef]
- Grodowitz, M.J.; Elliot, B.; Evans, R.; Rojas, G.; Ramos, J.M. Development of a physiological age-grading system for Nezara viridula (Hemiptera: Pentatomidae). J. Insect Sci. 2020, 20, 6. [Google Scholar] [CrossRef]
- Kuang, X.Q.; Calvin, D.; Knapp, M.; Poston, F. Female European corn borer (Lepidoptera: Crambidae) ovarian developmental stages: Their association with oviposition and use in a classification system. J. Econ. Entomol. 2004, 97, 828–835. [Google Scholar]
- Su, S.; Wang, X.T.; Jian, C.Z.; Ignatus, A.D.; Zhang, X.H.; Peng, X.; Chen, M.H. Life-history traits and flight capacity of Grapholita molesta (Lepidoptera: Tortricidae) using artificial diets with varying sugar content. J. Econ. Entomol. 2021, 114, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.J.; Chen, E.Z.; Li, H.Z. ZIR: Modified Wilcoxon Rank Test and Kruskal Wallis Test for Zero-Inflated Data, ZIR (Zero-Inflated Rank Test) R Package Version 1.0.0, 2021. Available online: https://github.com/chvlyl/ZIR. (accessed on 7 August 2022).
- Balestrino, F.; Puggioli, A.; Carrieri, M.; Bouyer, J.; Bellini, R. Quality control methods for Aedes albopictus sterile male production. PLoS Negl. Trop. Dis. 2017, 11, e0005881. [Google Scholar] [CrossRef] [PubMed]
- Bond, J.G.; Osorio, A.R.; Avila, N.; Gomez-Simuta, Y.; Marina, C.F.; Fernandez-Salas, I.; Liedo, P.; Dor, A.; Carvalho, D.O.; Bourtzis, K.; et al. Optimization of irradiation dose to Aedes aegypti and Ae. albopictus in a sterile insect technique program. PLoS ONE. 2019, 14, e0212520. [Google Scholar] [CrossRef] [PubMed]
- Hoddle, M.S.; Hoddle, C.D.; Milosavljevic, I. Quantification of the life time flight capabilities of the south American palm weevil, Rhynchophorus palmarum (L.) (Coleoptera: Curculionidae). Insects 2021, 12, 126. [Google Scholar] [CrossRef]
- Barkan, S.; Hoffman, A.; Hezroni, A.; Soroker, V. Flight performance and dispersal potential of red palm weevil estimated by repeated flights on flight mill. J. Insect Behav. 2018, 31, 66–82. [Google Scholar] [CrossRef]
- Luo, L.Z.; Johnson, S.J.; Hammond, A.M.; Lopez, J.D.; Geaghan, J.P.; Beerwinkle, K.R.; Westbrook, J.K. Determination and consideration of flight potential in a laboratory population of true armyworm (Lepidoptera: Noctuidae). Environ. Entomol. 2002, 1, 1–9. [Google Scholar] [CrossRef]
- Su, S.; Zhang, X.; Jian, C.; Huang, B.; Peng, X.; Vreysen, M.J.B.; Chen, M. Effects of adult feeding treatments on longevity, fecundity, flight ability, and energy metabolism enzymes of Grapholita molesta moths. Insects 2022, 13, 725. [Google Scholar] [CrossRef]
- Nation, J.L. Insect Physiology and Biochemistry, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 223–349. [Google Scholar]
- Jiang, X.F.; Luo, L.Z.; Sappington, T.W. Relationship of flight and reproduction in beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae), a migrant lacking the oogenesis-flight syndrome. J. Insect Physiol. 2010, 56, 1631–1637. [Google Scholar] [CrossRef]
- Ishiguri, Y.; Shirai, Y. Flight activity of the peach fruit moth, Carposina sasakii (Lepidoptera: Carposinidae), measured by a flight mill. Appl. Entomol. Zool. 2004, 39, 127–131. [Google Scholar] [CrossRef]
- Shirai, Y. Laboratory evaluation of flight ability of the Oriental corn borer, Ostrinia furnacalis (Lepidoptera: Pyralidae). Bull. Entomol. Res. 1998, 88, 327–333. [Google Scholar] [CrossRef]
- Shirai, Y.; Kosugi, Y.; Noguchi, I. Effects of sex, mating status and temperature on flight activity of the Oriental tea tortrix, Homona magnanima (Lepidoptera: Tortricidae). Appl. Entomol. Zool. 1998, 33, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Avalos, J.A.; Marti-Campoy, A.; Soto, A. Study of the flying ability of Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) adults using a computer-monitored flight mill. Bull. Entomol. Res. 2014, 104, 462–470. [Google Scholar] [CrossRef]
- Yu, H.L.; Zhang, Y.J.; Wu, K.M.; Wyckhuys, K.A.G.; Guo, Y.Y. Flight potential of Microplitis mediator, a parasitoid of various lepidopteran pests. BioControl 2009, 54, 183–193. [Google Scholar] [CrossRef]
- Hughes, J.; Dorn, S. Sexual differences in the flight performance of the oriental fruit moth. Entomol. Exp. Appl. 2002, 103, 171–182. [Google Scholar] [CrossRef]
- Timm, A.E.; Geertsema, H.; Warnich, L. Population genetic structure of Grapholita molesta (Lepidoptera: Tortricidae) in South Africa. Ann. Entomol. Soc. Am. 2008, 101, 197–203. [Google Scholar] [CrossRef]
- Dorn, S.; Gu, H. Laboratory evaluation of influence of surface residues of azinphos-methyl on adult activity of the codling moth, Cydia pomonella L. (Lepidoptera: Tortricidae). IOBC/WPRS Bull. 1999, 22, 195–199. [Google Scholar]
- Klassen, W. Area-wide integrated pest management and the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 39–69. [Google Scholar]
- Graziosi, I.; Rieske, L.K. Potential fecundity of a highly invasive gall maker, Dryocosmus kuriphilus (Hymenoptera: Cynipidae). Environ. Entomol. 2014, 43, 1053–1058. [Google Scholar] [CrossRef] [PubMed]
- Souna, D.A.; Bokonon-Ganta, A.; Ravallec, M.; Cusumano, A.; Pittendrigh, B.R.; Volkoff, A.N.; Tamo, M. An insight in the reproductive biology of Therophilus javanus (Hymenoptera, Braconidae, and Agathidinae), a potential biological control agent against the legume pod borer (Lepidoptera, Crambidae). Psyche 2017, 2017, 3156534. [Google Scholar]
- Patrick, C.; Tobin, P.C.; Bolyard, J.L.; Onufrieva, K.S.; Hickman, A.D. The effect of male and female age on Lymantria dispar (Lepidoptera: Lymantriidae) fecundity. J. Econ. Entomol. 2014, 107, 1076–1083. [Google Scholar]
- Chen, L.; Wang, G.L.; Wei, H.Y. Circadian rhythm of emergence and reproduction of Micromelalopha troglodyta (Lepidoptera: Notodontidae). Chin. J. Appl. Ecol. 2014, 25, 2425–2430. [Google Scholar]
- Papaj, D.R. Ovarian dynamics and host use. Annu. Rev. Entomol. 2000, 45, 423–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, S.; Zhang, X.; Zhang, J.; Huang, B.; Jian, C.; Peng, X.; Vreysen, M.J.B.; Chen, M. Flight Performance, Fecundity, and Ovary Development of Grapholita molesta (Lepidoptera: Torticidae) at Different Ages. Insects 2022, 13, 837. https://doi.org/10.3390/insects13090837
Su S, Zhang X, Zhang J, Huang B, Jian C, Peng X, Vreysen MJB, Chen M. Flight Performance, Fecundity, and Ovary Development of Grapholita molesta (Lepidoptera: Torticidae) at Different Ages. Insects. 2022; 13(9):837. https://doi.org/10.3390/insects13090837
Chicago/Turabian StyleSu, Sha, Xiaohe Zhang, Jilong Zhang, Baojian Huang, Chengzhi Jian, Xiong Peng, Marc J. B. Vreysen, and Maohua Chen. 2022. "Flight Performance, Fecundity, and Ovary Development of Grapholita molesta (Lepidoptera: Torticidae) at Different Ages" Insects 13, no. 9: 837. https://doi.org/10.3390/insects13090837
APA StyleSu, S., Zhang, X., Zhang, J., Huang, B., Jian, C., Peng, X., Vreysen, M. J. B., & Chen, M. (2022). Flight Performance, Fecundity, and Ovary Development of Grapholita molesta (Lepidoptera: Torticidae) at Different Ages. Insects, 13(9), 837. https://doi.org/10.3390/insects13090837